
Analysis of the Nutritional Composition and Drought Tolerance Traits of Sweet Potato: Selection Criteria for Breeding Lines


Abstract
:1. Introduction
2. Results and Discussion
2.1. Composite Analysis
2.2. Sugars and Starch
2.3. Vitamins
2.4. Mineral Content
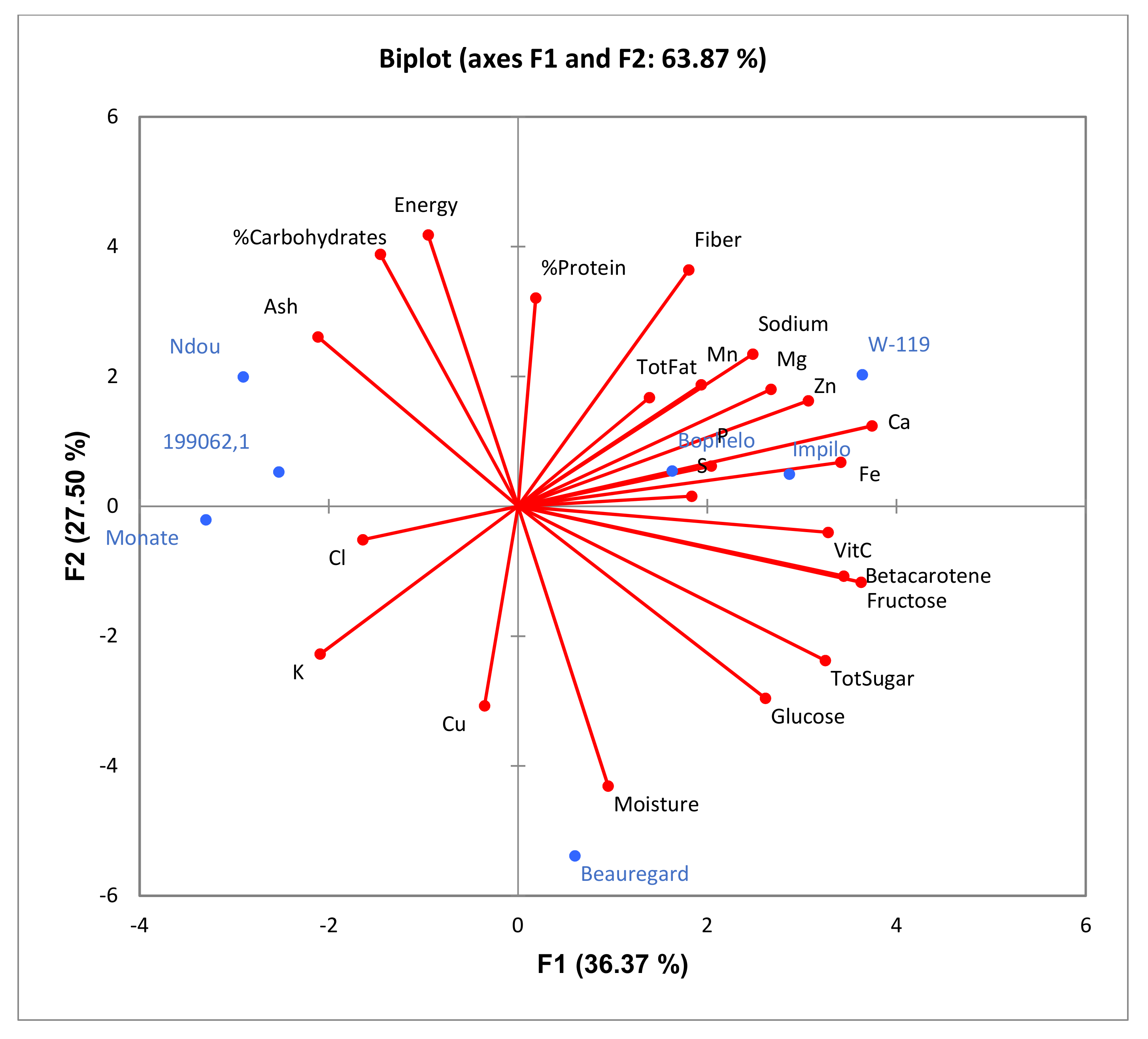
2.5. Principal Component Analysis
2.6. Correlation among Nutrients
2.7. General Discussion on Nutritional Composition
2.8. Results of Biochemical Assays Screening for Drought Tolerance
