
Effects of Magnesium, Pyrophosphate and Phosphonates on Pyrophosphorolytic Reaction of UDP-Glucose Pyrophosphorylase



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Is Magnesium Required as Substrate for UGPase?
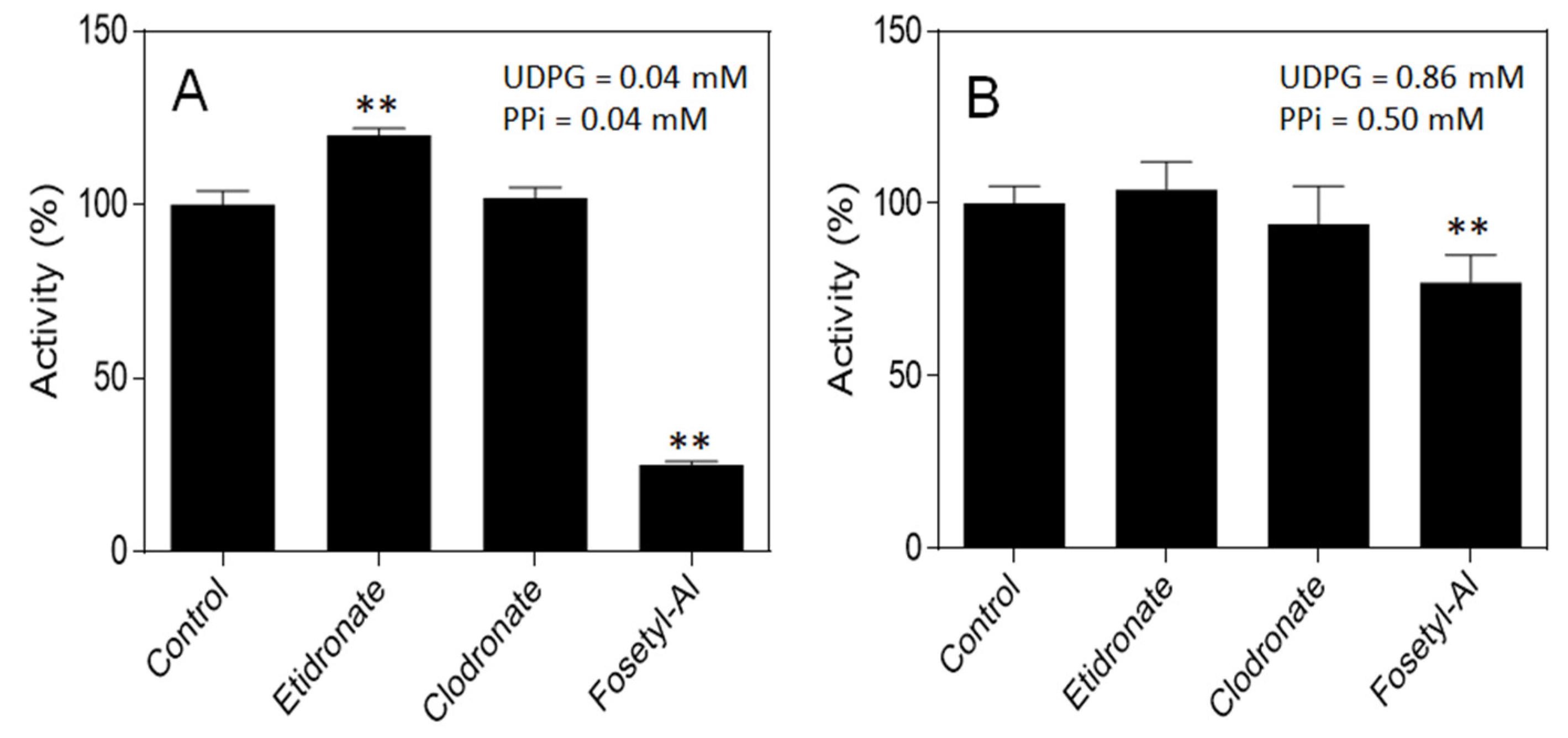
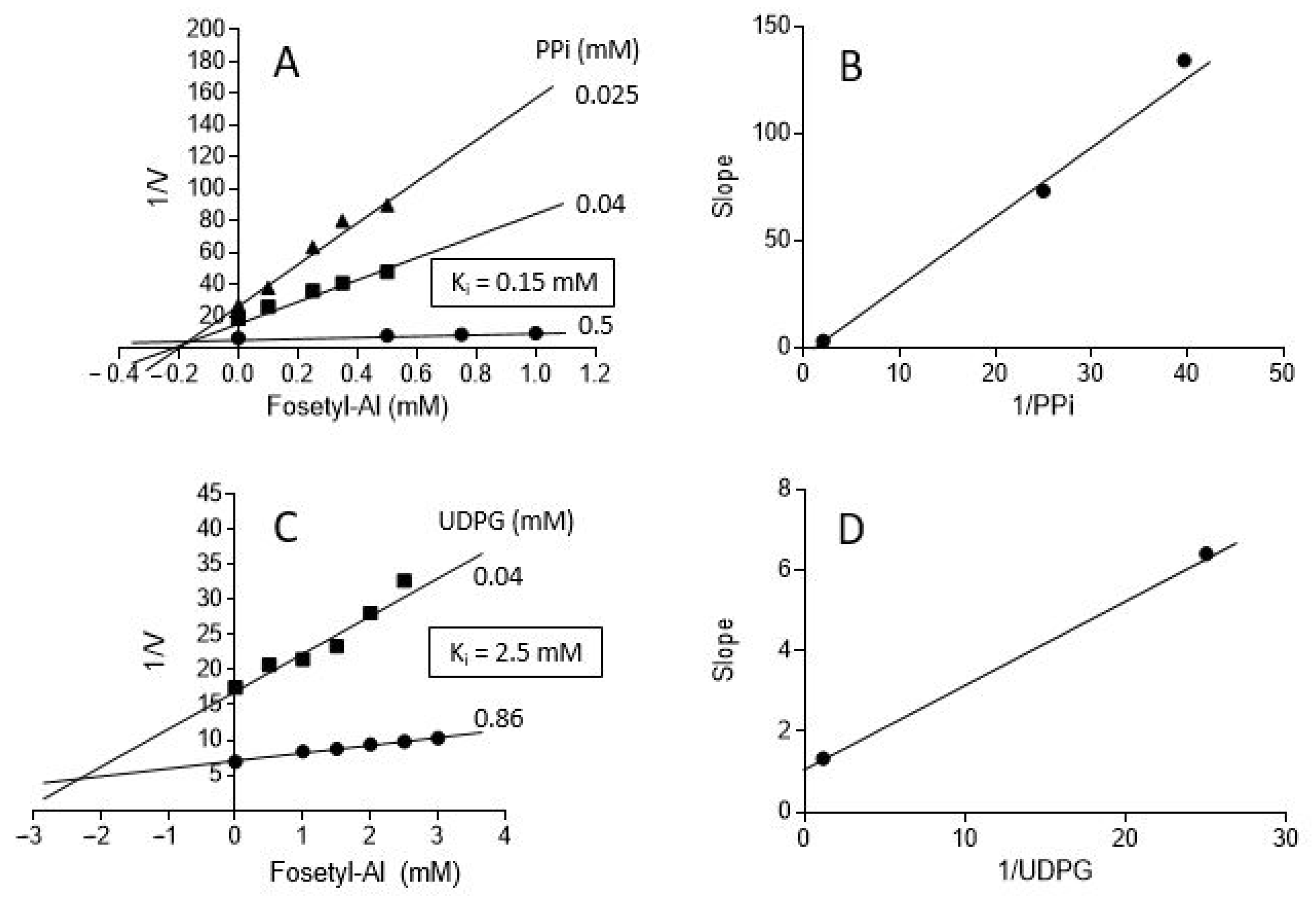
2.2. Effects of Selected PPi and Pi Analogs on UGPase Activity
3. Material and Methods
3.1. Materials
3.2. UGPase Assays
3.3. Magnesium and PPi Requirements
3.4. Effects of Phosphonates on UGPase Activity
3.5. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Meng, M.; Wilczynska, M.; Kleczkowski, L.A. Molecular and kinetic characterization of two UDP-glucose pyrophosphorylases, products of distinct genes, from Arabidopsis. Biochim. Biophys. Acta 2008, 1784, 967–972. [Google Scholar] [CrossRef] [PubMed]
- Decker, D.; Kleczkowski, L.A. UDP-sugar producing pyrophosphorylases—Distinct and essential enzymes with overlapping substrate specificities, providing de novo precursors for glycosylation reactions. Front. Plant Sci. 2019, 9, 1822. [Google Scholar] [CrossRef] [PubMed]
- Igamberdiev, A.U.; Kleczkowski, L.A. Magnesium and cell energetics in plants under anoxia. Biochem. J. 2011, 437, 373–379. [Google Scholar] [CrossRef] [PubMed]
- Igamberdiev, A.U.; Kleczkowski, L.A. Pyrophosphate as an alternative energy currency in plants. Biochem. J. 2021, 478, 1515–1524. [Google Scholar] [CrossRef] [PubMed]
- Kleczkowski, L.A. Back to the drawing board: Redefining starch synthesis in cereals. Trends Plant Sci. 1996, 1, 363–364. [Google Scholar] [CrossRef]
- Long, W.; Dong, B.; Wang, Y.; Pan, P.; Wang, Y.; Liu, L.; Chen, X.; Liu, X.; Liu, S.; Tian, Y.; et al. FLOURY ENDOSPERM8, encoding the UDP-glucose pyrophosphorylase 1, affects the synthesis and structure of starch in rice endosperm. J. Plant Biol. 2017, 60, 513–522. [Google Scholar] [CrossRef]
- Kleczkowski, L.A.; Igamberdiev, A.U. Optimization of nucleotide sugar supply for polysaccharide formation via thermodynamic buffering. Biochem. J. 2020, 477, 341–356. [Google Scholar] [CrossRef]
- Nakano, K.; Omura, M.; Tagaya, M.; Fukui, T. UDP-glucose pyrophosphorylase from potato tuber: Purification and characterization. J. Biochem. 1989, 106, 528–532. [Google Scholar] [CrossRef]
- Sowokinos, J.R.; Spychalla, J.P.; Desborough, J. Pyrophosphorylases in Solanum tuberosum. IV. Purification, tissue localization, and physicochemical properties of UDP-glucose pyrophosphorylase. Plant Physiol. 1993, 101, 1073–1080. [Google Scholar] [CrossRef] [Green Version]
