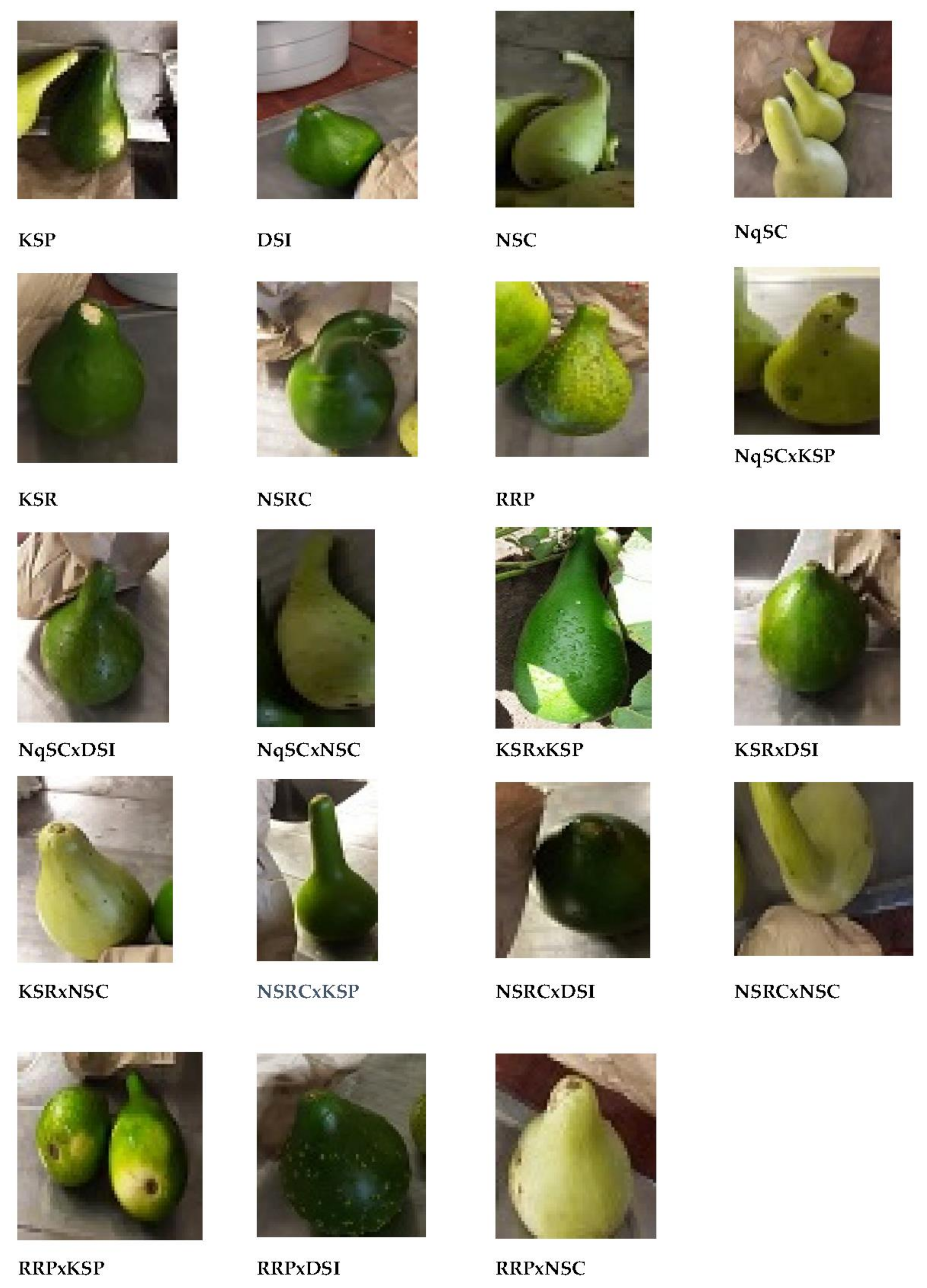
3.2. Qualitative Traits Evaluation
The 19 genotypes (seven parental landraces and 12 F
1 progenies) evaluated in the present study showed a wide range of diversity in qualitative traits including growth habit, stem shape, leaf color, leaf margin, fruit color, fruit texture, fruit shape, seed color, seed shape, and seed size (
Table 1 and
Table 2). Related findings were reported on
L. siceraria landraces characterization, where the greatest variation was in fruit shape, fruit texture, fruit color, seed size, seed shape, and seed color [
10]. The present results revealed that different landraces display different morphology. However, plant vigor, tendrils, stem color, leaf shape, flower color, flower size, tendrils branching, tendril type, fruit pulp color, seed texture, and seed lines were constant for all evaluated genotypes. Corresponding findings were reported by Kalyanrao et al. [
10] who observed that
L. siceraria genotypes were similar in stem color, tendrils, tendril branching, leaf shape, and flower color but differ in fruit and most seed traits.