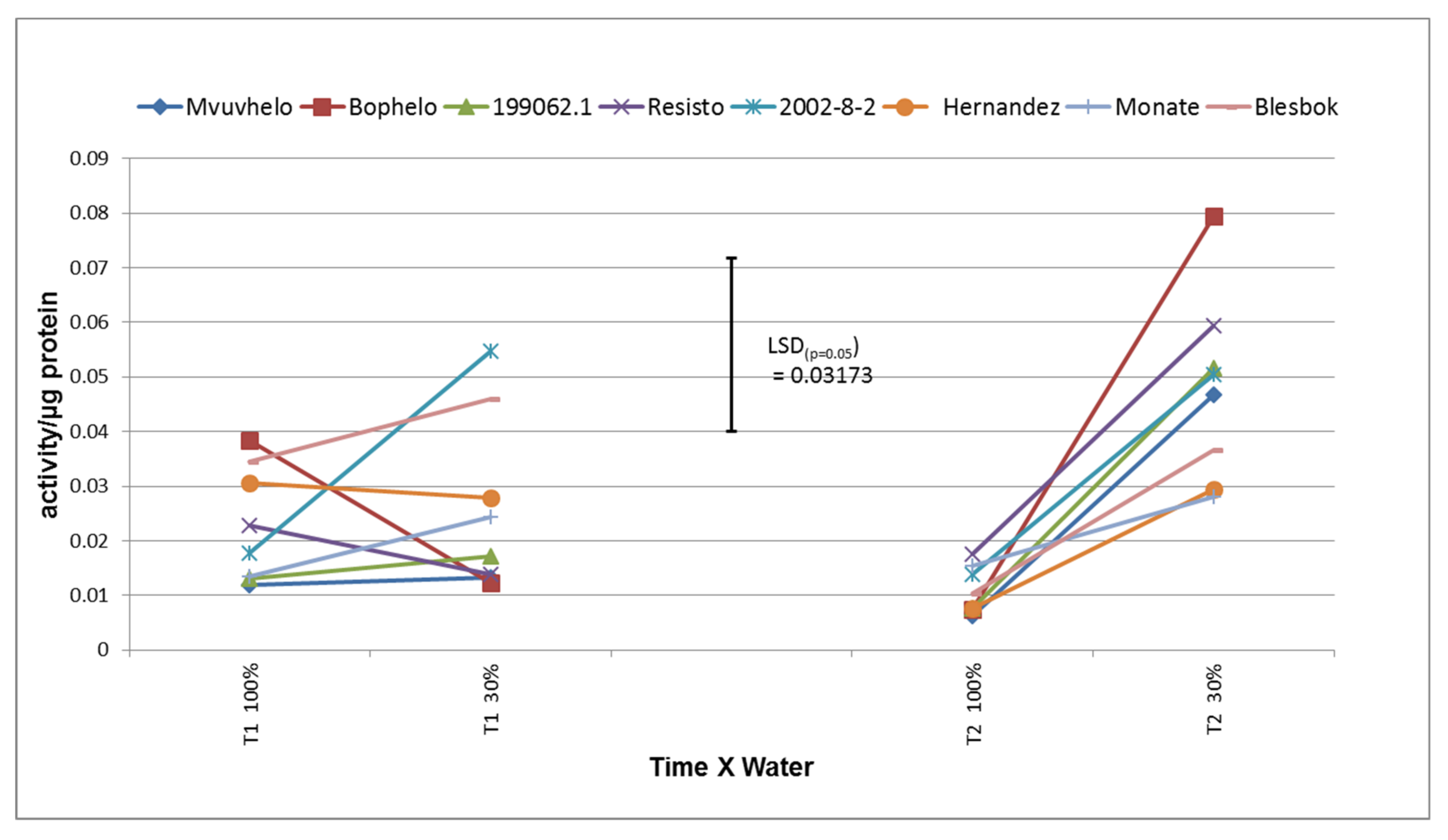
2.8.1. Ascorbate Peroxidase (APX)
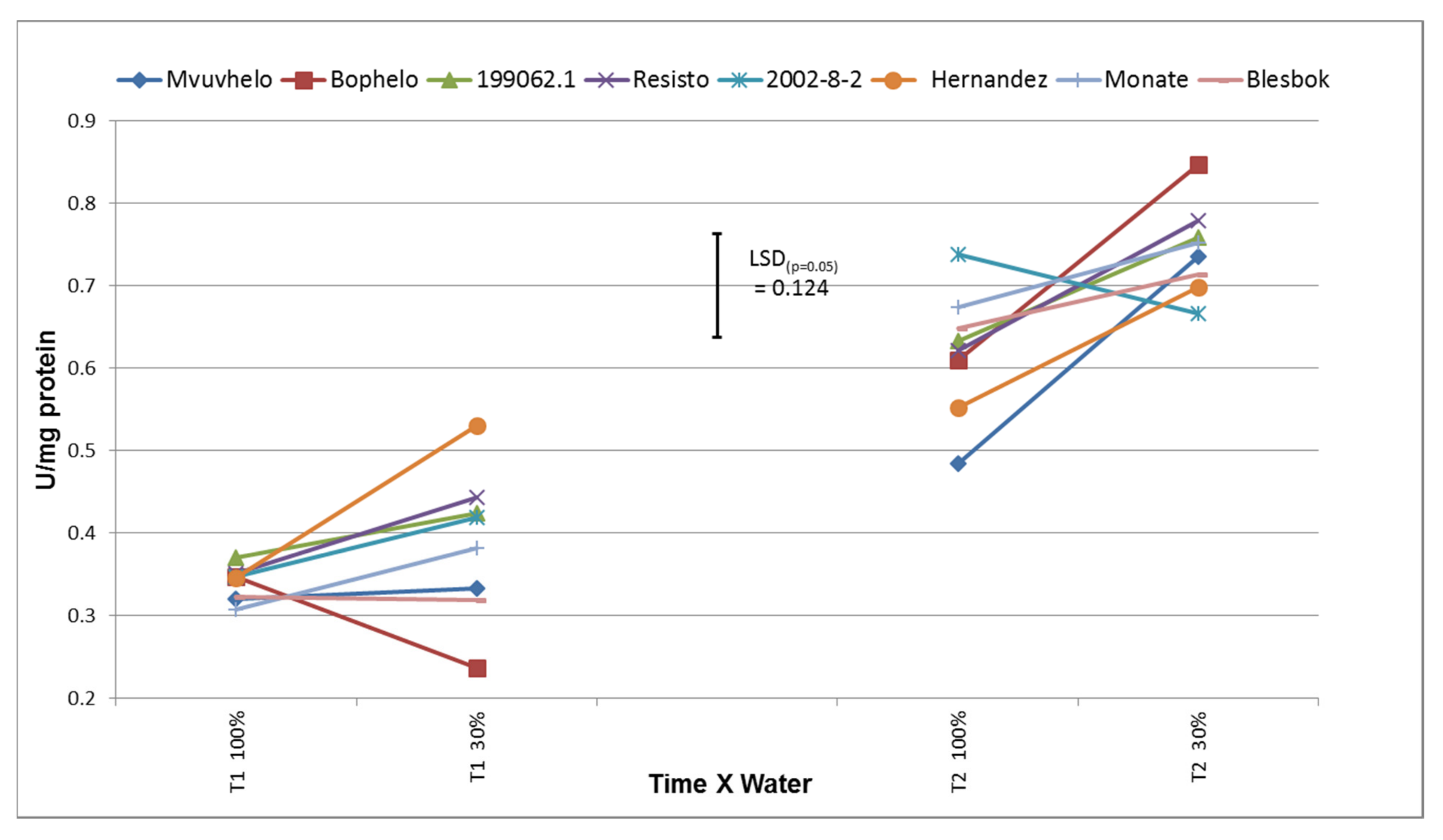
2.8.2. Super Oxide Dismutase (SOD)
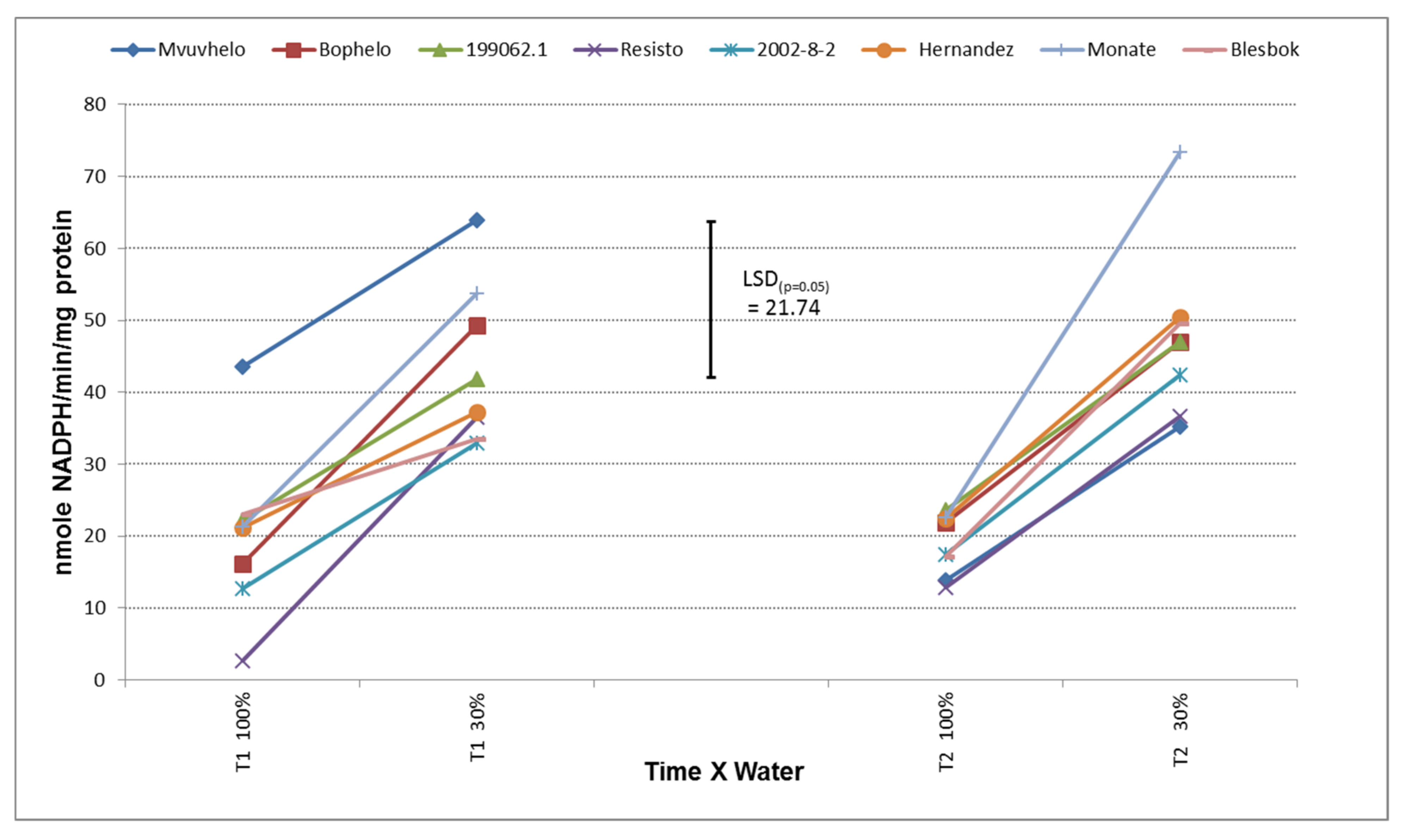
2.8.3. Glutathione Reductase (GR)
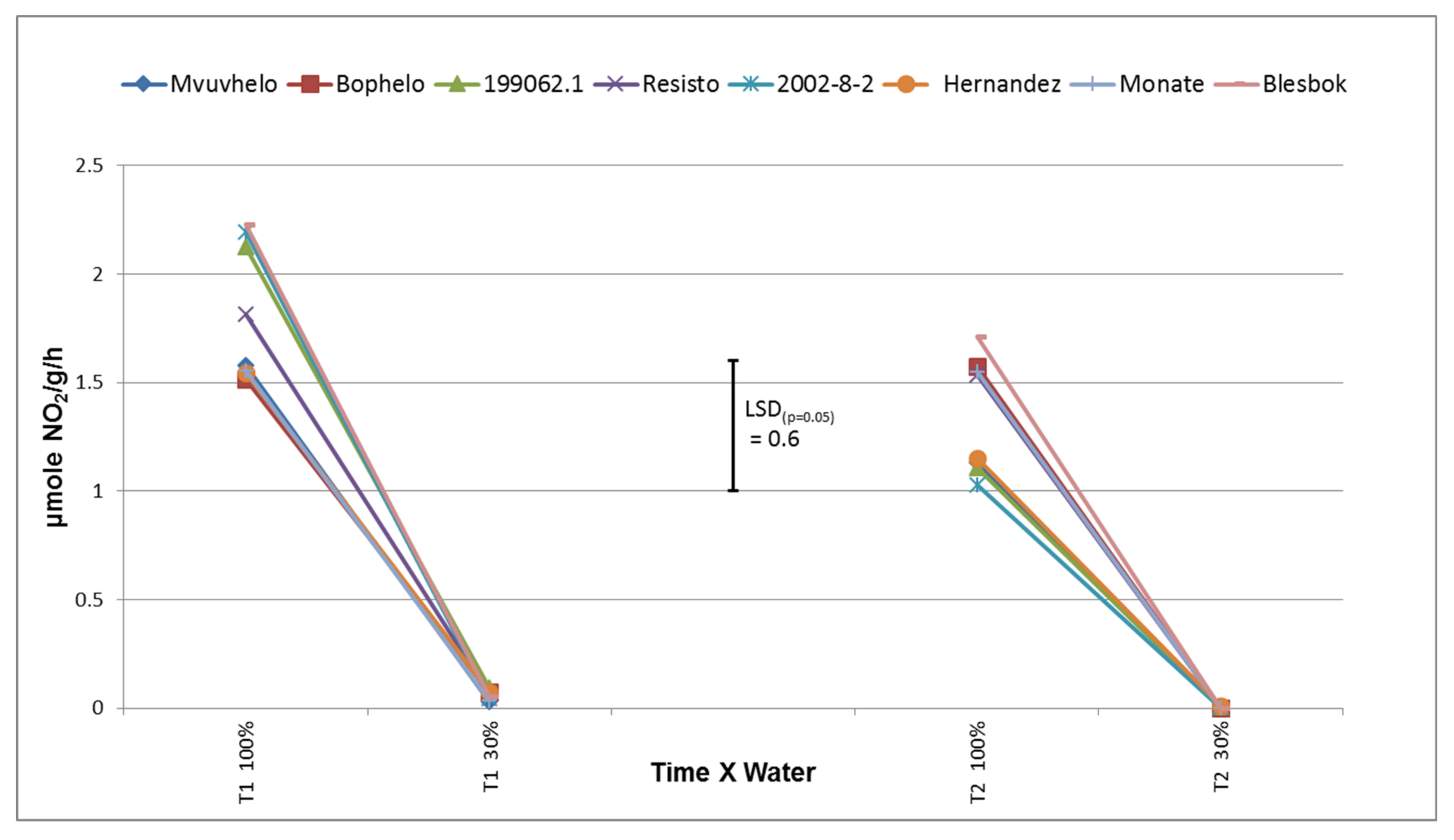
2.8.4. Nitrate Reductase (NR)
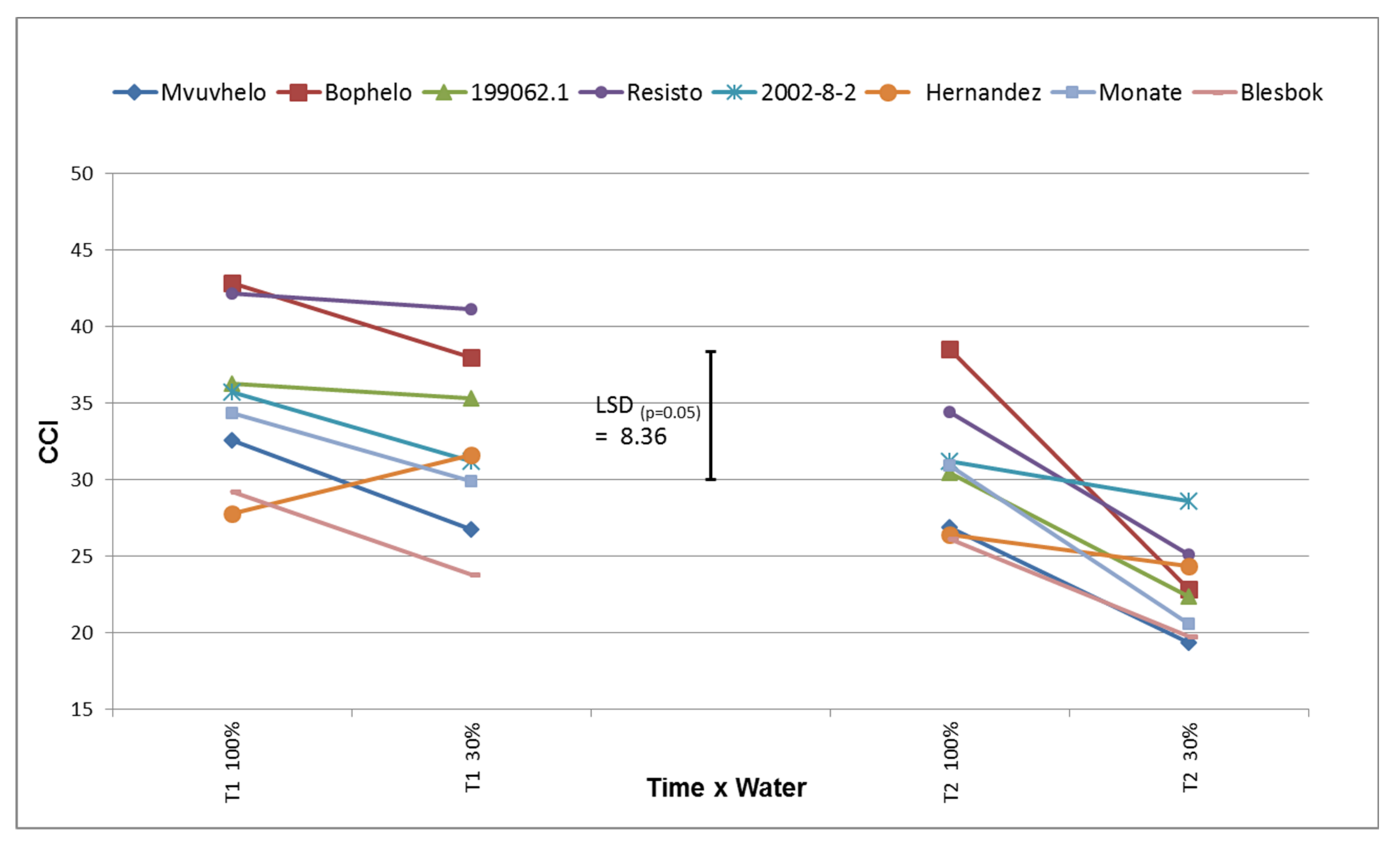
2.8.5. Chlorophyll Content Index (CCI)
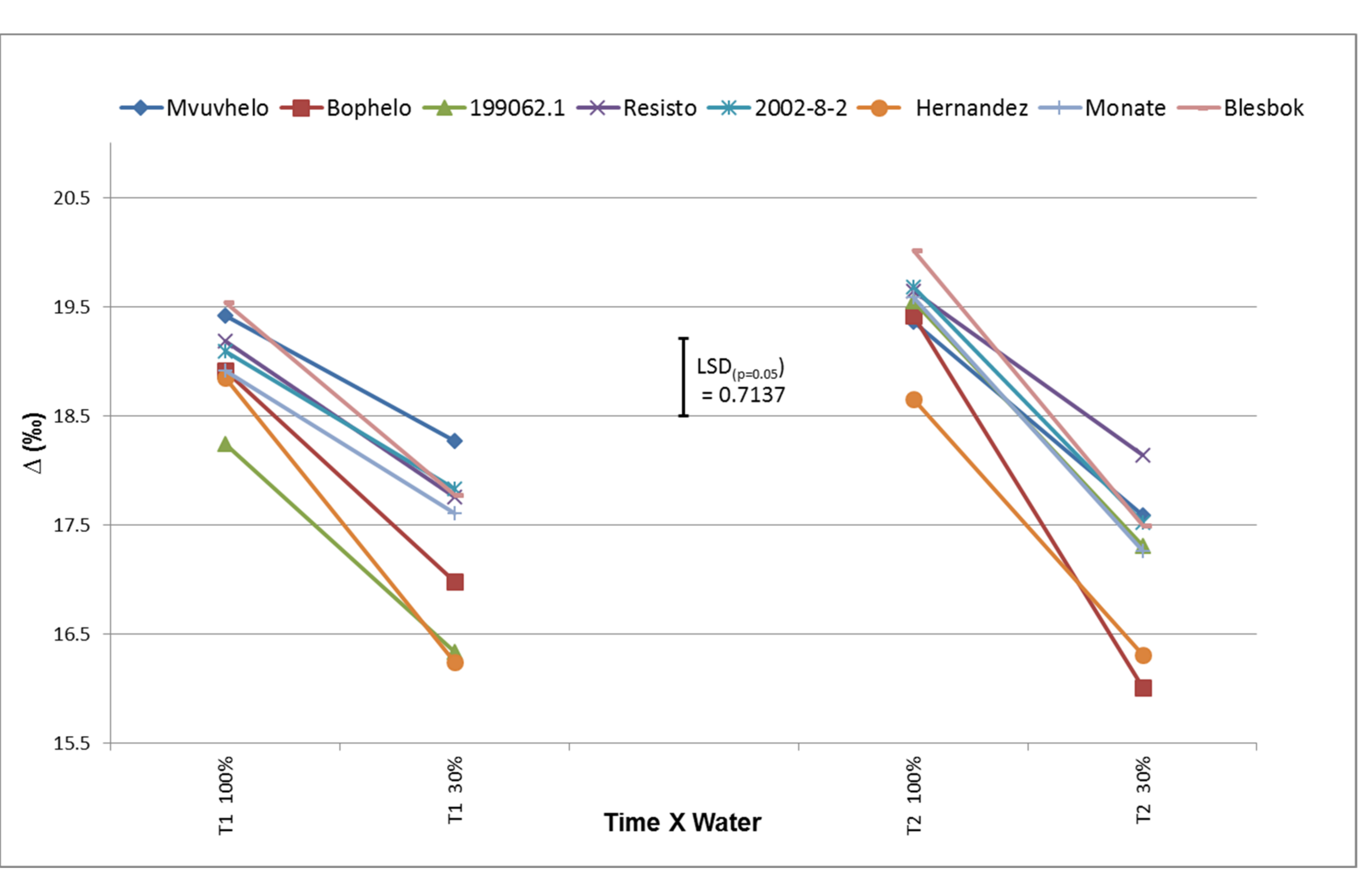
2.8.6. 13C Discrimination (Δ13C)
