
Impact of Horse Grazing on Floristic Diversity in Mediterranean Small Standing-Water Ecosystems (SWEs)

 ,
,  ,
,
Abstract
:1. Introduction
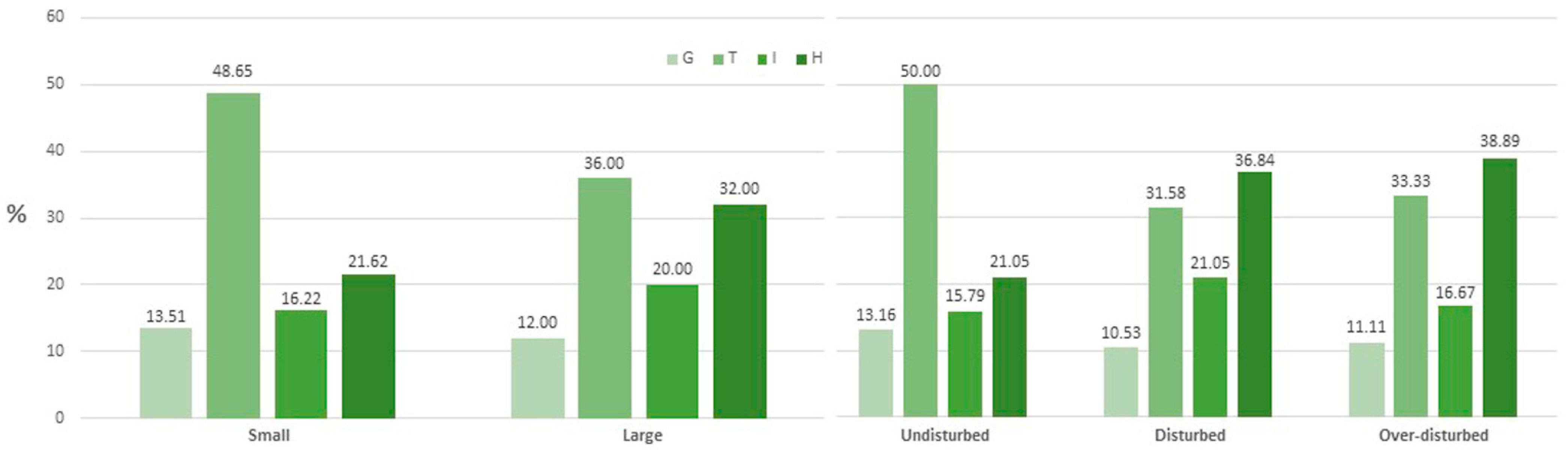
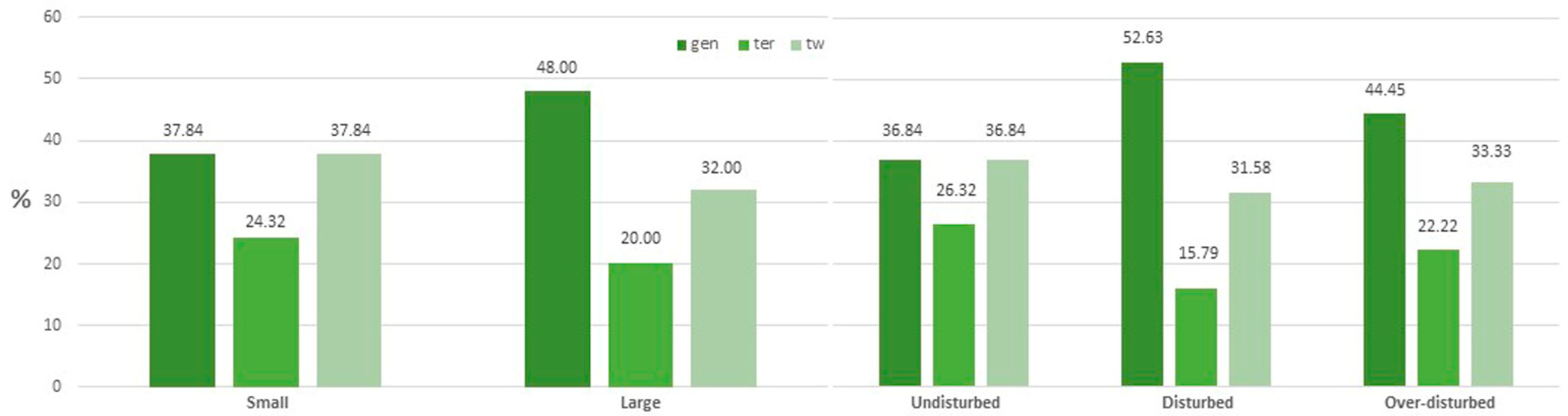
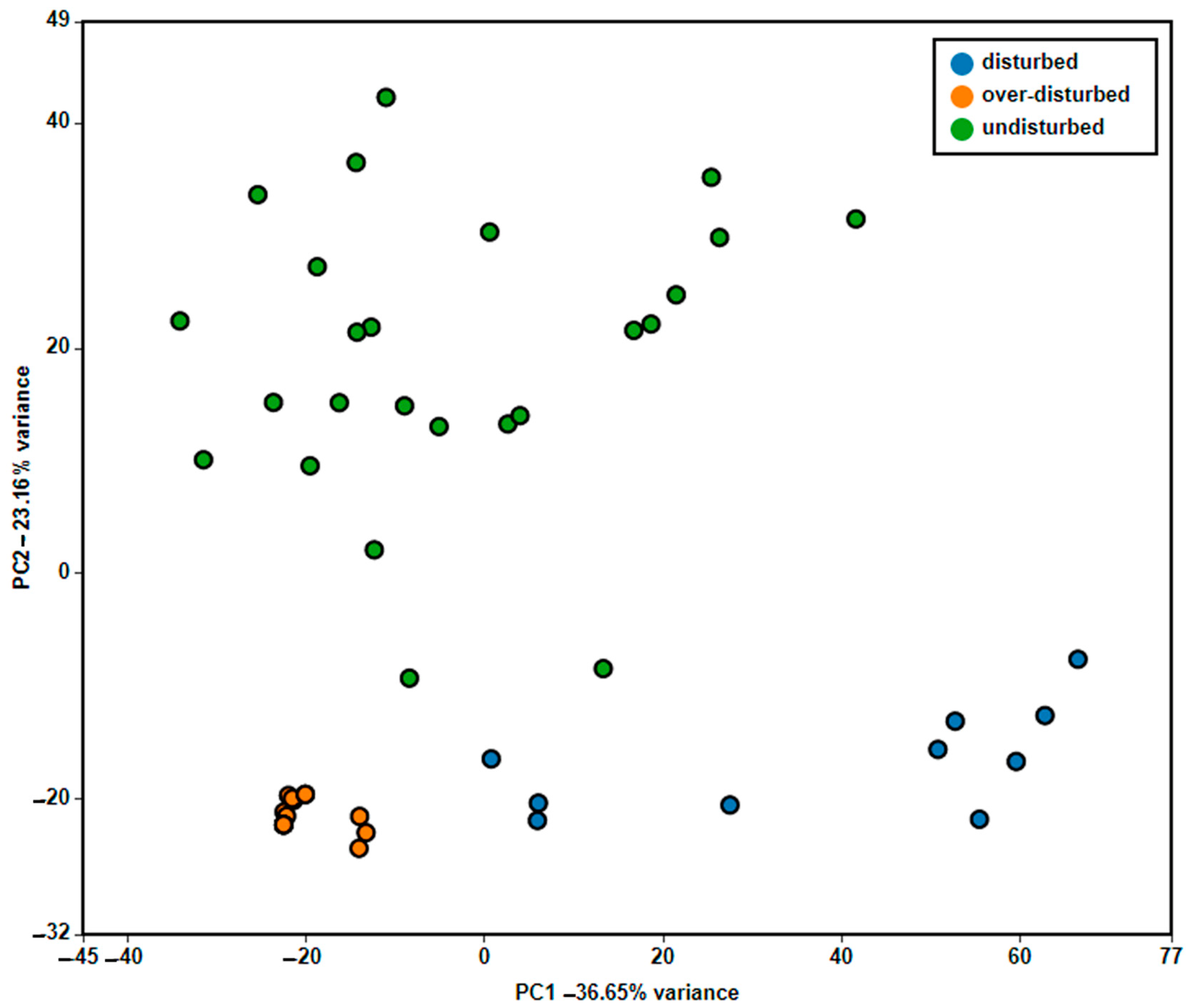
2. Results
Diversity Indices
3. Discussion
Implications for Conservation
4. Materials and Methods
4.1. Study Area
4.2. Experimental Design and Data Collection
4.3. Data Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Céréghino, R.; Biggs, J.; Oertli, B.; Declerck, S. The ecology of European ponds: Defining the characteristics of a neglected freshwater habitat. Hydrobiologia 2008, 597, 1–6. [Google Scholar] [CrossRef]
- Céréghino, R.; Boix, D.; Cauchie, H.M.; Martens, K.; Oertli, B. The ecological role of ponds in a changing world. Hydrobiologia 2014, 723, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Mullin, M.L.; Doyle, R.D. Big things come in small packages: Why limnologists should care about small ponds. Acta Limnol. Bras. 2019, 31, e105. [Google Scholar] [CrossRef] [Green Version]
- Bolpagni, R.; Poikane, S.; Laini, A.; Bagella, S.; Bartoli, M.; Cantonati, M. Ecological and Conservation Value of Small Standing-Water Ecosystems: A Systematic Review of Current Knowledge and Future Challenges. Water 2019, 11, 402. [Google Scholar] [CrossRef] [Green Version]
- Williams, P.; Whitfield, M.; Biggs, J.; Bray, S.; Fox, G.; Nicolet, P.; Sear, D. Comparative biodiversity of rivers, streams, ditches and ponds in an agricultural landscape in Southern England. Biol. Conserv. 2004, 115, 329–341. [Google Scholar] [CrossRef]
- Bagella, S.; Caria, M.C.; Zuccarello, V. Patterns of emblematic habitat types in Mediterranean temporary wetlands. Comptes Rendus Biol. 2010, 333, 694–700. [Google Scholar] [CrossRef]
- Calhoun, A.J.K.; Mushet, D.M.; Bell, K.P.; Boix, D.; Fitzsimons, J.A.; Isselin-Nondedeu, F. Temporary wetlands: Challenges and solutions to conserving a ‘disappearing’ ecosystem. Biol. Conserv. 2017, 211, 3–11. [Google Scholar] [CrossRef]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef]
- Rhazi, L.; Grillas, P.; Toure, A.M.; Ham, L.T. Impact of land use in catchment and human activities on water, sediment and vegetation of Mediterranean temporary pools. Comptes Rendus Acad. Sci. III-Vie 2001, 324, 165–177. [Google Scholar] [CrossRef]
