
The Genus Kalanchoe (Crassulaceae) in Ecuador: From Gardens to the Wild

 , , , ,
, , , ,  and
and
Abstract
:1. Introduction
2. Results
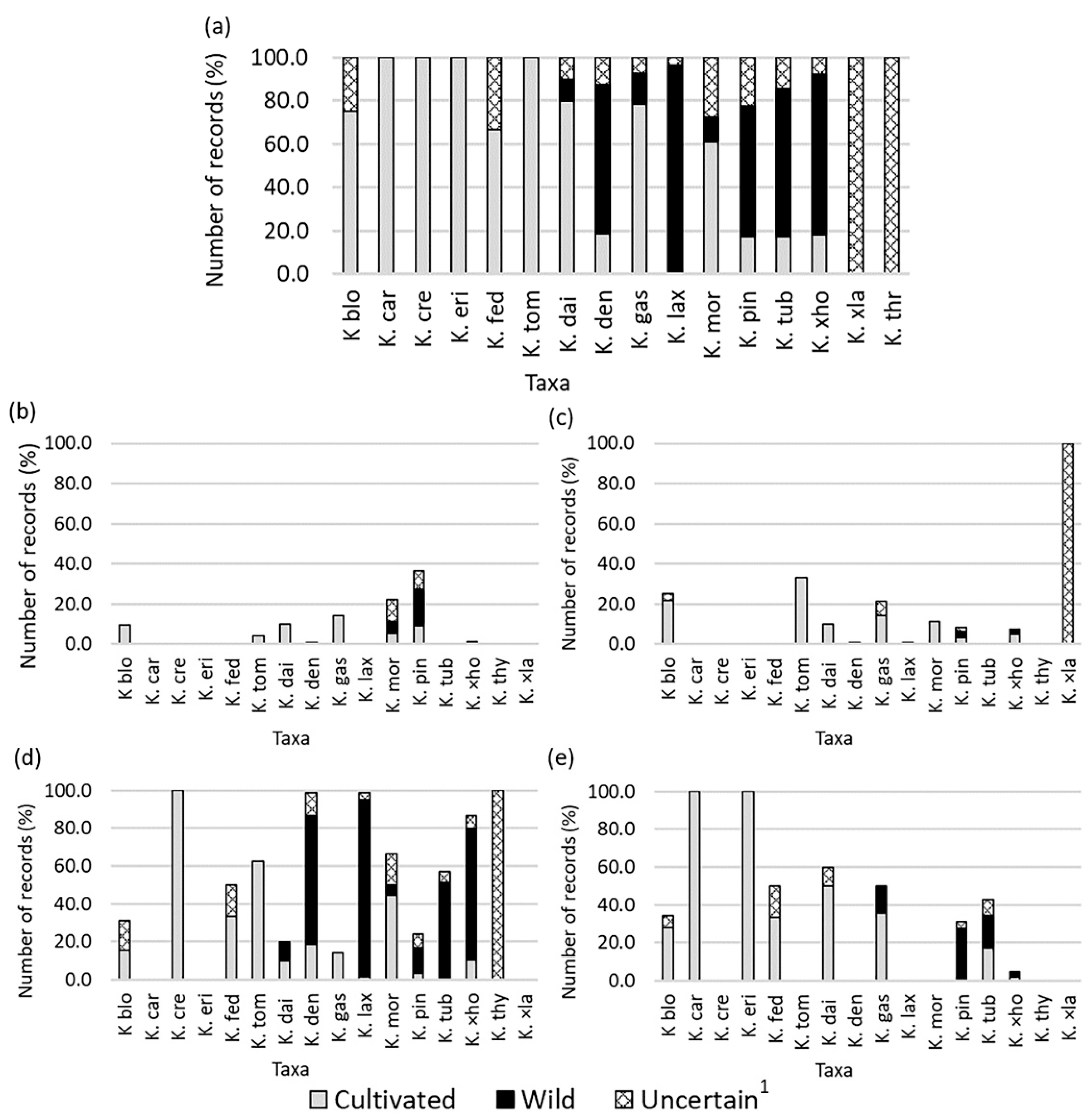
2.1. Richness of Kalanchoe Taxa in Ecuador
2.2. Key to Identify the Taxa of Kalanchoe Introduced in Ecuador Detected in the Wild with Certainty
- 1.
- Leaves pinnately compound, at least in part………………………..………..K. pinnata
- 2.
- Leaves always simple
- 2.1.
- Leaf blade suborbicular, ovate, or elliptic (length/width ratio between 1:1 and 2:1)
- 2.1.1.
- Plants 0.3–1(3) m. Green leaves without spots, finely crenate. Inflorescence of erect flowers. The only taxon of the group under study that does not produce bulbils on the leaves……………………………………..K. densiflora
- 2.1.2.
- Plants up to 0.5 m. Green or somewhat greyish leaves, sometimes with scattered brown spots and a reddish line on the edge, which may have small auricles at the base, with a crenate margin with few lobes. Inflorescence of pendulous flowers…………………………………...…K. laxiflora
- 2.2.
- Leaf blade narrowly ovate, oblong, subdeltoid, linear, or subcylindrical (ratio 3:1 or greater)
- 2.2.1.
- Plants with narrowly ovate, oblong, or subcylindrical leaves, crenate, with spots on the reverse or without spots, which form bulbils only at the apex
- 2.2.1.1.
- Narrowly ovate to oblong leaves
- 2.2.1.1.1.
- Green or bluish-green leaves generally without spots…..K. mortagei
- 2.2.1.1.2.
- More or less light green leaves, with irregular spots………………………………………………...……..K. gastonis-bonnieri
- 2.2.1.2.