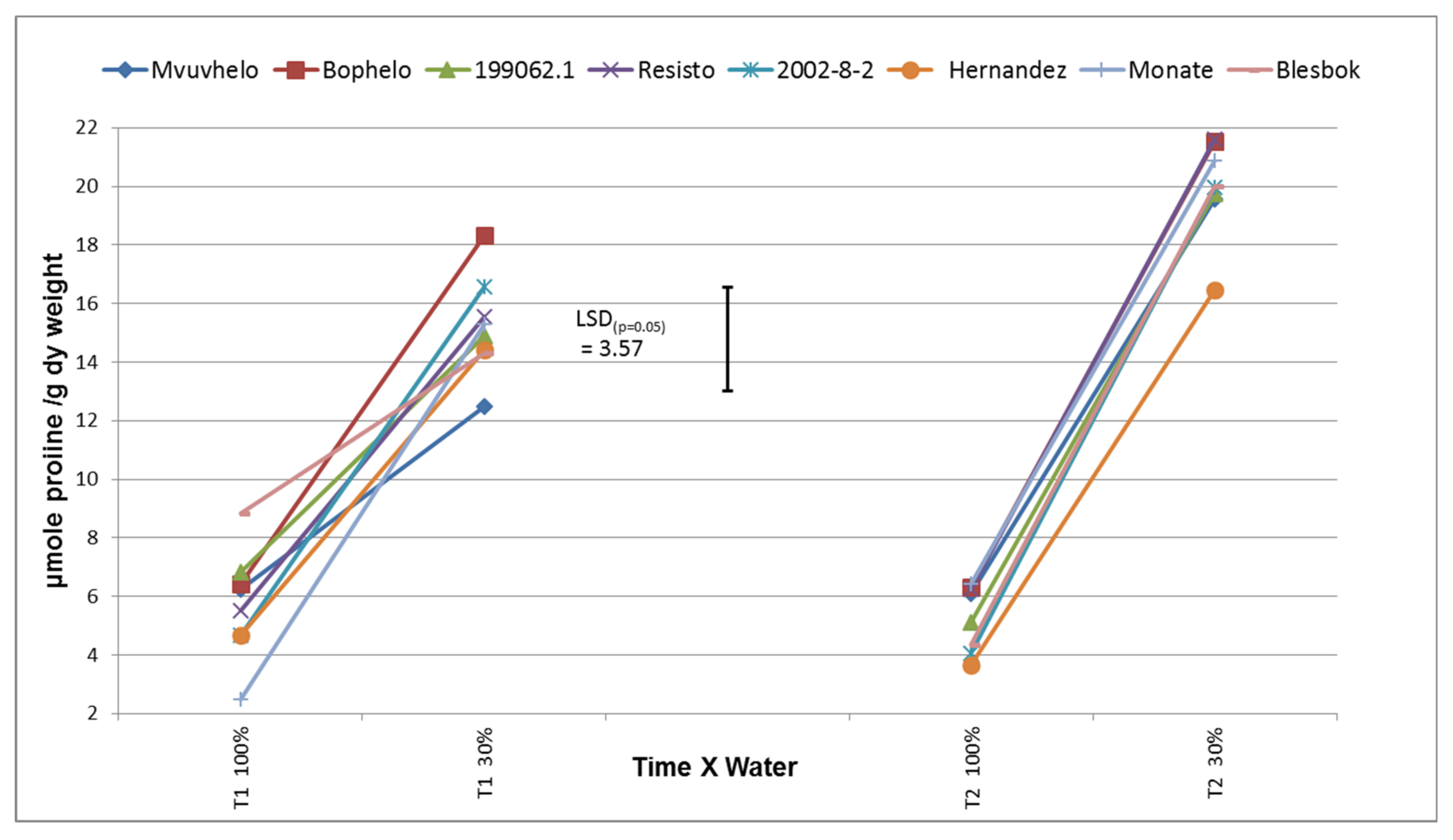
2.8.7. Free Proline Content
2.8.8. Sweet Potato Yield
2.8.9. Relationship between Biochemical Compounds and Yield
3. Conclusions
4. Materials and Methods
4.1. Breeding Procedures
4.2. Plant Material and Sampling for Nutritional Analysis
4.3. Nutritional Analysis
4.4. Experimental Conditions for Drought Stress Screening
4.5. Biochemical Assays Related to Drought Stress
4.5.1. Ascorbate Peroxidase (APX)
4.5.2. Superoxide Dismutase (SOD)
4.5.3. Glutathione Reductase (GR)
4.5.4. Nitrate Reductase (NR)
4.5.5. Chlorophyll Content Index (CCI)
4.5.6. Carbon Isotope Discrimination
4.5.7. Proline Analysis
4.5.8. Yield Parameters
4.5.9. Statistical Analysis
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAOTSTAT. The State of Global Nutrition. 2021. Available online: https://globalnutritionreport.org/reports/2021-global-nutrition-report/ (accessed on 17 April 2022).