Observed diversity in fruit and seed qualitative traits among the genotypes in the present study could be due to the different sites of origin of the seven landraces crossed and the mixing of genes in the controlled cross-pollination performed for generating 12 first filial generations (F1 progenies). It was observed that the F1 offspring inherited either the male or female fruit morphological traits (fruit color, fruit texture, and fruit shape) in the current findings. For example, the cross between the DSI (maternal parent) with dark green smooth isodiametric fruits and the RRP (paternal parent) with green rough isodiametric fruits generated green rough isodiametric fruits of the RRPxDSI (F1 progeny).
3.3. Quantitative Traits Evaluation
Seven landraces and twelve F
1 progenies evaluated in the present study showed a wide range of diversity in quantitative traits including days to emergence, emergence percentage, seedling height, leaf area, main vine length, days to flowering, days to edible harvest maturity, days to drying harvest maturity, fruit neck length, seed width, and leaf growth percentage. There was no significant difference for days to 50% emergence in a study by Mashilo et al. [
11] in
Lagenaria siceraria landraces collected from Limpopo, whereas in the present study there was a significant difference in days to emergence among the 19 evaluated genotypes. The variation observed in the days to emergence was mainly due to differences in sites of origin for the landraces. In the current study, two landraces (RRP and NSRC) reached 50% emergence before all F
1 offspring emerged and one parental line (NqSC) emerged later than all F
1 progenies. Similar findings were reported on the significance of emergence percentage among
L. siceraria genotypes that ranged from 65.8% to 85.3% [
12].
The highest emergence percentage was obtained from F
1 progeny (KSRxKSP), while the lowest was observed from the parental line (NqSC). This means F
1 progenies displayed heterosis, where the progeny of a cross performs better than either parent. Accessions of
L. siceraria collected from Turkey displayed the same range in seedling height (22–56 mm), but their mean (39 mm) was lower than the one obtained in the present study [
3]. This observation confirms that different growing environments affect the performance of
Lagenaria siceraria populations. Buthelezi et al. [
9] obtained lower cotyledon leaf areas which ranged from 880–1726 mm
2 and a mean of 1289 mm
2 amongst the landraces of
L. siceraria. The diversity in seedling traits obtained in the current study may be due to the genetics of different
L. siceraria genotypes [
13].
A shorter main vine length was obtained in the present study than reported amongst
Lagenaria siceraria lines (4.8 to 6.8 m) by Uddin et al. [
13]. In the present study, we observed a larger leaf area (2581 to 7000 mm
2) than the results reported on
L. siceraria landraces by Mashilo et al. [
11]. The findings obtained in the current study agree with the observations on days to flowering in
L. siceraria that ranged from 44.0 to 65.8 days from a study by Iqbal et al. [
12]. The current findings on days to drying harvest maturity correspond with published findings that
L. siceraria reaches maturity from 60–120 days after planting [
14]. Mashilo et al. [
11] reported a longer maximum fruit neck length (24.1 cm) and shorter minimum fruit neck length (0.53 cm) than the present study amongst
L. siceraria landraces. Thus, different genotypes differ in the fruit neck length which affects their usage when the fruits are dry. For example, isodiametric-shaped fruits have a short neck, therefore the matured dried fruits are used for storing water and milk. The dried curvilinear shaped fruits with long necks are used as cups for traditional beer in rural communities.
Results on seed width from the present study agreed with a reported study on African
L. siceraria accessions, where the seed width ranged from 5.30 to 12.34 mm [
2]. A greater
L. siceraria leaf growth percentage with a mean of 5281% was recorded in the previous study by Buthelezi et al. [
9]. However, cotyledon leaf area, number of leaves, number of branches, leaf chlorophyll content, total fruit mass per plot, number of seeds per fruit, fruit mass, fruit rind thickness, hundred seed mass, total seed mass, fruit length, fruit width, seed length, shoot growth percentage, fruit yield, and yield per plant were not significantly different (
p > 0.05) for the evaluated genotypes in the current study. These results of the present study disagree with reported results, where significant differences (
p < 0.05) were obtained in most quantitative traits of
Lagenaria siceraria [
11].
The positive correlation between emergence percentage with fruit yield, number of seeds per fruit, and vine length suggests that the selection of genotypes with high emergence percentage results in genotypes with higher fruit yield, many seeds per fruit, and long vines. A positive correlation was observed in the current study between the number of seeds per fruit with the vine length, the number of branches, and total fruit mass, which is in accordance with the reported findings [
15]. Similar correlations were reported between the number of branches and the vine length h (r = 0.614) and the number of leaves (r = 0.678) [
9].