- McCoy, J.G.; Bitto, E.; Bingman, C.A.; Wesenberg, G.E.; Bannen, R.M.; Kondrashov, D.A.; Phillips, G.N. Structure and dynamics of UDPglucose pyrophosphorylase from Arabidopsis thaliana with bound UDPglucose and UTP. J. Mol. Biol. 2007, 366, 830–841. [Google Scholar] [CrossRef] [Green Version]
- Tsuboi, K.K.; Fukunaga, K.; Petricciani, J.C. Purification and specific kinetic properties of erythrocyte uridine diphosphate glucose pyrophosphorylase. J. Biol. Chem. 1969, 244, 1008–1015. [Google Scholar] [CrossRef]
- Rudick, V.L.; Weisman, R.A. Uridine diphosphate glucose pyrophosphorylase of Acanthamoeba castellanii. Purification, kinetic, and developmental studies. J. Biol. Chem. 1974, 249, 7832–7840. [Google Scholar] [CrossRef]
- Yamagami, Y.; Huang, R.; Bevilacqua, P.C. Cellular concentrations of nucleotide diphosphate-chelated magnesium ions accelerate catalysis by RNA and DNA enzymes. Biochemistry 2019, 58, 3971–3979. [Google Scholar] [CrossRef] [PubMed]
- Kleczkowski, L.A.; Igamberdiev, A.U. Magnesium signaling in plants. Int. J. Mol. Sci. 2021, 22, 1159. [Google Scholar] [CrossRef]
- Zea, C.J.; Camci-Unal, G.; Pohl, N.L. Thermodynamics of binding of divalent magnesium and manganese to uridine phosphates: Implications for diabetes-related hypomagnesaemia and carbohydrate biocatalysis. Chem. Cent. J. 2008, 2, 15. [Google Scholar] [CrossRef] [Green Version]
- Igamberdiev, A.U.; Kleczkowski, L.A. Equilibration of adenylates in the mitochondrial intermembrane space maintains respiration and regulates cytosolic metabolism. J. Exp. Bot. 2006, 57, 2133–2141. [Google Scholar] [CrossRef] [Green Version]
- Park, J.; Pandya, V.R.; Ezekiel, S.J.; Berghuis, A.M. Phosphonate and bisphosphonate inhibitors of farnesyl pyrophosphate synthases: A structure-guided perspective. Front. Chem. 2021, 8, 612728. [Google Scholar] [CrossRef]
- Guest, D.I.; Grant, B. The complex action of phosphonates as antifungal agents. Biol. Rev. 1991, 66, 159–187. [Google Scholar] [CrossRef]
- Rogers, M.J. From molds and macrophages to mevalonate: A decade of progress in understanding the molecular mode of action of bisphosphonates. Calcif. Tissue Int. 2004, 75, 451–461. [Google Scholar] [CrossRef]