- Subcylindrical leaves…………………………………………...K. tubiflora
- 2.2.2.
- Plants with narrowly ovate, oblong, or subdeltoid leaves, toothed, with spots on the reverse, which form bulbils along almost the entire leaf margin
- 2.2.2.1.
- Leaves subdeltoid or triangular, the largest subpeltate with the base forming a very conspicuous fold perpendicular to the blade………………………………………………………...K. daigremontiana
- 2.2.1.2.
- Leaves variable in shape and size, from narrowly lanceolate with a decurrent base towards the petiole to non-decurrent subdeltoid, which do not form a basal fold………………………………………..K. ×houghtonii
2.3. Temporal Accumulation and Distribution of Kalanchoe Records
2.4. Kalanchoe Taxa Categorized
2.5. Environmental Preferences
3. Discussion
3.1. Kalanchoe Taxa in Ecuador
3.2. Suggestions for Management
4. Materials and Methods
4.1. Study Area
4.2. List of Kalanchoe Taxa and Occurrences
4.3. Spatial–Temporal Distribution of the Records of Kalanchoe Taxa
4.4. Categorization of Kalanchoe Taxa by Invasion Status
4.5. Environmental Preferences
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Richardson, D.M.; Py, P.; Rejmánek, M.; Barbour, M.G.; Panetta, F.D.; West, C.J. Naturalization and Invasion of Alien Plants: Concepts and Definitions. Divers. Distrib. 2000, 15, 93–107. [Google Scholar] [CrossRef]
- Essl, F.; Bacher, S.; Genovesi, P.; Hulme, P.E.; Jeschke, J.M.; Katsanevakis, S.; Kowarik, I.; Kühn, I.; Pyšek, P.; Rabitsch, W.; et al. Which Taxa are Alien? Criteria, Applications, and Uncertainties. BioScience 2018, 68, 496–509. [Google Scholar] [CrossRef]
- Sharma, G.P.; Singh, J.S.; Raghubanshi, A.S. Plant Invasions: Emerging Trends and Future Implications. Curr. Sci. 2005, 88, 726–734. [Google Scholar]
- Olenin, S.; Gollasch, S.; Lehtiniemi, M.; Sapota, M.; Zaiko, A. Biological Invasions. In Biological Oceanography of the Baltic Sea; Snoeijs-Leijonmalm, P., Schubert, H., Radziejewska, T., Eds.; Springer: Dordrecht, The Netherlands, 2017; pp. 193–232. [Google Scholar]
- Alston, K.P.; Richardson, D.M. The Roles of Habitat Features, Disturbance, and Distance from Putative Source Populations in Structuring Alien Plant Invasions at the Urban/Wildland Interface on the Cape Peninsula, South Africa. Biol. Conserv. 2006, 132, 183–198. [Google Scholar] [CrossRef]
- Meyerson, L.A.; Mooney, H.A. Invasive Alien Species in an Era of Globalization. Front. Ecol. Environ. 2007, 5, 199–208. [Google Scholar] [CrossRef]
- Seebens, H.; Essl, F.; Dawson, W.; Fuentes, N.; Moser, D.; Pergl, J.; Pyšek, P.; van Kleunen, M.; Weber, E.; Winter, M.; et al. Global Trade will Accelerate Plant Invasions in Emerging Economies under Climate Change. Glob. Chang. Biol. 2015, 21, 4128–4140. [Google Scholar] [CrossRef]
- Dehnen-Schmutz, K.; Touza, J. Plant Invasions and Ornamental Horticulture: Pathway, Propagule Pressure and the Legal Framework. In Floriculture, Ornamental and Plant Biotechnology; Global Science Books: Isleworth, UK, 2008; Volume 5, pp. 15–21. [Google Scholar]
- Lawson, R.H. Economic Importance and Trends in Ornamental Horticulture. Acta Hortic. 1996, 432, 226–237. [Google Scholar] [CrossRef]
- Hulme, P.E.; Brundu, G.; Carboni, M.; Dehnen-Schmutz, K.; Dullinger, S.; Early, R.; Essl, F.; González-Moreno, P.; Groom, Q.J.; Kueffer, C.; et al. Integrating Invasive Species Policies across Ornamental Horticulture Supply Chains to Prevent Plant Invasions. J. Appl. Ecol. 2018, 55, 92–98. [Google Scholar] [CrossRef]
- van Kleunen, M.; Essl, F.; Pergl, J.; Brundu, G.; Carboni, M.; Dullinger, S.; Early, R.; González-Moreno, P.; Groom, Q.J.; Hulme, P.E.; et al. The Changing Role of Ornamental Horticulture in Alien Plant Invasions: Horticulture and Plant Invasions. Biol. Rev. 2018, 93, 1421–1437. [Google Scholar] [CrossRef]
- Kühn, I.; Klotz, S. Floristischer Status und Gebietsfremde Arten. In BIOLFLOR: Eine Datenbank mit Biologisch-Ökologischen Merkmalen zur Flora von Deutschland; Schriftenreihe für Vegetationskunde 38, Bundesamt fur Naturschutz: Bonn, Germany, 2002; pp. 47–56. [Google Scholar]
- Pyšek, P.; Sádlo, J.; Mandák, B. Catalogue of Alien Plants of the Czech Republic. Preslia 2002, 74, 97–186. [Google Scholar]
- Mack, R.N.; Erneberg, M. The United States Naturalized Flora: Largely the Product of Deliberate Introductions. Ann. Mo. Bot. Gard. 2002, 89, 176–189. [Google Scholar] [CrossRef]