- Mwanga, R.O.M.; Swanckaert, J.; Pereira, G.D.S.; Andrade, M.I.; Makunde, G.; Grüneberg, W.J.; Kreuze, J.; David, M.; De Boeck, B.; Carey, E.; et al. Breeding Progress for Vitamin A, Iron and Zinc Biofortification, Drought Tolerance, and Sweetpotato Virus Disease Resistance in Sweetpotato. Front. Sustain. Food Syst. 2021, 5, 616674. [Google Scholar] [CrossRef]
- FAOSTAT. Crops and Livestock Products. 2021. Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on 17 April 2022).
- Truong, V.D.; Avula, R.Y.; Pecota, K.V.; Yencho, G.C. Sweetpotato Production, Processing, and Nutritional Quality. In Handbook of Vegetables and Vegetable Processing, 2nd ed.; Siddiq, M., Uebersax, M.A., Eds.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2018; Volume II. [Google Scholar]
- Echodu, R.; Edema, H.; Wokorach, G.; Zawedde, C.; Otim, G.; Luambano, N.; Ateka, E.M.; Asiimwe, T. Farmers’ practices and their knowledge of biotic constraints to sweetpotato production in East Africa. Physiol. Mol. Plant Pathol. 2018, 105, 3–16. [Google Scholar] [CrossRef] [PubMed]
- DALLRD Sweetpotato Report: A Profile of the South African Sweetpotato Market Value Chain. 2021. Available online: http://webapps.daff.gov.za/amis/Link.amis?method=VegetablesValueChain (accessed on 24 April 2022).
- Motsa, N.M.; Modi, A.T.; Mabhaudhi, T. Sweet potato (Ipomoea batatas L.) as a drought tolerant and food security crop. S. Afr. J. Sci. 2015, 111, 8. [Google Scholar] [CrossRef] [Green Version]
- Bester, C.; Louw, J.H. Sweet potato breeding in South Africa. J. S. Afr. Soc. Hortic. 1992, 2, 96–98. [Google Scholar]
- Jones, A. Sweetpotato. In Hybridization of Crop Plants; American Society of Agronomy-Crop Science Society of America: Madison, WI, USA, 1980; Volume 46, pp. 645–655. [Google Scholar]
- Collins, W.W.; Wilson, L.G.; Arrendell, S.; Dickey, L.F. Genotype x environment interactions in sweetpotato yield and quality factors. J. Am. Soc. Hortic. Sci. 1987, 112, 579–583. [Google Scholar]
- Naskar, S.K.; Singh, D.P. Genotype x environment interaction for tuber yield in sweetpotato. J. Root Crops 1992, 18, 85–88. [Google Scholar]
- Laurie, S.M.; Tjale, S.S.; van den Berg, A.A.; Mtileni, M.M.; Labuschagne, M.T. Agronomic performance of new cream to yellow-orange sweetpotato varieties in diverse environments across South Africa. S. Afr. J. Plant Soil 2015, 32, 147–155. [Google Scholar] [CrossRef]
- FAO; IFAD; UNICEF; WFP; WHO. The State of Food Security and Nutrition in the World 2019: Safeguarding against Economic Slowdowns and Downturns; FAO: Rome, Italy, 2019. [Google Scholar]
- Bovell-Benjamin, A.C. Sweet potato: A review of its past, present, and future role in human nutrition. In Advances in Food and Nutrition Research; Department of Food and Nutritional Sciences, Tuskegee/NASA Center for Food and Environmental Systems for Human Exploration of Space (CFESH) Tuskegee University: Tuskegee, AL, USA, 2007. [Google Scholar]
- Low, J.W.; Ortiz, R.; Vandamme, E.; Andrade, M.; Biazin, B.; Grüneberg, W.J. Nutrient-Dense Orange-Fleshed Sweetpotato: Advances in Drought-Tolerance Breeding and Understanding of Management Practices for Sustainable Next-Generation Cropping Systems in Sub-Saharan Africa. Front. Sustain. Food Syst. 2020, 4, 50. [Google Scholar] [CrossRef]
- Laurie, S.M.; Booyse, M.; Labuschagne, M.T.; Greyling, M.M. Multi-environment performance of new orange-fleshed sweetpotato varieties in South Africa. Crop Sci. 2015, 55, 1585–1595. [Google Scholar] [CrossRef]
- Laurie, S.M.; Mtileni, M.M.; Mphela, W.M.; Tjale, S.S. Performance of informal market sweet potato cultivars in on-farm trials in South Africa. Open Agric. 2017, 2, 431–441. [Google Scholar] [CrossRef] [Green Version]
- Schönfeldt, H.C.; Hall, N.; Pretorius, B. The important role of food composition in policies and programmes for better public health: A South Africa case study. Food Chem. 2018, 238, 94–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Placide, R.; Shimelis, H.; Laing, M.; Gahakwa, D. Physiological mechanisms and conventional breeding of sweet potato (Ipomoea batatas (L.) Lam.) to drought-tolerance. Afr. J. Agric. Res. 2013, 8, 1837–1846. [Google Scholar]
- Niang, I.; Ruppel, O.C.; Abdrabo, M.A.; Essel, A.; Lennard, C.; Padgham, J.; Urquhart, P. Africa. In Climate Change 2014: Impacts, Adaptation, and Vulnerability. Part B: Regional Aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Barros, V.R., Field, C.B., Dokken, D.J., Mastrandrea, M.D., Eds.; Cambridge University Press: New York, NY, USA, 2014; pp. 1199–1265. [Google Scholar]
- Ekanayake, I.J.; Collins, W. Effect of irrigation on sweet potato root carbohydrates and nitrogenous compounds. J. Food Agric. Environ. 2004, 2, 243–248. [Google Scholar]
- Gajanayake, B.; Reddy, K.R.; Shankle, M.W.; Arancibia, R.A. Growth, developmental, and physiological responses of two sweetpotato (Ipomoea batatas L. [Lam]) cultivars to early season soil moisture deficit. Sci. Hortic. 2015, 168, 218–228. [Google Scholar] [CrossRef]
- Andrade, M.I.; Naico, A.; Ricardo, J.; Eyzaguirre, R.; Makunde, G.S.; Ortiz, R.; Grűneberg, W.J. Genotype × environment interaction and selection for drought adaptation in sweetpotato (Ipomoea batatas [L.] Lam.) in Mozambique. Euphytica 2016, 209, 261–280. [Google Scholar] [CrossRef] [Green Version]
- Foyer, C.H.; Valadier, M.H.; Migge, A.; Becker, T.W. Drought-Induced effects on nitrate reductase activity and mRNA and on the coordination of nitrogen and carbon metabolism in maize leaves. Plant Physiol. 1998, 117, 283–292. [Google Scholar] [CrossRef] [Green Version]