- Junk, W.J.; An, S.; Finlayson, C.M.; Gopal, B.; Květ, J.; Mitchell, S.A.; Mitsch, W.J.; Robarts, R.D. Current state of knowledge regarding the world’s wetlands and their future under global climate change: A synthesis. Aquat. Sci. 2012, 75, 151–167. [Google Scholar] [CrossRef] [Green Version]
- Kneitel, J.M.; Lessin, C.L. Ecosystem-phase interactions: Aquatic eutrophication decreases terrestrial plant diversity in California vernal pools. Oecologia 2010, 163, 461–469. [Google Scholar] [CrossRef] [PubMed]
- Rolls, R.J.; Heino, J.; Ryder, D.S.; Chessman, B.C.; Growns, I.O.; Thompson, R.M.; Gido, K.B. Scaling 498 biodiversity responses to hydrological regimes. Biol. Rev. 2018, 93, 971–995. [Google Scholar] [CrossRef] [PubMed]
- Lefebvre, G.; Redmond, L.; Germain, C.; Palazzi, E.; Terzago, S.; Willm, L.; Poulin, B. Predicting the vulnerability of seasonally-flooded wetlands to climate change across the Mediterranean Basin. Sci. Total Environ. 2019, 692, 546–555. [Google Scholar] [CrossRef] [PubMed]
- Cramer, W.; Guiot, J.; Fader, M.; Garrabou, J.; Gattuso, J.-P.; Iglesias, A.; Lange, M.A.; Lionello, P.; Llasat, M.C.; Paz, S.; et al. Climate change and interconnected risks to sustainable development in the Mediterranean. Nat. Clim. Change 2018, 8, 972–980. [Google Scholar] [CrossRef] [Green Version]
- Čížková, H.; Květ, J.; Comín, F.A.; Laiho, R.; Pokorný, J.; Pithart, D. Actual state of European wetlands and their possible future in the context of global climate change. Aquati. Sci. 2013, 75, 3–26. [Google Scholar] [CrossRef] [Green Version]
- Pinna, M.S.; Loi, M.C.; Calderisi, G.; Fenu, G. Extremes Rainfall Events on Riparian Flora and Vegetation in the Mediterranean Basin: A Challenging but Completely Unexplored Theme. Water 2022, 14, 817. [Google Scholar] [CrossRef]
- Rhazi, L.; Grillas, P.; Rhazi, M.; Aznar, J.-C. Ten-year dynamics of vegetation in a Mediterranean temporary pool in western Morocco. Hydrobiologia 2009, 634, 185–194. [Google Scholar] [CrossRef]
- Caria, M.C.; Grillas, P.; Rivieccio, G.; Bagella, S. Effect of inter-annual meteorological fluctuations and wild boar disturbance on the vegetation of Mediterranean temporary ponds. Appl. Veg. Sci. 2021, 24, e12624. [Google Scholar] [CrossRef]
- Hulsmans, A.; Vanschoenwinkel, B.; Pyke, C.; Riddoch, B.J.; Brendonck, L. Quantifying the hydroregime of a temporary pool habitat: A modelling approach for ephemeral rock pools in SE Botswana. Ecosystems 2007, 11, 89–100. [Google Scholar] [CrossRef]