- Zhang, A.; Hu, X.; Yao, S.; Yu, M.; Ying, Z. Alien, Naturalized and Invasive Plants in China. Plants 2021, 10, 2241. [Google Scholar] [CrossRef] [PubMed]
- Smith, G. Kalanchoe Species Poisoning in Pets. Toxicology Brief. Vet. Med. 2004, 1, 933–936. [Google Scholar]
- Foxcroft, L.C.; Richardson, D.M.; Wilson, J.R.U. Ornamental Plants as Invasive Aliens: Problems and Solutions in Kruger National Park, South Africa. Environ. Manag. 2008, 41, 32–51. [Google Scholar] [CrossRef]
- Hurrell, J.A.; Delucchi, G.; Keller, H.A.; Stampella, P.C.; Guerrero, E.L. Bryophyllum (Crassulaceae): Especies Ornamentales Naturalizadas en la Argentina. Bonplandia 2012, 21, 169–181. [Google Scholar] [CrossRef]
- Hannan-Jones, M.A.; Playford, J. The Biology of Australian Weeds 40. Bryophyllum Salisb. Species. Plant Prot. Q. 2002, 17, 42–57. [Google Scholar]
- Akulova-Barlow, Z. Kalanchoe. Cact. Succ. J. 2009, 81, 268–276. [Google Scholar] [CrossRef]
- Descoings, B. Kalanchoe. In Illustrated Handbook of Succulent Plants: Crassulaceae; Eggli, U., Ed.; Springer: Berlin, Germany, 2003; pp. 143–181. [Google Scholar]
- Baptiste, M.P.; Castaño, N.; Cárdenas López, D.; de Gutiérrez, F.P.; Gil, D.; Lasso, C. Análisis de Riesgo y Propuesta de Categorización de Especies Introducidas para Colombia; Instituto de Investigación en Recursos Biológicos Alexander von Humboldt: Bogotá, Colombia, 2010. [Google Scholar]
- Herrera, I.; Nassar, J.M. Reproductive and Recruitment Traits as Indicators of the Invasive Potential of Kalanchoe daigremontiana (Crassulaceae) and Stapelia gigantea (Apocynaceae) in a Neotropical Arid Zone. J. Arid. Environ. 2009, 73, 978–986. [Google Scholar] [CrossRef]
- Guerra-García, A.; Barrales-Alcalá, D.; Argueta-Guzmán, M.; Cruz, A.; Mandujano, M.C.; Arévalo-Ramírez, J.A.; Milligan, B.G.; Golubov, J. Biomass Allocation, Plantlet Survival, and Chemical Control of the Invasive Chandelier Plant Kalanchoe delagoensis (Crassulaceae). Invasive Plant Sci. Manag. 2018, 11, 33–39. [Google Scholar] [CrossRef]
- Nassis, C.Z.; Haebisch, E.M.; Giesbrecht, A.M. Antihistamine Activity of Bryophyllum calycinum. Braz. J. Med. Biol. Res. 1992, 25, 929–936. [Google Scholar]
- Ojewole, J.A.O. Antinociceptive, Anti-Inflammatory and Antidiabetic Effects of Bryophyllum pinnatum (Crassulaceae) Leaf Aqueous Extract. J. Ethnopharmacol. 2005, 99, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Nayak, B.S.; Marshall, J.R.; Isitor, G. Wound Healing Potential of Ethanolic Extract of Kalanchoe pinnata Lam. Leaf—A Preliminary Study. Indian J. Exp. Biol. 2010, 48, 572–576. [Google Scholar] [PubMed]
- Kolodziejczyk-Czepas, J.; Stochmal, A. Bufadienolides of Kalanchoe Species: An Overview of Chemical Structure, Biological Activity and Prospects for Pharmacological Use. Phytochem. Rev. 2017, 16, 1155–1171. [Google Scholar] [CrossRef] [PubMed]
- Stefanowicz-Hajduk, J.; Hering, A.; Gucwa, M.; Hałasa, R.; Soluch, A.; Kowalczyk, M.; Stochmal, A.; Ochocka, R. Biological Activities of Leaf Extracts from Selected Kalanchoe Species and their Relationship with Bufadienolides Content. Pharm. Biol. 2020, 58, 732–740. [Google Scholar] [CrossRef]
- Saravanan, V.; Murugan, S.S.; Kumaravel, T.S. Genotoxicity Studies with an Ethanolic Extract of Kalanchoe pinnata Leaves. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2020, 856–857, 503229. [Google Scholar] [CrossRef] [PubMed]
- McKenzie, R.A.; Dunster, P.J. Hearts and Flowers: Bryophyllum Poisoning of Cattle. Aust. Vet. J. 1986, 63, 222–227. [Google Scholar] [CrossRef]
- Botha, C.J.; Penrith, M.-L. Potential Plant Poisonings in Dogs and Cats in Southern Africa. J. S. Afr. Vet. Assoc. 2009, 80, 63–74. [Google Scholar] [CrossRef]
- Mendonça, F.S.; Nascimento, N.C.F.; Almeida, V.M.; Braga, T.C.; Ribeiro, D.P.; Chaves, H.A.S.; Silva Filho, G.B.; Riet-Correa, F. An Outbreak of Poisoning by Kalanchoe blossfeldiana in Cattle in Northeastern Brazil. Trop. Anim. Health Prod. 2018, 50, 693–696. [Google Scholar] [CrossRef]
- Randall, R.P. The Introduced Flora of Australia and Its Weed Status; Cooperative Research Centre for Australian Weed Management: Glen Osmond, Australia, 2007. [Google Scholar]
- González Gutiérrez, P.A.; López-Pujol, J.; Gómez-Bellver, C. Notas sobre las Especies de Kalanchoe (Crassulaceae) Ocasionales y Naturalizadas en Cuba. Collect. Bot. 2019, 38, e011. [Google Scholar] [CrossRef]