- Deblonde, P.M.K.; Haverkort, A.J.; Ledent, J.F. Responses of early and late potato cultivars to moderate drought conditions: Agronomic parameters and carbon discrimination. Eur. J. Agron. 1999, 11, 91–105. [Google Scholar] [CrossRef]
- Wang, H.L.; Lee, P.D.; Liu, L.F.; Su, J.C. Effect of sorbitol induced osmotic stress of the changes of carbohydrate and free amino acid pools in sweet potato cell suspension cultures. BBAS 1999, 40, 219–225. [Google Scholar]
- Woolfe, J.A. Sweetpotato an Untapped Food Resource; Cambridge University Press: Cambridge, UK, 1992. [Google Scholar]
- Shekhar, S.; Mishra, D.; Buragohain, A.K.; Chakraborty, S.; Chakraborty, N. Comparative analysis of phytochemicals and nutrient availability in two contrasting cultivars of sweet potato (Ipomoea batatas L.). Food Chem. 2015, 173, 957–965. [Google Scholar] [CrossRef]
- Leighton, C.S. Nutrient and Sensory Quality of Orange-Fleshed Sweet Potato. Master’s Thesis, University of Pretoria, Pretoria, South Africa, 2007. [Google Scholar]
- ILSI (International Life Sciences Institute). Chapter 4: Nutritionally improved sweetpotato. Compr. Rev. Food Sci. Food Safety 2008, 7, 81–91. [Google Scholar] [CrossRef]
- Wolmarans, P.; Danster, N.; Dalton, A.; Rossouw, K.; Schonfeldt, H. Condensed Food Composition Tables for South Africa; Medical Research Council: Cape Town, South Africa, 2010.
- Trumbo, P.; Yates, A.A.; Schlicker, S.; Poos, M. Dietary reference intakes: Vitamin A, vitamin K, arsenic, boron, copper, iodine, iron, manganese, molybdenum, nickel, silicon, vanadium, and zinc. J. Am. Diet. Assoc. 2001, 101, 294–301. [Google Scholar] [CrossRef]
- Teow, C.C.; Truong, V.-D.; McFeeters, R.F.; Thompson, R.L.; Pecota, K.V.; Yencho, G.C. Antioxidant activities, phenolic and β-carotene contents of sweet potato genotypes with varying flesh colours. Food Chem. 2007, 103, 829–838. [Google Scholar] [CrossRef]
- Ojimelukwe, P.C.; Okpanku, U. Soil Nutrient Management Practices Influence the Carotenoid Content and Profile of Orange Fleshed Sweet Potato Variety (UMUSPO 3). Sustain. Food Prod. 2020, 8, 38–46. [Google Scholar] [CrossRef]
- Laurie, S.M.; van Jaarsveld, P.J.; Faber, M.; Philpott, M.F.; Labuschagne, M.T. Trans-β-carotene, selected mineral content and potential nutritional contribution of 12 sweetpotato varieties. J. Food Compos. Anal. 2012, 27, 151–159. [Google Scholar] [CrossRef]
- Sanoussi, A.F.; Adjatin, A.; Dansi, A.; Adebowale, A.; Sanni, L.O.; Sanni, A. Mineral Composition of Ten Elites Sweet Potato (Ipomoea Batatas [L.] Lam.) Landraces of Benin. Int. J. Curr. Microbiol. App. Sci. 2016, 5, 103–115. [Google Scholar] [CrossRef]
- Zikankuba, V.L.; Mteremko, D.; James, A. Staple Crops Biofortification Linking Agriculture, Food and Nutrition towards Eliminating Hidden Hunger. Eur. J. Nutr. Food Saf. 2019, 9, 112–121. [Google Scholar] [CrossRef]
- Jongstra, R.; Mwangi, M.N.; Burgos, G.; Zeder, C.; Low, J.W.; Mzembe, G.; Liria, R.; Penny, M.; Andrade, M.I.; Fairweather-Tait, S.; et al. Iron Absorption from Iron-Biofortified Sweetpotato Is Higher Than Regular Sweetpotato in Malawian Women while Iron Absorption from Regular and Iron-Biofortified Potatoes Is High in Peruvian Women. J. Nutr. 2020, 150, 3094–3102. [Google Scholar] [CrossRef] [PubMed]
- Phahlane, C.J.; Laurie, S.M.; Shoko, T.; Manhivi, V.E.; Sivakumar, D. Comparison of caffeoylquinic acids and antioxidant properties of domestic sweet potato (Ipomoea batatas (L.) Lam.) storage roots with overseas varieties. Foods 2022, 11, 1329. [Google Scholar] [CrossRef]
- Zhang, H.; Duan, W.; Xie, B.; Wang, B.; Hou, F.; Li, A.; Dong, S.; Qin, Z.; Wang, Q.; Zhang, L. Root yield, antioxidant capacities, and hormone contents in differnet drought-tolerant sweet potato cultivars treated with ABA under early drought stress. Acta Physiol. Plant. 2020, 42, 132. [Google Scholar] [CrossRef]
- Dalton, D.A.; Russell, S.A.; Hanus, F.J.; Pascoe, G.A.; Evans, H.J. Enzymatic reactions of ascorbate and glutathione that prevent peroxide damage in soybean root nodules. Proc. Natl. Acad. Sci. USA 1986, 83, 3811–3815. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Deng, Z.; Kwak, S. Over expression of CuZn superoxide dismutase (CuZn SOD) and ascorbate peroxidase (APX) in transgenic sweet potato enhances tolerance and recovery from drought stress. Afr. J. Biotechnol. 2010, 9, 8378–8391. [Google Scholar]
- Asada, K. The water-water cycle in chloroplasts: Scavenging of active oxygens and dissipation of excess photons. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 601–639. [Google Scholar] [CrossRef] [PubMed]
- Masoumi, H.; Darvish, F.; Daneshian, J.; Normohammadi, G.; Habibi, D. Effects of water deficit stress on seed yield and antioxidants content in soybean (Glycine max L.) cultivars. Afr. J. Agric. Res. 2011, 6, 1209–1218. [Google Scholar]
- Selote, D.S.; Khanna-Chopra, R. Drought acclimation confers oxidative stress tolerance by inducing co-ordinated antioxidant defense at cellular and subcellular level in leaves of wheat seedlings. Physiol. Plant. 2006, 127, 494–506. [Google Scholar] [CrossRef]
- Malan, C.; Greyling, M.M.; Gresse, J. Correlation between CuZn superoxide dismutase and glutathione reductase, and environmental and xenobiotic stress tolerance in maize inbreds. Plant Sci. 1990, 69, 157–166. [Google Scholar] [CrossRef]