- Špoljar, M.; Shumka, S.; Tasevska, O.; Tomljanović, T.; Ostojić, A.; Balkić, A.G.; Lajtner, J.; Pepa, B.; Dražina, T.; Ternjej, I. Small Standing-Water Ecosystems in the Transitional Temperate Climate of the Western Balkans. In Small Water Bodies of the Western Balkans, 1st ed.; Pešić, V., Milošević, D., Miliša, M., Eds.; Springer: Cham, Switzerland, 2022; pp. 21–51. [Google Scholar]
- Sun, J.; Ma, B.; Lu, X. Grazing enhances soil nutrient effects: Trade-offs between aboveground and belowground biomass in alpine grasslands of the Tibetan Plateau. Land Degrad. Dev. 2018, 29, 337–348. [Google Scholar] [CrossRef]
- Huaranca, J.C.; Novaro, A.J.; Valdivia, C.E. Effects of livestock grazing on biodiversity: A meta-analysis on three trophic levels. J. Nat. Conserv. 2022, 66, 126126. [Google Scholar] [CrossRef]
- Fenu, G.; Cogoni, D.; Navarro, F.; Concas, E.; Bacchetta, G. The importance of the Cisto-Lavanduletalia coastal habitat on population persistence of the narrow endemic Dianthus morisianus (Caryophyllaceae). Plant Spec. Biol. 2017, 32, 156–168. [Google Scholar] [CrossRef]
- Lorite, J.; Salazar-Mendías, C.; Pawlak, R.; Cañadas, E.M. Assessing effectiveness of exclusion fences in protecting threatened plants. Sci. Rep. 2021, 11, 16124. [Google Scholar] [CrossRef]
- Pakeman, R.J.; Fielding, D.A.; Everts, L.; Littlewood, N.A. Long-term impacts of changed grazing regimes on the vegetation of heterogeneous upland grasslands. J. Appl. Ecol. 2019, 56, 1794–1805. [Google Scholar] [CrossRef]
- Wang, J.; Li, W.; Cao, W.; Wang, S. Effects of different intensities of long-term grazing on plant diversity, biomass and carbon stock in alpine shrubland on the Qinghai-Tibetan Plateau. PeerJ 2022, 10, e12771. [Google Scholar] [CrossRef] [PubMed]
- Barry, S.; Huntsinger, L. Rangeland land-sharing, livestock grazing’s role in the conservation of imperiled species. Sustainability 2021, 13, 4466. [Google Scholar] [CrossRef]
- Augustine, D.J.; McNaughton, S.J. Ungulate effects on the functional species composition of plant communities: Herbivore selectivity and plant tolerance. J. Wildl. Manag. 1998, 62, 1165–1183. [Google Scholar] [CrossRef]
- Marty, J.T. Effects of Cattle Grazing on Diversity in Ephemeral Wetlands. Conserv. Biol. 2005, 19, 1626–1632. [Google Scholar] [CrossRef]
- Rysiak, A.; Chabuz, W.; Sawicka-Zugaj, W.; Zdulski, J.; Grzywaczewski, G.; Kulik, M. Comparative impacts of grazing and mowing on the floristics of grasslands in the buffer zone of Polesie National Park, eastern Poland. Glob. Ecol. Conserv. 2021, 27, e01612. [Google Scholar] [CrossRef]