- Meng, M.; Fitzek, E.; Gajowniczek, A.; Wilczynska, M.; Kleczkowski, L.A. Domain-specific determinants of catalysis/substrate binding and the oligomerization status of barley UDP-glucose pyrophosphorylase. Biochim. Biophys. Acta 2009, 1794, 1734–1742. [Google Scholar] [CrossRef]
- Martin, H.; Grant, B.R.; Stehmann, C. Inhibition of inorganic pyrophosphatase by phosphonate—A site of action in Phytophthora spp. Pestic. Biochem. Physiol. 1998, 61, 65–77. [Google Scholar] [CrossRef]
- Grzechowiak, M.; Ruszkowski, M.; Sliwiak, J.; Szpotkowski, K.; Sikorski, M.; Jaskolski, M. Crystal structures of plant inorganic pyrophosphatase, an enzyme with a moonlighting autoproteolytic activity. Biochem. J. 2019, 476, 2297–2319. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, W.J.; Smithers, G.W. Stability constants for biologically important metal-ligand complexes. Methods Enzymol. 1979, 63, 294–336. [Google Scholar] [PubMed]
- Segel, I.H. Simple inhibition systems: Competitive inhibition. Enzyme Kinetics: Behavior and Analysis of Rapid Equilibrium and Steady-State Enzyme Systems, 1st ed.; John Wiley & Sons Inc.: New York, NY, USA, 1975; Volume 1, pp. 109–112. [Google Scholar]
- Kim, H.; Choi, J.; Kim, T.; Lokanath, N.K.; Ha, S.C.; Suh, S.W.; Hwang, H.W.; Kim, K.K. Structural basis for the reaction mechanism of UDP-glucose pyrophosphorylase. Mol. Cells 2010, 29, 397–405. [Google Scholar] [CrossRef]
- Geisler, M.; Wilczynska, M.; Karpinski, S.; Kleczkowski, L.A. Toward a blueprint for UDP-glucose pyrophosphorylase structure/function properties: Homology-modeling analyses. Plant Mol. Biol. 2004, 56, 783–794. [Google Scholar] [CrossRef]
- Figuerola, C.M.; Asencion Diez, M.D.; Ballicora, M.A.; Iglesias, A.A. Structure, function, and evolution of plant ADP-glucose pyrophosphorylase. Plant Mol. Biol. 2022, 108, 307–323. [Google Scholar] [CrossRef]
- Gustafson, G.L.; Ganders, J.E. Uridine diphosphate glucose pyrophosphorylase from Sorghum vulgare. Purification and kinetic properties. J. Biol. Chem. 1972, 247, 1387–1397. [Google Scholar] [CrossRef]
- Kotake, T.; Yamaguchi, D.; Ohzono, H.; Hojo, S.; Kaneko, S.; Ishida, H.K.; Tsumuraya, Y. UDP-sugar pyrophosphorylase with broad substrate specificity toward various monosaccharide 1-phosphates from pea sprouts. J. Biol. Chem. 2004, 279, 45728–45736. [Google Scholar] [CrossRef] [Green Version]
- Kleczkowski, L.A. Is leaf ADP-glucose pyrophosphorylase an allosteric enzyme? Biochim. Biophys. Acta 2000, 1476, 103–108. [Google Scholar] [CrossRef]