- Armstrong, T.R.; Swarbrick, J.T. Bryophyllum spp. in Queensland; Wilson, B.J., Swarbrick, J.T., Eds.; Council of Australian Weed Science Sociaties: Queensland, Australia, 1981; p. 211. [Google Scholar]
- Moran, R.V. Crassulaceae. In Flora of North America North of Mexico; Flora of North America Editorial Committee, Ed.; Oxford University Press: New York, NY, USA, 2009; Volume 8, pp. 147–230. [Google Scholar]
- Staples, G.W.; Herbst, D.; Imada, C.T. Survey of Invasive or Potentially Invasive Cultivated Plants in Hawaii. Bishop Mus. Occas. Pap. 2000, 31, 33. [Google Scholar]
- Herrando-Moraira, S.; Vitales, D.; Nualart, N.; Gómez-Bellver, C.; Ibáñez, N.; Massó, S.; Cachón-Ferrero, P.; González-Gutiérrez, P.A.; Guillot, D.; Herrera, I.; et al. Global Distribution Patterns and Niche Modelling of the Invasive Kalanchoe × houghtonii (Crassulaceae). Sci. Rep. 2020, 10, 3143. [Google Scholar] [CrossRef] [PubMed]
- Boiteau, P.; Allorge-Boiteau, L. Kalanchoe de Madagascar. In Systématique, Écophysiologie et Phytochemie; Karthala: Paris, France, 1995. [Google Scholar]
- González de León, S. Distribución Potencial de la Especie Invasora Kalanchoe pinnata ¿Un ejemplo de Expansión de Nicho? BioInvasiones 2015, 5, 10–17. [Google Scholar]
- Herrera, I.; Espinoza, F.; Solas, S.A.; Laporta, M.G.; Ruiz, E.M.; Rodriguez, R.; Tejena, N.; Narváez, S.; Ferri, M.; Pagad, S. GRIIS Checklist of Introduced and Invasive Species—Ecuador. Version 1.7. Invasive Species Specialist Group ISSG. Checklist Dataset. 2021. Available online: https://www.gbif.org/es/dataset/1df9467e-0c60-4e5a-a7b0-2b60bd6648ed (accessed on 10 January 2022).
- Guézou, A.; Trueman, M.; Buddenhagen, C.E.; Chamorro, S.; Guerrero, A.M.; Pozo, P.; Atkinson, R. An Extensive Alien Plant Inventory from the Inhabited Areas of Galapagos. PLoS ONE 2010, 5, e10276. [Google Scholar] [CrossRef] [PubMed]
- Jaramillo, P.; Guézou, A.; Mauchamp, A.; Tye, A. CDF Checklist of Galapagos Flowering Plants. In Charles Darwin Foundation Galapagos Species Checklist; Fundación Charles Darwin: Puerto Ayora, Ecuador, 2017; pp. 1–320. [Google Scholar]
- Causton, C.; Jäger, H.; Jiménez-Uzcátegui, G.; Keith, I.; Wong, L.J.; Pagad, S. Global Register of Introduced and Invasive Species–Galápagos Islands, Ecuador, Version 1.5. Invasive Species Specialist Group ISSG. Checklist Dataset; 2020. Available online: https://www.gbif.org/es/dataset/2a25bba9-eb59-461e-b458-a4a609b5154e (accessed on 10 January 2022).
- Tye, A. Invasive Plant Problems and Requirements for Weed Risk Assessment in the Galapagos Islands. In Weed Risk Assessment; Groves, R.H., Panetta, F.D., Virtue, J.G., Eds.; CSIRO Publishing: Collingwood, Australia, 2001; pp. 153–175. [Google Scholar]
- Freire, A. Crassulaceae. In Flora of Ecuador; Harling, C., Andersson, L., Eds.; Botanical Institute, University of Göteborg: Göteborg, Sweden, 2004; Volume 73, pp. 3–16. [Google Scholar]
- Neill, A.K.; Kieschnick, S.R. Noccaea perfoliata or Microthlaspi perfoliatum (Brassicaceae), New to the Flora of Texas, U.S.A. J. Bot. Res. Inst. Tex. 2021, 15, 309–315. [Google Scholar] [CrossRef]
- Batianoff, G.N.; Franks, A.J. Invasion of Sandy Beachfronts by Ornamental Plant Species in Queensland. Plant Prot. Q. 1997, 12, 180–186. [Google Scholar]
- Mackenzie, K.K.; Lütken, H.; Coelho, L.L.; Kaaber, M.D.; Hegelund, J.N.; Müller, R. Kalanchoë. In Ornamental Crops; Van Huylenbroeck, J., Ed.; Handbook of Plant Breeding; Springer International Publishing: Cham, Switzerland, 2018; Volume 11, pp. 453–479. [Google Scholar]
- Reichard, S.H.; White, P. Horticulture as a Pathway of Invasive Plant Introductions in the United States. BioScience 2001, 51, 103–113. [Google Scholar] [CrossRef]
- Bradley, B.A.; Blumenthal, D.M.; Early, R.; Grosholz, E.D.; Lawler, J.J.; Miller, L.P.; Sorte, C.J.; D’Antonio, C.M.; Diez, J.M.; Dukes, J.S.; et al. Global Change, Global Trade, and the next Wave of Plant Invasions. Front. Ecol. Environ. 2012, 10, 20–28. [Google Scholar] [CrossRef]
- Arianoutsou, M.; Bazos, I.; Christopoulou, A.; Kokkoris, Y.; Zikos, A.; Zervou, S.; Delipetrou, P.; Cardoso, A.C.; Deriu, I.; Gervasini, E.; et al. Alien Plants of Europe: Introduction Pathways, Gateways and Time Trends. PeerJ 2021, 9, e11270. [Google Scholar] [CrossRef]
- McGrannachan, C.M.; Pagad, S.; McGeoch, M.A. A Multiregional Assessment of Transnational Pathways of Introduction. NeoBiota 2021, 64, 43–67. [Google Scholar] [CrossRef]