- Fenu, G.; Bacchetta, G.; Christodoulou, C.S.; Cogoni, D.; Fournaraki, C.; Gian Pietro, G.G.; Gotsiou, P.; Kyratzis, A.; Piazza, C.; Vicens, M.; et al. A common approach to the conservation of threatened island vascular plants: First results in the Mediterranean Basin. Diversity 2020, 12, 157. [Google Scholar] [CrossRef] [Green Version]
- Al-Rowaily, S.L.; El-Bana, M.I.; Al-Bakre, D.A.; Assaeed, A.M.; Hegazy, A.K.; Ali, M.B. Effects of open grazing and livestock exclusion on floristic composition and diversity in natural ecosystem of Western Saudi Arabia. Saudi J. Biol. Sci. 2015, 22, 430–437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lange-Bertalot, H.; Cavacini, P.; Tagliaventi, N.; Alfinito, S. Diatoms of Sardinia. Rare and 76 New Species in Rock Pools and Other Ephemeral Waters; Iconographia Diatomologica 12; A.R.G. Gantner Verlag KG: Ruggell, Germany, 2003. [Google Scholar]
- Rhazi, L.; Rhazi, M.; Grillas, P.; Khyari, D.E. Richness and structure of plant communities in temporary pools from western Morocco: Influence of human activities. Hydrobiologia 2006, 570, 197–203. [Google Scholar] [CrossRef]
- Schmitz, A.; Isselstein, J. Effect of Grazing System on Grassland Plant Species Richness and Vegetation Characteristics: Comparing Horse and Cattle Grazing. Sustainability 2020, 12, 3300. [Google Scholar] [CrossRef] [Green Version]
- Köhler, M.; Hiller, G.; Tischew, S. Year-round horse grazing supports typical vascular plant species, orchids and rare bird communities in a dry calcareous grassland. Agric. Ecosyst. Environ. 2016, 234, 48–57. [Google Scholar] [CrossRef]
- Saastamoinen, M.; Herzon, I.; Särkijärvi, S.; Schreurs, C.; Myllymäki, M. Horse Welfare and Natural Values on Semi-Natural and Extensive Pastures in Finland: Synergies and Trade-Offs. Land 2017, 6, 69. [Google Scholar] [CrossRef] [Green Version]
- Marion, B.; Bonis, A.; Bouzillé, J.-B. How much does grazing-induced heterogeneity impact plant diversity in wet grasslands? Écoscience 2010, 17, 229–239. [Google Scholar] [CrossRef]
- Davies, K.W.; Collins, G.; Boyd, C.S. Effects of feral free-roaming horses on semi-arid rangeland ecosystems: An example from the sagebrush steppe. Ecosphere 2014, 5, 1–14. [Google Scholar] [CrossRef]
- Marty, J.T. Loss of biodiversity and hydrologic function in seasonal wetlands persists over 10 years of livestock grazing removal. Restor. Ecol. 2015, 23, 548–554. [Google Scholar] [CrossRef]