- Schnurr, J.A.; Storey, K.K.; Jung, H.G.; Somers, D.A.; Gronwald, J.W. UDP-sugar pyrophosphorylase is essential for pollen development in Arabidopsis. Planta 2006, 224, 520–532. [Google Scholar] [CrossRef]
- Igamberdiev, A.U.; Kleczkowski, L.A. Thermodynamic buffering, stable non-equilibrium and establishment of the computable structure of plant metabolism. Prog. Biophys. Mol. Biol. 2019, 146, 23–36. [Google Scholar] [CrossRef]
- Gutiérrez-Luna, F.M.; Hernández-Domínguez, E.E.; Valencia-Turcotte, L.G.; Rodríguez-Sotres, R. Pyrophosphate and pyrophosphatases in plants, their involvement in stress responses and their possible relationship to secondary metabolism. Plant Sci. 2018, 267, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Hørder, M. Enzyme catalyzed hydrolysis of inorganic pyrophosphate. Enzyme 1975, 19, 165–191. [Google Scholar] [CrossRef] [PubMed]
- Ferjani, A.; Kawade, K.; Asaoka, M.; Oikawa, A.; Okada, T.; Mochizuki, A.; Maeshima, M.; Hirai, M.Y.; Saito, K.; Tsukaya, H. Pyrophosphate inhibits gluconeogenesis by restricting UDP-glucose formation in vivo. Sci. Rep. 2018, 8, 14696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nilima, K.; Vinay, S. V-PPase in plants: An overview. Res. J. Biotechnol. 2008, 3, 57–63. [Google Scholar]
- Niere, J.; DeAngelis, G.; Grant, B.R. The effect of phosphonate on the acid-soluble phosphorus components in the genus Phytophthora. Microbiology 1994, 140, 1661–1670. [Google Scholar] [CrossRef] [Green Version]
- Dixon, M. The determination of enzyme inhibitor constants. Biochem. J. 1953, 55, 170–171. [Google Scholar] [CrossRef]
- Kazuta, Y.; Tagaya, M.; Tanizawa, K.; Fukui, T. Probing the pyrophosphate-binding site in potato tuber UDP-glucose pyrophosphorylase with pyridoxal diphosphate. Protein Sci. 1993, 2, 119–125. [Google Scholar] [CrossRef] [Green Version]
- Çal, Ö. The effects of fosetyl-Al application on morphology and viability of Lycopersicon esculentum Mill. pollen. Plant Soil Environ. 2008, 54, 336–340. [Google Scholar]
- Silva, P.V.; Vèlez, M.L.; Hernàndez Otano, D.; Nunez, C.; Greslebin, A.G. Action of fosetyl-al and metalaxyl against Phytophthora austrocedri. For. Path. 2016, 46, 54–66. [Google Scholar] [CrossRef]
- Hillebrand, S.; Zundel, J.L.; Tietjen, K. Fungicides with unknown mode of action. In Modern Crop Protection Compounds, 2nd ed.; Kramer, W., Ed.; Wiley-VCH Verlag: Weinheim, Germany, 2012; pp. 865–885. [Google Scholar] [CrossRef]