- Mazid, M.; Khan, T.A.; Mohammad, F. Role of nitrate reductase in nitrogen fixation under photosynthetic regulation. World J. Pharm. Res. 2012, 1, 386–414. [Google Scholar]
- Pandey, H.C.; Baig, M.J.; Bhatt, R.K. Effect of moisture stress on chlorophyll accumulation and nitrate reductase activity at vegetative and flowering stage in Avena species. Agric. Sci. Res. J. 2012, 2, 111–118. [Google Scholar]
- Nikolaeva, M.K.; Maevskaya, S.N.; Shugaev, A.G.; Bukhov, N.G. Effect of drought on chlorophyll content and antioxidant enzyme activities in leaves of three wheat cultivars varying in productivity. Russ. J. Plant Physiol. 2010, 57, 87–95. [Google Scholar] [CrossRef]
- Heider, B.; Struelens, Q.; Faye, E.; Flores, C.; Palacios, J.E.; Eyzaguirre, R.; de Haan, S.; Dangles, O. Intraspecific diversity as a reservoir for heat-stress tolerance in sweet potato. Nat. Clim. Chang. 2020, 11, 64–69. [Google Scholar] [CrossRef]
- Akhter, J.; Sabir, A.; Lateef, Z.; Ashraf, M.Y.; Haq, M.A. Relationships between carbon isotope discrimination and grain yield, water use efficiency and growth parameters in wheat (Triticum aestivum L.) under different water regimes. Pak. J. Bot. 2008, 40, 1441–1454. [Google Scholar]
- Leidi, E.; López, M.; Gorham, J.; Gutiérrez, J. Variation in carbon isotope discrimination and other traits related to drought tolerance in upland cotton cultivars under dryland conditions. Field Crop Res. 1999, 61, 109–123. [Google Scholar] [CrossRef]
- Gouveia, C.S.S.; Lebot, V.; De Carvalho, M.P. NIRS Estimation of Drought Stress on Chemical Quality Constituents of Taro (Colocasia esculenta L.) and Sweet Potato (Ipomoea batatas L.) Flours. Appl. Sci. 2020, 10, 8724. [Google Scholar] [CrossRef]
- Armengaud, P.; Thiery, L.; Buhot, N.; March, G.G.-D.; Savoure, A. Transcriptional regulation of proline biosynthesis in Medicago truncatula reveals developmental and environmental specific features. Physiol. Plant. 2004, 120, 442–450. [Google Scholar] [CrossRef] [PubMed]
- Hare, P.; Cress, W. Metabolic implications of stress-induced proline accumulation in plants. Plant Growth Regul. 1997, 21, 79–102. [Google Scholar] [CrossRef]
- Laurie, R.N.; Laurie, S.M.; Du Plooy, C.P.; Finnie, J.F.; Van Staden, J. Yield of Drought-Stressed Sweet Potato in Relation to Canopy Cover, Stem Length and Stomatal Conductance. J. Agric. Sci. 2014, 7, 201–214. [Google Scholar] [CrossRef]
- Van Heerden, P.D.R.; Laurie, R. Effects of prolonged restriction in water supply on photosynthesis, shoot development and storage root yield in sweet potato. Physiol. Plant. 2008, 134, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Lethwaite, S.L.; Triggs, C.M. Sweetpotato cultivar response to prolonged drought. N. Z. J. Agric. Res 2012, 42, 1–10. [Google Scholar]
- Kivuva, B.M. Breeding Sweetpotato (Ipomoea batatas [L.] Lam.) for Drought Tolerance in Kenya. Ph.D. Thesis, University of KwaZulu-Natal, Pietermaritzburg, South Africa, 2013. Available online: https://researchspace.ukzn.ac.za/xmlui/handle/10413/10941 (accessed on 28 February 2018).
- Solis, J.; Villordon, A.; Baisakh, N.; LaBonte, D.; Firon, N. Effect of Drought on Storage Root Development and Gene Expression Profile of Sweetpotato under Greenhouse and Field Conditions. J. Am. Soc. Hortic. Sci. 2014, 139, 317–324. [Google Scholar] [CrossRef] [Green Version]
- AOAC. Official Methods of Analysis of AOAC International, 19th ed.; Association of Official Analytical Chemists International: Rockville, MD, USA, 2011. [Google Scholar]
- Van Jaarsveld, P.; Marais, D.W.; Harmse, E.; Nestel, P.; Rodriguez-Amaya, D. Retention of β-carotene in boiled, mashed orange-fleshed sweet potato. J. Food Compos. Anal. 2006, 19, 321–329. [Google Scholar] [CrossRef]
- Low, J.W.; Van Jaarsveld, P.J. The Potential Contribution of Bread Buns Fortified with β-Carotene–Rich Sweet Potato in Central Mozambique. Food Nutr. Bull. 2008, 29, 98–107. [Google Scholar] [CrossRef] [Green Version]
- Odriozola-Serrano, I.; Hernández-Jover, T.; Martin-Belloso, O. Comparative evaluation of UV-HPLC methods and reducing agents to determine vitamin C in fruits. Food Chem. 2007, 105, 1151–1158. [Google Scholar] [CrossRef]
- Rautenbach, F.; Faber, M.; Laurie, S.; Laurie, R. Antioxidant Capacity and Antioxidant Content in Roots of 4 Sweetpotato Varieties. J. Food Sci. 2010, 75, C400–C405. [Google Scholar] [CrossRef]
- Huang, C.L.; Schulte, E.E. Digestion of plant tissue for analysis by ICP emission spectroscopy. Commun. Soil Sci. Plant Anal. 1985, 16, 943–958. [Google Scholar] [CrossRef]
- Elstner, E.F.; Heupel, A. Inhibition of nitrate formation from hydroxylammoniumchloride: A simple assay for superoxide dismutase. Anal. Biochem. 1976, 70, 616–620. [Google Scholar] [CrossRef]
- Carlberg, I.; Mannervik, B. Glutathione reductase. Methods Enzymol. 1985, 113, 484–490. [Google Scholar] [PubMed]
- Prosser, I.M.; Purves, J.V.; Saker, L.R.; Clarkson, D.T. Rapid disruption of nitrogen metabolism and nitrate transport in spinach plants deprived of sulphate. J. Exp. Bot. 2001, 52, 113–121. [Google Scholar] [CrossRef] [PubMed]
- De Ronde, J.A.; Van Der Mescht, A.; Steyn, H.S.F. Proline accumulation in response to drought and heat stress in cotton. Afr. Crop Sci. J. 2000, 8, 85–91. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar | Moisture | Energy | Protein | Total Fat | Ash | Carbohydrates | Dietary Fiber | Starch * |
|---|---|---|---|---|---|---|---|---|