- Milad, R. Genus Kalanchoe (Crassulaceae): A Review of Its Ethnomedicinal, Botanical, Chemical and Pharmacological Properties. Eur. J. Med. Plants 2014, 4, 86–104. [Google Scholar] [CrossRef]
- Quazi Majaz, A.; Tatiya, A.U.; Khurshid, M.; Nazim, S.; Siraj, S. The Miracle Plant (Kalanchoe pinnata): A Phytochemical and Pharmacological Review. Int. J. Res. Ayurveda Pharm. 2011, 2, 1478–1482. [Google Scholar]
- da Silva Ferreira, R.G.; de Souza Fernandes, N.; da Veiga-Junior, V.F. Kalanchoe brasiliensis Camb. and Kalanchoe pinnata (Lamk.) Pers. In Medicinal and Aromatic Plants of South America; Albuquerque, U.P., Patil, U., Máthé, Á., Eds.; Medicinal and Aromatic Plants of the World; Springer: Dordrecht, The Netherlands, 2018; Volume 5, pp. 265–273. [Google Scholar]
- Paredes, D.J.; Allauca, M.P.B.; Rodríguez, N.J.M. Usos de plantas medicinales en la comunidad San Jacinto del Cantón Ventanas, Los Ríos—Ecuador. Rev. UDCA Actual. Divulg. Cient. 2015, 18, 39–50. [Google Scholar]
- Muñoz Martínez, A.S.; Monar, M.; Ugsha Jimenez, Y.E.; Cabrera Valarezo, J.V.; Puma Vaque, L.H.; Ríos, M. Etnofarmacología de Kalanchoe pinnata en Amazonía: Uso Medicinal de “Chugri Yuyu”. Med. Plant Commun. 2019, 2, 233. [Google Scholar]
- Zambrano-Intriago, L.F.; Buenaño-Allauca, M.P.; Mancera-Rodríguez, N.J.; Jiménez-Romero, E. Estudio Etnobotánico de Plantas Medicinales Utilizadas por los Habitantes del Área Rural de la Parroquia San Carlos, Quevedo, Ecuador. Univ. Salud 2015, 17, 97–111. [Google Scholar]
- Murillo López, D. Inventario de la Biodiversidad de Plantas Medicinales en el Área Rural del Sur de Manabi. Bachelor’s Thesis, Universidad Estatal del Sur de Manabí, Jipijapa, Ecuador, 2018. [Google Scholar]
- Duque Gualpa, M.B. Etnoecología de las Plantas Medicinales en las Chacras Familiares de la Comunidad Fakcha Llakta, Cantón Otavalo, Provincia de Imbabura. Bachelor’s Thesis, Universidad Técnica del Norte, Otavalo, Ecuador, 2018. [Google Scholar]
- Rivero-Guerra, A.O. Uso Tradicional de Especies de Plantas en Trece Provincias de Ecuador. Collect. Bot. 2021, 40, e002. [Google Scholar] [CrossRef]
- Aguilar Valverde, W.D.; Bastidas Sánchez, C.J. Procesamiento de la Hoja de Kalanchoe Gastonis-Bonnieri y Aloe vera, para Obtener una Bebida Nutracética. Bachelor’s Thesis, Universidad de Guayaquil, Guayaquil, Ecuador, 2016. [Google Scholar]
- Pinela Castro, D.L.; Zambrano Alarcón, M.E.; Arreaga Pantaleón, K.A. Formulación y Caracterización de una Bebida Antioxidante y Refrescante a Base de la Planta Dulcamara: Bryophyllum gastonis-bonnieri. RECIAMUC 2017, 1, 37–56. [Google Scholar]
- Guamushig, F.N.G. Etapa de Crecimiento y Engorde en Pollos Broiler en el Barrio Satán del Cantón Latacunga. Bachelor’s Thesis, Universidad Técnica de Cotopaxi, Latacunga, Ecuador, 2016. [Google Scholar]
- Concha, V.; Negrete, J.; Toapanta, S.; Condo, L.; Wamputsrik, E. Cálculo en la Dosificación de Dulcamara Kalanchoe gastonis-bonnieri sobre Parámetros Productivos en Pollos en Morona Santiago, Ecuador. Perfiles 2021, 25, 30–36. [Google Scholar] [CrossRef]
- Otto, R.; Verloove, F. Nuevos Xenófitos de la Palma (Islas Canarias, España), con Énfasis en las Especies Naturalizadas y (Potencialmente) Invasoras. Collect. Bot. 2016, 35, e001. [Google Scholar] [CrossRef]
- Fabricante, J.R.; Targino de Araújo, K.C.; Almeida, T.S.; Bispo Santos, J.P.; Oliveira Reis, D. Invasive Alien Plants in Sergipe, North-Eastern Brazil. Neotrop. Biol. Conserv. 2021, 16, 89–104. [Google Scholar] [CrossRef]
- Nelson, C.H. Adiciones y Comentarios a la Flora de Honduras. Ceiba 2010, 51, 70–88. [Google Scholar] [CrossRef]
- Guillot, D. Apuntes Corológicos sobre Neófitos de la Flora Valenciana. Flora Montiberica 2003, 23, 13–17. [Google Scholar]
- Smith, G.F.; Figueiredo, E.; Van Wyk, A.E. Kalanchoe (Crassulaceae) in Southern Africa: Classification, Biology, and Cultivation; Academic Press: London, UK, 2019. [Google Scholar]
- Stanley, T.D.; Ross, E.M. Flora of South-Eastern Queensland; Queensland Deparment of Primary Industries: Brisbane, Australia, 1986; Volume 2. [Google Scholar]
- Xu, H.; Qiang, S.; Genovesi, P.; Ding, H.; Wu, J.; Meng, L.; Han, Z.; Miao, J.; Hu, B.; Guo, J.; et al. An Inventory of Invasive Alien Species in China. NeoBiota 2012, 15, 1–26. [Google Scholar] [CrossRef]
- US Forest Service. Kalanchoe pinnata: Info from PIER (PIER Species Info). Available online: http://www.hear.org/pier/species/kalanchoe_pinnata.htm (accessed on 12 May 2022).