- Bagella, S.; Caria, M.C.; Farris, E.; Filigheddu, R. Phytosociological analysis in Sardinian Mediterranean temporary wet habitats. Fitosociologia 2009, 46, 11–26. [Google Scholar]
- Bagella, S.; Caria, M.C.; Farris, E.; Filigheddu, R. Spatial-time variability and conservation relevance of plant communities in Mediterranean temporary wet habitats: A case study in Sardinia (Italy). Plant Biosyst. 2009, 143, 435–442. [Google Scholar] [CrossRef]
- Bagella, S.; Caria, M.C. Diversity and ecological characteristics of vascular flora in Mediterranean temporary pools. Comptes Rendus Biol. 2012, 335, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Deil, U. Distribution and ecology of some rare plant species in Northern Morocco II: Stemmacantha longifolia var. ericeticola. Acta Bot. Malacitana 1997, 22, 255–259. [Google Scholar] [CrossRef] [Green Version]
- Barbour, M.; Solomeshch, A.; Witham, C.; Holland, R.; MacDonald, R.; Cilliers, S.; Molina, J.A.; Buck, J.; Hillman, J. Vernal pool vegetation of California: Variation within pools. Madroño 2003, 50, 129–146. [Google Scholar]
- Papanastasis, V.P.; Kyriakakis, S.; Kazakis, G. Plant diversity in relation to overgrazing and burning in mountain Mediterranean ecosystems. J. Mediterr. Ecol. 2002, 3, 53–64. [Google Scholar]
- Sawidis, T.; Kalyva, S.; Delivopoulos, S. The root-tuber anatomy of Asphodelus aestivus. Flora 2005, 200, 332–338. [Google Scholar] [CrossRef]
- Noy-Meir, I.; Oron, T. Effects of grazing on geophytes in Mediterranean vegetation. J. Veg. Sci. 2001, 12, 749–760. [Google Scholar] [CrossRef]
- Deil, U. A review on habitats, plant traits and vegetation of ephemeral wetlands-A global perspective. Phytocoenologia 2005, 35, 533–705. [Google Scholar] [CrossRef]
- Caria, M.C.; Capra, G.F.; Buondonno, A.; Seddaiu, G.; Vacca, S.; Bagella, S. Small-scale patterns of plant functional types and soil features within Mediterranean temporary ponds. Plant Biosyst. 2015, 149, 384–394. [Google Scholar] [CrossRef]
- Pinna, M.S.; Cogoni, D.; Fenu, G.; Bacchetta, G. The conservation status and anthropogenic impacts assessments of Mediterranean coastal dunes. Estuar. Coast. Shelf Sci. 2015, 167, 25–31. [Google Scholar] [CrossRef]
- Pinna, M.S.; Bacchetta, G.; Cogoni, D.; Fenu, G. Is vegetation an indicator for evaluating the impact of tourism on the conservation status of Mediterranean coastal dunes? Sci. Total Environ. 2019, 674, 255–263. [Google Scholar] [CrossRef]