| (g 100 g−1) | (kJ 100 g−1) | (%) | (g 100 g−1) | (g 100 g−1) | (%) | (g 100 g−1) | (g 100 g−1) | |
| Beauregard | 82.40 a | 266.8 d | 0.80 d | ND | 0.70 d | 13.80 e | 2.37 c | 9.57 |
| Bophelo | 76.10 bc | 355.2 c | 1.00 c | ND | 0.87 c | 17.97 d | 4.07 a | 12.87 |
| Impilo | 77.07 b | 344.2 c | 1.57 a | 0.10 ab | 0.90 bc | 16.67 d | 3.73 ab | 11.37 |
| W-119 | 72.47 de | 430.3 a | 1.20 b | 0.10 ab | 0.70 d | 21.57 ab | 3.93 a | 16.77 |
| 199062.1 | 74.30 cd | 391.7 b | 1.10 bc | 0.17 a | 0.97 ab | 19.97 c | 3.57 ab | 15.67 |
| Monate | 74.77 c | 389.2 b | 1.07 bc | ND | 0.80 c | 20.47 bc | 2.87 bc | 16.33 |
| Ndou | 71.87 e | 431.3 a | 1.53 a | ND | 0.97 a | 22.27 a | 3.37 abc | 16.43 |
| Mean | 75.57 | 372.6 | 1.180 | 0.058 | 0.842 | 18.95 | 3.41 | 14.14 |
| p-Value | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | - |
| SEM | 0.60 | 9.31 | 0.069 | 0.007 | 0.026 | 0.512 | 0.339 | - |
| LSD | 1.809 | 28.23 | 0.209 | 0.022 | 0.077 | 1.552 | 1.029 | - |
| CV% | 1.4 | 4.3 | 10.1 | 21.4 | 5.3 | 4.7 | 17.2 | - |
| Cultivar | Total Sugar | Fructose | Glucose |
|---|---|---|---|
| (g 100g−1) | (g 100g−1) | (g 100g−1) | |
| Beauregard | 4.183 a | 1.497 | 2.557 a |
| Bophelo | 3.517 abc | 1.433 | 1.730 bc |
| Impilo | 3.657 ab | 1.650 | 2.010 ab |
| W-119 | 4.020 a | 1.523 | 2.273 ab |
| 199062.1 | 2.970 bc | 1.290 | 1.683 bc |
| Monate | 3.100 bc | 1.210 | 1.693 bc |
| Ndou | 2.803 c | 1.100 | 1.293 c |
| Mean | 3.464 | 1.386 | 1.891 |
| p-Value | 0.018 | 0.638 | 0.008 |
| SEM | 0.270 | 0.228 | 0.195 |
| LSD | 0.820 | NS | 0.591 |
| CV% | 13.50 | 28.4 | 17.8 |
| Cultivar | Vitamin B9 | Vitamin B6 | β-Carotene | Vitamin C |
|---|---|---|---|---|
| mg 100g−1 | mg 100g−1 | µg 100g−1 | mg 100g−1 | |
| Beauregard | 0.00 | 0.000 c | 6510 b | 5.42 ab |
| Bophelo | 0.07 | 0.410 a | 6318 b | 5.58 a |
| Impilo | 0.00 | 0.000 c | 3837 c | 4.82 ab |
| W-119 | 0.00 | 0.000 c | 8293 a | 7.02 a |
| 199062.1 | 0.00 | 0.000 c | 2561 d | 4.94 ab |
| Monate | 0.00 | 0.000 c | 0 e | 1.45 c |
| Ndou | 0.07 | 0.313 b | 0 e | 2.32 bc |
| Mean | 0.02 | 0.103 | 3931 | 4.51 |
| p-Value | - | <0.001 | <0.001 | 0.027 |
| SEM | - | 0.158 | 236.7 | 1.050 |
| LSD | - | 0.048 | 718 | 3.184 |
| CV% | - | 26.5 | 10.4 | 40.3 |
| Cultivar | K | Cl | p | Na | Mg | Ca |
|---|---|---|---|---|---|---|
| mg 100g−1 DW | mg 100g−1 DW | mg 100g−1 DW | 100mg g−1 DW | mg 100g−1 DW | mg 100g−1 DW | |
| Beauregard | 1536 a | 678.5 | 153.1 bc | 41.4 d | 71.7 d | 54.8 bc |
| Bophelo | 1284 b | 339.8 | 167.3 b | 141.4 a | 103.9 b | 72.3 ab |
| Impilo | 1494 ab | 400.7 | 198.4 a | 97.9 bc | 136.2 a | 82.5 a |
| W-119 | 1004 c | 343.0 | 152.7 bc | 121.6 ab | 97.9 bc | 90.6 a |
| 199062.1 | 1507 a | 407.0 | 147.1 c | 40.8 d | 74.6 cd | 36.4 c |
| Monate | 1401 ab | 366.3 | 152.3 bc | 91.2 bc | 76.3 cd | 36.9 c |
| Ndou | 1450 ab | 388.3 | 157.8 bc | 69.2 cd | 87.4 bcd | 53.4 bc |
| Mean | 1383 | 374.8 | 161.2 | 86.22 | 92.58 | 61.00 |
| p-Value | 0.001 | 0.065 | 0.001 | <0.001 | <0.001 | <0.001 |
| SEM | 69.4 | 16.39 | 6.50 | 10.13 | 7.77 | 7.38 |
| LSD | 210.6 | NS | 19.71 | 30.71 | 23.56 | 22.38 |
| CV% | 8.7 | 7.6 | 7.0 | 20.3 | 14.5 | 21.0 |
| Cultivar | Zn | Mn | Fe | Cu |
|---|---|---|---|---|
| mg 100g−1 DW | mg 100g−1 DW | mg 100g−1 DW | mg 100g−1 DW | |
| Beauregard | 0.577 | 0.450 c | 1.590 bc | 0.5767 a |
| Bophelo | 0.663 | 1.357 a | 1.720 ab | 0.4500 bc |
| Impilo | 0.730 | 0.800 bc | 1.920 a | 0.4367 bc |
| W-119 | 0.690 | 0.873 b | 1.727 ab | 0.3500 d |
| 199062.1 | 0.520 | 0.720 bc | 1.400 cd | 0.3800 cd |
| Monate | 0.473 | 0.617 bc | 1.280 d | 0.3867 cd |
| Ndou | 0.653 | 0.550 bc | 1.587 bc | 0.5100 ab |
| Mean | 0.6152 | 0.767 | 1.603 | 0.4414 |
| p-Value | 0.654 | 0.007 | <0.001 | <0.001 |
| SEM | 0.075 | 0.136 | 0.067 | 0.0245 |
| LSD | ns | 0.411 | 0.205 | 0.0743 |
| CV% | 21.0 | 30.6 | 7.3 | 9.6 |
| Parameter | |||||||
|---|---|---|---|---|---|---|---|
| CCI | GR | APX | SOD | NR | Proline | ∆13C | |
| Yield | 0.521 * | −0.563 * | −0.611 * | −0.508 * | 0.764 * | −0.757 * | 0.681 * |
| Analysis | Method | Instrumentation |
|---|---|---|
| Moisture | AOAC method 950.46 | |
| Ash | AOAC method 923.03 | |
| Protein | AOAC method 992.23 | LECO nitrogen analyzer |
| Sugars | AOAC method 982.14 (amended) | GC-FID |
| Starch | AOAC method 996.11 | HPLC |
| Fat | AOAC method 996.06 | GC-FID |
| Total Dietary Fiber | AOAC method 991.43 | Enzymatic/gravimetric |
| Carbohydrates | By calculation (Carbohydrates = 100 − moisture − ash − protein − total fat) | |
| Energy | By calculation | |
| (Energy (in Kcal) = 4 × (proteins and carbohydrates mass in grams) + 9 × mass of fat in grams)(convert to kJ: 1 Kcal = 4.184 kJ) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Laurie, S.M.; Bairu, M.W.; Laurie, R.N. Analysis of the Nutritional Composition and Drought Tolerance Traits of Sweet Potato: Selection Criteria for Breeding Lines. Plants 2022, 11, 1804. https://doi.org/10.3390/plants11141804
Laurie SM, Bairu MW, Laurie RN. Analysis of the Nutritional Composition and Drought Tolerance Traits of Sweet Potato: Selection Criteria for Breeding Lines. Plants. 2022; 11(14):1804. https://doi.org/10.3390/plants11141804
Chicago/Turabian StyleLaurie, Sunette M., Michael W. Bairu, and Robert N. Laurie. 2022. "Analysis of the Nutritional Composition and Drought Tolerance Traits of Sweet Potato: Selection Criteria for Breeding Lines" Plants 11, no. 14: 1804. https://doi.org/10.3390/plants11141804