- González de León, S.; Herrera, I.; Guevara, R. Mating System, Population Growth, and Management Scenario for Kalanchoe pinnata in an Invaded Seasonally Dry Tropical Forest. Ecol. Evol. 2016, 6, 4541–4550. [Google Scholar] [CrossRef] [PubMed]
- Witt, A.; Luke, Q. Guide to the Naturalized and Invasive Plants of Eastern Africa; CABI: Wallingford, UK, 2017. [Google Scholar]
- Oviedo Prieto, R.; Herrera, P.; Caluff, M.G.; Regalado, L.; Ventosa, I.; Plasencia, J.M.; Baró, I.; Camacho, J.P.; Schwesinger, H.; González-Oliva, L.; et al. Lista Nacional de Especies de Plantas Invasoras y Potencialmente Invasoras en la República de Cuba 2011. Bissea 2012, 6, 22–112. [Google Scholar]
- Schofield, E.K. Effects of Introduced Plants and Animals on Island Vegetation: Examples from Galapagos Archipelago. Conserv. Biol. 1989, 3, 227–239. [Google Scholar] [CrossRef]
- Mauchamp, A. Threats from Alien Plant Species in the Galápagos Islands. Conserv. Biol. 1997, 11, 260–263. [Google Scholar] [CrossRef]
- Gardener, M.R.; Tye, A.; Wilkinson, S.R. Control of Introduced Plants in the Galapagos Islands. In Proceedings from Twelth Australian Weeds Conference; Bishop, A.C., Boersma, M., Barnes, C.D., Eds.; University of Tasmania: Hobart, Australia, 1999; pp. 396–400. [Google Scholar]
- Schofield, E.K. Galápagos Flora: The Threat of Introduced Plants. Biol. Conserv. 1973, 5, 48–51. [Google Scholar] [CrossRef]
- Guevara, R.; Ruiz-Guerra, B.; Aguilar-Chama, A.; Velázquez-Rosas, N.; Sevillano García-Mayeya, L.; De Los Santos, M. Estado Actual de las Poblaciones de Bryophyllum pinnatum y su Impacto en la Regeneración de Plantas Nativas de la Selva Baja Caducifolia que Crece cn Mal País, en la Planicie Costera Veracruzana; Final Report of Proyect- N°LI022 Conabio: Ciudad de México, México, 2017. [Google Scholar]
- Nazir, T.; Uniyal, A.K.; Ahmed, M. Allelopathic Response of Medicinal Plants on Germination and Growth of Traditional Field Crops. Indian For. 2014, 140, 695–700. [Google Scholar]
- Cruz Montilla, J. Estado de Transformación y Fisonomía de una Relicto de Enclave Subxerofitico en el Suroccidente de la Sabana de Bogotá. Bachelor’s Thesis, Pontificia Universidad Javeriana, Bogotá, Colombia, 2018. [Google Scholar]
- Sandoval Pulgarín, L.M. Patrones y Procesos de Invasión de Plantas en los Alrededores del Macizo de Iguaque (Boyacá, Colombia). Bachelor’s Thesis, Universidad Nacional de Colombia, Bogotá, Colombia, 2020. [Google Scholar]
- Mesquida, V.; Gómez-Bellver, C.; Guillot, D.; Herrando-Moraira, S.; Nualart, N.; Sáez, L.; López Pujol, J. El Gènere Kalanchoe (Crassulaceae) a Catalunya: Situació i Distribució Potencial del Tàxon Invasor K. × houghtonii. Orsis 2017, 31, 37–64. [Google Scholar] [CrossRef]
- Shtein, R.; Smith, G.F. A Revision of the Climbing Kalanchoes (Crassulaceae Subfam. Kalanchooideae) of Madagascar Including the Description of Kalanchoe sect. Invasores and K. ser. Vilana. Phytotaxa 2021, 482, 93–120. [Google Scholar] [CrossRef]
- Witt, A.B.R.; Nongogo, A.X. The Impact of Fire, and its Potential Role in Limiting the Distribution of Bryophyllum delagoense (Crassulaceae) in Southern Africa. Biol. Invasions 2011, 13, 125–133. [Google Scholar] [CrossRef]
- Bär, W.; Pfeifer, P.; Dettner, K. Intra-and Interspecific Allelochemical Effects in Three Kalanchoe-Species (Crassulaceae). Z. Naturforsch. C. J. Biosci. 1997, 52, 441–449. [Google Scholar] [CrossRef]
- Groner, M.G. Allelopathic Influence of Kalanchoe daigremontiana on other Species of Plants. Bot. Gaz. 1975, 136, 207–211. [Google Scholar] [CrossRef]
- Herrera, I.; Ferrer-Paris, J.R.; Hernández-Rosas, J.I.; Nassar, J.M. Impact of Two Invasive Succulents on Native-Seedling Recruitment in Neotropical Arid Environments. J. Arid. Environ. 2016, 132, 15–25. [Google Scholar] [CrossRef]