- Calderisi, G.; Cogoni, D.; Pinna, M.S.; Fenu, G. Recognizing the relative effects of environmental versus human factors to understand the conservation of coastal dunes areas. Reg. Stud. Mar. Sci. 2021, 48, 102070. [Google Scholar] [CrossRef]
- Menard, C.; Duncan, P.; Fleurance, G.; Georges, J.-Y.; Lila, M. Comparative foraging and nutrition of horses and cattle in European wetlands. J. Appl. Ecol. 2002, 39, 120–133. [Google Scholar] [CrossRef] [Green Version]
- Santoro, R.; Jucker, T.; Prisco, I.; Carboni, M.; Battisti, C.; Acosta, A.T.R. Effects of Trampling Limitation on Coastal Dune Plant Communities. Environ. Manag. 2012, 49, 534–542. [Google Scholar] [CrossRef] [PubMed]
- Biggs, J.; Corfield, A.; Walker, D.; Whitfield, M.; Williams, P. New approaches to the management of ponds. Br. Wildl. 1994, 5, 273–287. [Google Scholar]
- Moeslund, J.E.; Andersen, D.K.; Brunbjerg, A.K.; Fløjgaard, C.; Nygaard, B.; Ejrnæs, R. High nutrient loads hinder successful restoration of natural habitats in freshwater wetlands. bioRxiv 2022, 483603. [Google Scholar] [CrossRef]
- Fenu, G.; Mattana, E.; Congiu, A.; Bacchetta, G. The Endemic Vascular Flora of Supramontes (Sardinia), a Priority Plant Conservation Area. Candollea 2010, 65, 347–358. [Google Scholar] [CrossRef]
- Cañadas, E.M.; Fenu, G.; Peñas, J.; Lorite, J.; Mattana, E.; Bacchetta, G. Hotspots within hotspots: Endemic plant richness, environmental drivers, and implications for conservation. Biol. Conserv. 2014, 170, 282–291. [Google Scholar] [CrossRef]
- Marchi, M.; Murgia, M.V.; Maccioni, L.; Mossa, L.; Scrugli, A.; Mulas, B.; Fogu, M.C.; Cogoni, A.; Schenk, H. Sa Jara. Un’area di interesse naturalistico da salvaguardare; Pubblisar editrice Cagliari: Cagliari, Italy, 1989. [Google Scholar]
- Nicolet, P.; Biggs, J.; Fox, G.; Hodson, M.J.; Reynolds, C.; Whitfield, M.; Williams, P. The wetland plant and macroinvertebrate assemblages of temporary ponds in England and Wales. Biol. Conserv. 2004, 120, 261–278. [Google Scholar] [CrossRef]
- Della Bella, V.; Bazzanti, M.; Dowgiallo, M.G.; Iberite, M. Macrophyte diversity and physico-chemical characteristics of Tyrrhenian coast ponds in central Italy: Implications for conservation. Hydrobiologia 2008, 597, 85–95. [Google Scholar] [CrossRef]
- Grillas, P.; Gauthier, P.; Yavercovski, N.; Perennou, C. Species Information Sheets. In Mediterranean Temporary Pools; Station Biologique de la Tour du Valat: Arles, France, 2004; Volume 2. [Google Scholar]
- Pignatti, S. Flora d’Italia, 1st ed.; Edagricole: Bologna, Italy, 1982; Volumes 1–3. [Google Scholar]