- Lin, B.; Qing, X.; Liao, J.; Zhuo, K. Role of protein glycosylation in host-pathogen interaction. Cells 2020, 9, 1022. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Ma, Z.; Qiu, M.; Guo, B.; Zhang, Q.; Jiang, H.; Zhang, B.; Lin, Y.; Xuan, M.; Sun, L.; et al. N-glycosylation shields Phytophthora sojae apoplastic effector PsXEG1 from a specific host aspartic protease. Proc. Natl. Acad. Sci. USA 2020, 117, 27685–27693. [Google Scholar] [CrossRef] [PubMed]
- Chahed, A.; Lazazzara, V.; Moretto, M.; Nesler, A.; Corneo, P.E.; Barka, E.A.; Pertot, I.; Puopolo, G.; Perazzolli, M. The differential growth inhibition of Phytophthora spp. caused by the rare sugar tagatose is associated with species-specific metabolic and transcriptional changes. Front. Microbiol. 2021, 12, 711545. [Google Scholar] [CrossRef] [PubMed]
- Kleczkowski, L.A.; Geisler, M.; Fitzek, E.; Wilczynska, M. A common structural blueprint for plant UDP-sugar producing pyrophosphorylases. Biochem. J. 2011, 439, 375–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Decker, D.; Öberg, C.; Kleczkowski, L.A. Identification and characterization of inhibitors of UDP-glucose and UDP-sugar pyrophosphorylases for in vivo studies. Plant J. 2017, 90, 1093–1107. [Google Scholar] [CrossRef] [Green Version]
- Decker, D.; Kleczkowski, L.A. Substrate specificity and inhibitor sensitivity of plant UDP-sugar producing pyrophosphorylases. Front. Plant Sci. 2017, 8, 1610. [Google Scholar] [CrossRef] [Green Version]
- Martz, F.; Wilczynska, M.; Kleczkowski, L.A. Oligomerization status, with the monomer as active species, defines catalytic efficiency of UDP-glucose pyrophosphorylase. Biochem. J. 2002, 367, 295–300. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kleczkowski, L.A.; Decker, D. Effects of Magnesium, Pyrophosphate and Phosphonates on Pyrophosphorolytic Reaction of UDP-Glucose Pyrophosphorylase. Plants 2022, 11, 1611. https://doi.org/10.3390/plants11121611
Kleczkowski LA, Decker D. Effects of Magnesium, Pyrophosphate and Phosphonates on Pyrophosphorolytic Reaction of UDP-Glucose Pyrophosphorylase. Plants. 2022; 11(12):1611. https://doi.org/10.3390/plants11121611
Chicago/Turabian StyleKleczkowski, Leszek A., and Daniel Decker. 2022. "Effects of Magnesium, Pyrophosphate and Phosphonates on Pyrophosphorolytic Reaction of UDP-Glucose Pyrophosphorylase" Plants 11, no. 12: 1611. https://doi.org/10.3390/plants11121611