- Herrera, I.; Ferrer-Paris, J.R.; Benzo, D.; Flores, S.; García, B.; Nassar, J.M. An Invasive Succulent Plant (Kalanchoe daigremontiana) Influences Soil Carbon and Nitrogen Mineralization in a Neotropical Semiarid Zone. Pedosphere 2018, 28, 632–643. [Google Scholar] [CrossRef]
- Smith, G.F.; Shtein, R.; Klein, D.-P.; Parihar, B.; Almeida, A.; Rodewald, S.; Kadereit, G. Sexual and Asexual Reproduction in Kalanchoe (Crassulaceae): A Review of Known and Newly Recorded Strategies. Haseltonia 2022, 28, 2–20. [Google Scholar] [CrossRef]
- Witt, A.B.R. Initial Screening of the Stem-Boring Weevil Osphilia tenuipes, a Candidate Agent for the Biological Control of Bryophyllum delagoense in Australia. BioControl 2004, 49, 197–209. [Google Scholar] [CrossRef]
- Witt, A.B.R.; McConnachie, A.J.; Stals, R. Alcidodes sedi (Col.: Curculionidae), a Natural Enemy of Bryophyllum delagoense (Crassulaceae) in South Africa and a Possible Candidate Agent for the Biological Control of this Weed in Australia. Biol. Control 2004, 31, 380–387. [Google Scholar] [CrossRef]
- Essl, F.; Nehring, S.; Klingenstein, F.; Milasowszky, N.; Nowack, C.; Rabitsch, W. Review of Risk Assessment Systems of IAS in Europe and Introducing the German–Austrian Black List Information System (GABLIS). J. Nat. Conserv. 2011, 19, 339–350. [Google Scholar] [CrossRef]
- Faulkner, K.T.; Robertson, M.P.; Rouget, M.; Wilson, J.R.U. A Simple, Rapid Methodology for Developing Invasive Species Watch Lists. Biol. Conserv. 2014, 179, 25–32. [Google Scholar] [CrossRef]
- Pergl, J.; Sádlo, J.; Petrusek, A.; Laštůvka, Z.; Musil, J.; Perglová, I.; Šanda, R.; Šefrová, H.; Šíma, J.; Vohralík, V.; et al. Black, Grey and Watch Lists of Alien Species in the Czech Republic Based on Environmental Impacts and Management Strategy. NeoBiota 2016, 28, 1–37. [Google Scholar] [CrossRef]
- Genovesi, P.; Shine, C. Stratégie Européenne Relative Aux Espèces Exotiques Envahissantes: Convention Relative à la Conservation de la Vie Sauvage et Du Milieu Naturel de l’Europe (Convention de Berne); Sauvegarde de la nature n°137; Editions du Conseil de l’Europe: Strasbourg, France, 2004. [Google Scholar]
- Ulloa Ulloa, C.; Acevedo-Rodríguez, P.; Beck, S.; Belgrano, M.J.; Bernal, R.; Berry, P.E.; Brako, L.; Celis, M.; Davidse, G.; Forzza, R.C.; et al. An Integrated Assessment of the Vascular Plant Species of the Americas. Science 2017, 358, 1614–1617. [Google Scholar] [CrossRef] [PubMed]
- Shtein, R.; Smith, G.F.; Ikeda, J. Aspects of the Taxonomy of the Kalanchoe daigremontiana Species Complex (Crassulaceae Subfam. Kalanchooideae) and Associated Interspecific Hybrids in Southern Madagascar, with the Description of a New Nothospecies, K. × descoingsii (=K. laetivirens × K. tubiflora). Phytotaxa 2021, 524, 235–260. [Google Scholar]
- Berger, P. Crassulaceae. Unterfam. II. Kalanchoideae [sic Kalanchooideae]. 6. Kalanchoe. In Die natürlichen Pflanzenfamilien; Engler, A., Prantl, K., Eds.; Verlag von Wilhelm Engelmann: Leipzig, Germany, 1935; pp. 402–412. [Google Scholar]
- Fu, K.; Ohba, H. Crassulaceae. In Flora of China; Wu, Z.Y., Raven, P.H., Eds.; Science Press: Beijing, China; Botanical Garden Press: St. Louis, MO, USA, 2001; Volume 8, pp. 202–268. [Google Scholar]
- Chernetskyy, M. Problems in Nomenclature and Systematics in the Subfamily Kalanchoideae (Crassulaceae) over the Years. Acta Agrobot. 2011, 64, 67–74. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 15 February 2022).
- UNEP-WCMC. IUCN Protected Planet—Ecuador. Available online: https://www.protectedplanet.net/country/ECU (accessed on 10 May 2022).
- UNESCO. Geospatial information System (GIS), Network and Library. Estructura_Suelo_Urbano_Rural. Available online: https://galapagos-geonode.unesco.org/#/search/?q=Estructura_suelo_urbano_rural_21_12_21 (accessed on 13 May 2022).