- Pignatti, S.; Guarino, R.; La Rosa, M. Flora d’Italia, 2nd ed.; Edagricole: Bologna, Italy, 2017; Volumes 1–4. [Google Scholar]
- Bartolucci, F.; Peruzzi, L.; Galasso, G.; Albano, A.; Alessandrini, A.; Ardenghi, N.M.G.; Astuti, G.; Bacchetta, G.; Ballelli, S.; Banfi, E.; et al. An updated checklist of the vascular flora native to Italy. Plant Biosyst. 2018, 152, 179–303. [Google Scholar] [CrossRef]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral. Ecol. 2001, 26, 32–46. [Google Scholar]
- Grunewald, R.; Schubert, H. The definition of a new plant diversity index “H′dune” for assessing human damage on coastal dunes-Derived from the Shannon index of entropy H′. Ecol. Indic. 2007, 7, 1–21. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Size | Treatment | ||||
|---|---|---|---|---|---|
| Small | Large | Undisturbed | Disturbed | Over-Disturbed | |
| Eudianthe laeta Rchb. ex Willk. (tw) | 1.76 | 0.00 | 1.76 | 0.00 | 0.00 |
| Eleocharis palustris (L.) Roem. & Schult. subsp. palustris (gen) | 1.43 | 0.92 | 1.41 | 1.25 | 0.25 |
| Helosciadium crassipes W.D.J.Koch ex Rchb. (tw) | 1.09 | 2.46 | 1.17 | 2.89 | 1.00 |
| Illecebrum verticillatum L. (ter) | 0.13 | 0.71 | 0.13 | 1.00 | 0.69 |
| Alisma plantago-aquatica L. (gen) | 0.00 | 0.86 | 0.00 | 0.00 | 0.86 |
| Av. Abund | Av. Sim | Sim/SD | Contrib.% | Cum% | |
|---|---|---|---|---|---|
| Small | |||||
| Eudianthe laeta Rchb. ex Willk. | 32.81 | 20.56 | 1.73 | 47.02 | 47.02 |
| Eleocharis palustris (L.) Roem. & Schult. subsp. palustris (gen) | 20.80 | 9.18 | 0.99 | 20.99 | 68.01 |
| Helosciadium crassipes W.D.J.Koch ex Rchb. | 16.03 | 5.57 | 0.71 | 12.74 | 80.75 |
| Large | |||||
| Alisma plantago-aquatica L. | 6.01 | 17.61 | 0.83 | 37.97 | 37.97 |
| Helosciadium crassipes W.D.J.Koch ex Rchb. | 24.80 | 13.78 | 0.62 | 29.72 | 67.69 |
| Illecebrum verticillatum L. | 4.02 | 9.16 | 0.56 | 19.76 | 84.45 |
| Undisturbed | |||||
| Eudianthe laeta Rchb. ex Willk. | 32.81 | 20.56 | 1.73 | 47.02 | 47.02 |
| Eleocharis palustris (L.) Roem. & Schult. subsp. palustris (gen) | 20.80 | 9.18 | 0.99 | 20.99 | 68.01 |
| Helosciadium crassipes W.D.J.Koch ex Rchb. | 16.03 | 5.57 | 0.71 | 12.74 | 80.75 |
| Disturbed | |||||
| Helosciadium crassipes W.D.J.Koch ex Rchb. | 59.00 | 44.39 | 2.74 | 77.78 | 77.78 |
| Over-disturbed | |||||
| Alisma plantago-aquatica L. | 10.01 | 25.15 | 1.18 | 60.18 | 60.18 |
| Illecebrum verticillatum L. | 6.03 | 13.09 | 0.72 | 31.32 | 91.50 |
| Size | Treatment | ||||
|---|---|---|---|---|---|
| Small | Large | Undisturbed | Disturbed | Over-Disturbed | |
| No. species | 14.44 ± 0.33 | 6.08 ± 0.28 | 14.44 ± 0.33 | 6.40 ± 0.37 | 5.87 ± 0.39 |
| Hmodified | 1.65 ± 0.07 | 0.72 ± 0.07 | 1.65 ± 0.07 | 0.96 ± 0.09 | 0.56 ± 0.08 |
| Hmodified-max | 3.02 ± 0.10 | 1.15 ± 0.12 | 3.02 ± 0.10 | 1.77 ± 0.11 | 0.74 ± 0.09 |
| Emodified | 0.55 ± 0.01 | 0.68 ± 0.03 | 0.55 ± 0.01 | 0.55 ± 0.05 | 0.76 ± 0.03 |
| N | 1 | 1 | 1 | 1 | 1 |
| EI | 0 | 0 | 0 | 0 | 0 |
| Variable | Max Neg. Difference | Max. Pos. Difference | p-Level | Mean Small | Mean Large | Std. Dev. Small | Std. Dev. Large | Valid N. Small | Valid N. Large |
|---|---|---|---|---|---|---|---|---|---|
| No. species | 0.0000 | 1.0000 | p < 0.001 | 14.4400 | 6.0800 | 1.6350 | 1.3820 | 25 | 25 |
| Hmodified | 0.0000 | 0.8400 | p < 0.001 | 1.6475 | 0.7223 | 0.3475 | 0.3523 | 25 | 25 |
| Emodified | −0.6400 | 0.0400 | p < 0.001 | 0.5464 | 0.6801 | 0.0738 | 0.1591 | 25 | 25 |
| Undisturbed | Disturbed | Over-Disturbed | ||
|---|---|---|---|---|
| No. species Kruskal–Wallis test: H(2, N = 50) = 37.6312; p < 0.0001 | Undisturbed | 4.363485 | 5.419084 | |
| Disturbed | 4.363485 | 0.336067 | ||
| Over-Disturbed | 5.419084 | 0.336067 | ||
| Hmodified Kruskal–Wallis test: H(2, N = 50) = 35.3240; p < 0.0001 | Undisturbed | 3.135109 | 5.797160 | |
| Disturbed | 3.135109 | 1.764353 | ||
| Over-Disturbed | 5.797160 | 1.764353 | ||
| Emodified Kruskal–Wallis test: H(2, N = 50) = 25.5510; p < 0.0001 | Undisturbed | 0.234675 | 4.847770 | |
| Disturbed | 0.234675 | 3.663133 | ||
| Over-Disturbed | 4.847770 | 3.663133 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fenu, G.; Melis, A.; Pinna, M.S.; Loi, M.C.; Calderisi, G.; Cogoni, D. Impact of Horse Grazing on Floristic Diversity in Mediterranean Small Standing-Water Ecosystems (SWEs). Plants 2022, 11, 1597. https://doi.org/10.3390/plants11121597
Fenu G, Melis A, Pinna MS, Loi MC, Calderisi G, Cogoni D. Impact of Horse Grazing on Floristic Diversity in Mediterranean Small Standing-Water Ecosystems (SWEs). Plants. 2022; 11(12):1597. https://doi.org/10.3390/plants11121597
Chicago/Turabian StyleFenu, Giuseppe, Arianna Melis, Maria Silvia Pinna, Maria Cecilia Loi, Giulia Calderisi, and Donatella Cogoni. 2022. "Impact of Horse Grazing on Floristic Diversity in Mediterranean Small Standing-Water Ecosystems (SWEs)" Plants 11, no. 12: 1597. https://doi.org/10.3390/plants11121597