- Pyšek, P.; Richardson, D.M.; Rejmánek, M.; Webster, G.L.; Williamson, M.; Kirschner, J. Alien Plants in Checklists and Floras: Towards Better Communication between Taxonomists and Ecologists. Taxon 2004, 53, 131–143. [Google Scholar] [CrossRef]
- Sanderson, E.W.; Jaiteh, M.; Levy, M.A.; Redford, K.H.; Wannebo, A.V.; Woolmer, G. The Human Footprint and the Last of the Wild: The Human Footprint is a Global Map of Human Influence on the Land Surface, which Suggests that Human Beings are Stewards of Nature, Whether we like It or Not. BioScience 2002, 52, 891–904. [Google Scholar] [CrossRef]
- Ellis, E.C.; Goldewijk, K.K.; Siebert, S.; Lightman, D.; Ramankutty, N. Anthropogenic Biomes of the World, Version 2: 2000; NASA Socioeconomic Data and Applications Center (SEDAC): New York, NY, USA, 2013.
- Racine, J.S. RStudio: A Platform-Independent IDE for R and Sweave. J. Appl. Econ. 2012, 27, 167–172. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Kalanchoe Taxa | Amazonian | Andean | Coastal | Insular | Total (%) |
|---|---|---|---|---|---|
| K. blossfeldiana Poelln. | 3 (0/3) | 10 (0/5) | 8 (0/7) | 11 (0/9) | 32 (3.3%) |
| K. carnea N.E. Br. | 0 | 0 | 0 | 1 (0/1) | 1 (0.1%) |
| K. crenata (Andrews) Haw. | 0 | 5 (0/5) | 0 | 0 | 5 (0.5%) |
| K. daigremontiana Raym.-Hamet and H. Perrier | 1 (0/1) | 2 (1/1) | 1 (0/1) | 6 (0/5) | 10 (1.0%) |
| K. densiflora Rolfe | 1 (1/0) | 180 (124/ 34) | 1 | 0 | 182 (18.8%) |
| K. eriophylla Hils. and Bojer ex Tul. | 0 | 0 | 0 | 1 (0/1) | 1 (0.1%) |
| K. fedtschenkoi Raym.-Hamet and H. Perrier | 0 | 3 (0/2) | 0 | 3 (0/2) | 6 (0.6%) |
| K. gastonis-bonnieri Raym.-Hamet and H. Perrier | 2 (0/2) | 2 (0/2) | 3 (0/2) | 7 (2/5) | 14 (1.4%) |
| K. laxiflora Baker | 1 (1/0) | 277 (263/4) | 2 | 0 | 280 (29.0%) |
| K. mortagei Raym.-Hamet and H.Perrier | 4 (1/1) | 12 (1/8) | 2 (0/2) | 0 | 18 (1.9%) |
| K. pinnata (Lam.) Pers. | 91 (45/23) | 60 (33/9) | 21 (8/8) | 78 (66/3) | 250 (25.9%) |
| K. thryrsiflora Harv. | 0 | 2 | 0 | 0 | 2 (0.2%) |
| K. tomentosa Baker | 1 (0/1) | 15 (0/15) | 8 (0/8) | 0 | 24 (2.5%) |
| K. tubiflora (Harv.) Raym.-Hamet | 0 | 20 (18/0) | 0 | 15 (6/6) | 35 (3.6%) |
| K. ×houghtonii D.B. Ward | 1 (0/1) | 91 (73/11) | 8 (2/5) | 5 (3/2) | 105 (10.9%) |
| K. ×laetivirens Desc. | 0 | 0 | 2 | 0 | 2 (0.2%) |
| Total records (%) | 105 (11%) | 679 (70%) | 56 (6%) | 127 (13%) | 967 (100%) |
| Protected Areas | Taxa | Records and Occurrence Status |
|---|---|---|
| Galapagos National Park | K. gastonis-bonnieri | Naturalized (1) |
| K. pinnata | Casual (2); Naturalized (5); Invasive (21); Uncertain (in the wild) (4) | |
| K. tubiflora | Invasive (1) | |
| K. ×houghtonii | Naturalized (2) | |
| Pululahua Geobotanical Reserve | K. pinnata | Uncertain (in the wild) (1); Uncertain (1) |
| K. densiflora | Naturalized (4) | |
| K. laxiflora | Casual (1); Naturalized (1); Invasive (14); Uncertain (in the wild) (2) | |
| Bellavista Private Protected Area | K. laxiflora | Casual (1) |
| Yasuní National Park | K. pinnata | Uncertain in the wild (2); Uncertain (1); Cultivated (1) |
| Cuyabeno–Lagartococha–Yasuní complex | K. pinnata | Naturalized (2); Uncertain (in the wild) (2); Cultivated (1) |
| Cayambe Coca National Park | K. ×houghtonii | Naturalized (1) |
| K. blossfeldiana | Cultivated (1) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vargas, A.; Herrera, I.; Nualart, N.; Guézou, A.; Gómez-Bellver, C.; Freire, E.; Jaramillo Díaz, P.; López-Pujol, J. The Genus Kalanchoe (Crassulaceae) in Ecuador: From Gardens to the Wild. Plants 2022, 11, 1746. https://doi.org/10.3390/plants11131746
Vargas A, Herrera I, Nualart N, Guézou A, Gómez-Bellver C, Freire E, Jaramillo Díaz P, López-Pujol J. The Genus Kalanchoe (Crassulaceae) in Ecuador: From Gardens to the Wild. Plants. 2022; 11(13):1746. https://doi.org/10.3390/plants11131746
Chicago/Turabian StyleVargas, Anahí, Ileana Herrera, Neus Nualart, Anne Guézou, Carlos Gómez-Bellver, Efraín Freire, Patricia Jaramillo Díaz, and Jordi López-Pujol. 2022. "The Genus Kalanchoe (Crassulaceae) in Ecuador: From Gardens to the Wild" Plants 11, no. 13: 1746. https://doi.org/10.3390/plants11131746