A comparable positive correlation was obtained by Mashilo et al. [
11] between hundred seed mass and the vine length (r = 0.45) and the number of branches (r = 0.42). Seed length was also reported to be positively correlated with fruit length and fruit rind thickness which also correlated positively with fruit circumference in the comparable study [
2]. The present results are like those of Buthelezi et al. [
9] who also reported that seed mass positively correlated with hundred seed mass (r = 0.966) and fruit width (r = 0.732); fruit yield was also reported to correlate positively with fruit width (r = 0.702) and seed width was also reported to be positively correlated to tendril traits. Total fruit mass positively correlated with all other variables studied on
L. siceraria accessions in a study by Mlandenovic et al. [
2].
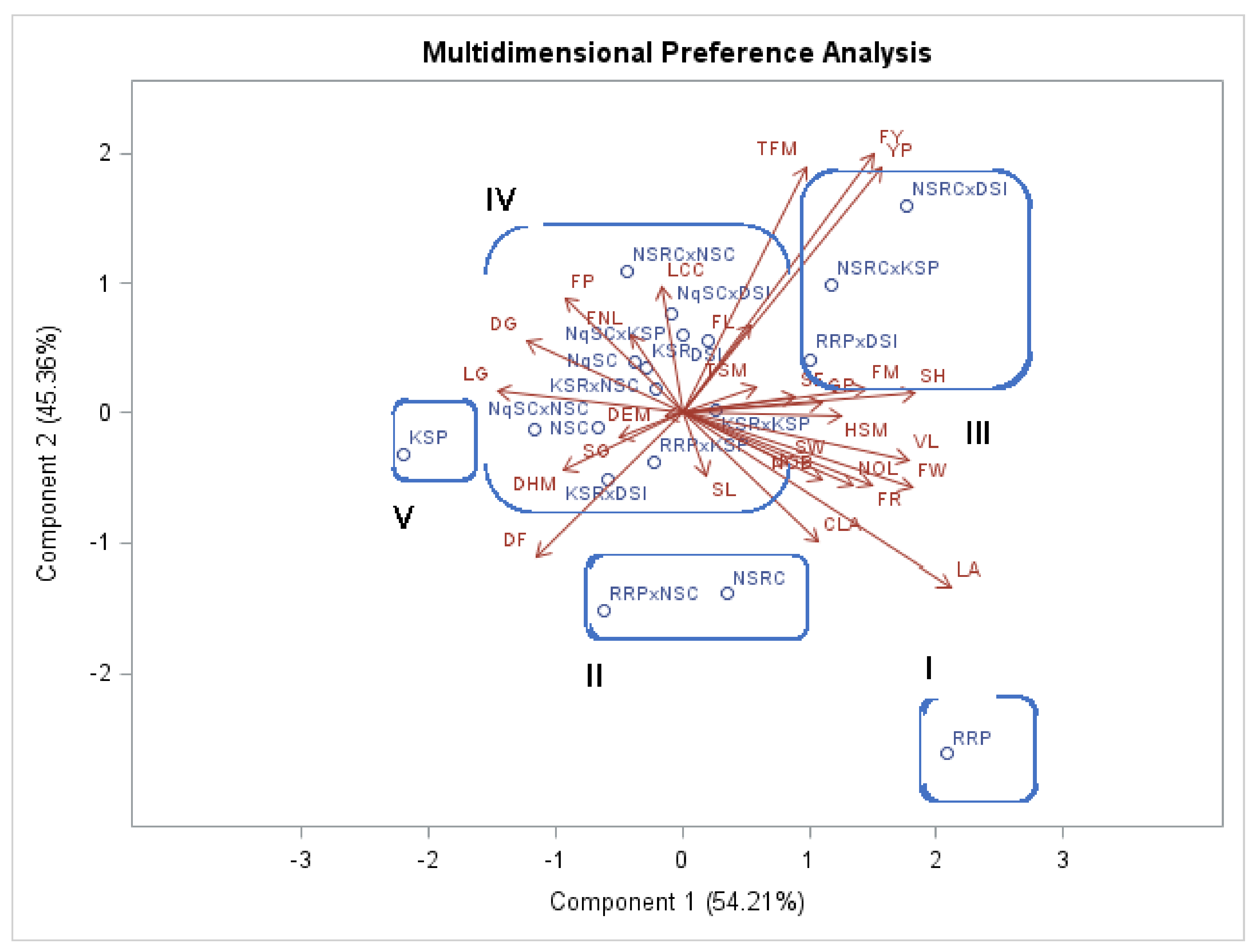
The first principal component (PC1) included several traits that contributed to a higher variation than PC2, as shown by the principal component analysis of the quantitative traits studied. Fruit width and fruit mass were negatively correlated with the first principal component. In the first and second principal components, the traits with high coefficients were considered more important for the explanation of total variability. Seedling height, cotyledon size, vine length, leaf area, number of leaves, number of branches, fruit mass, fruit rind thickness, fruit width, hundred seed mass, and number of seeds per fruit established the greatest variability among the genotypes. Mashilo et al. [
11] reported comparable findings, where the principal component analysis indicated that most of the variation in
L. siceraria is contributed by fruit and seed traits.
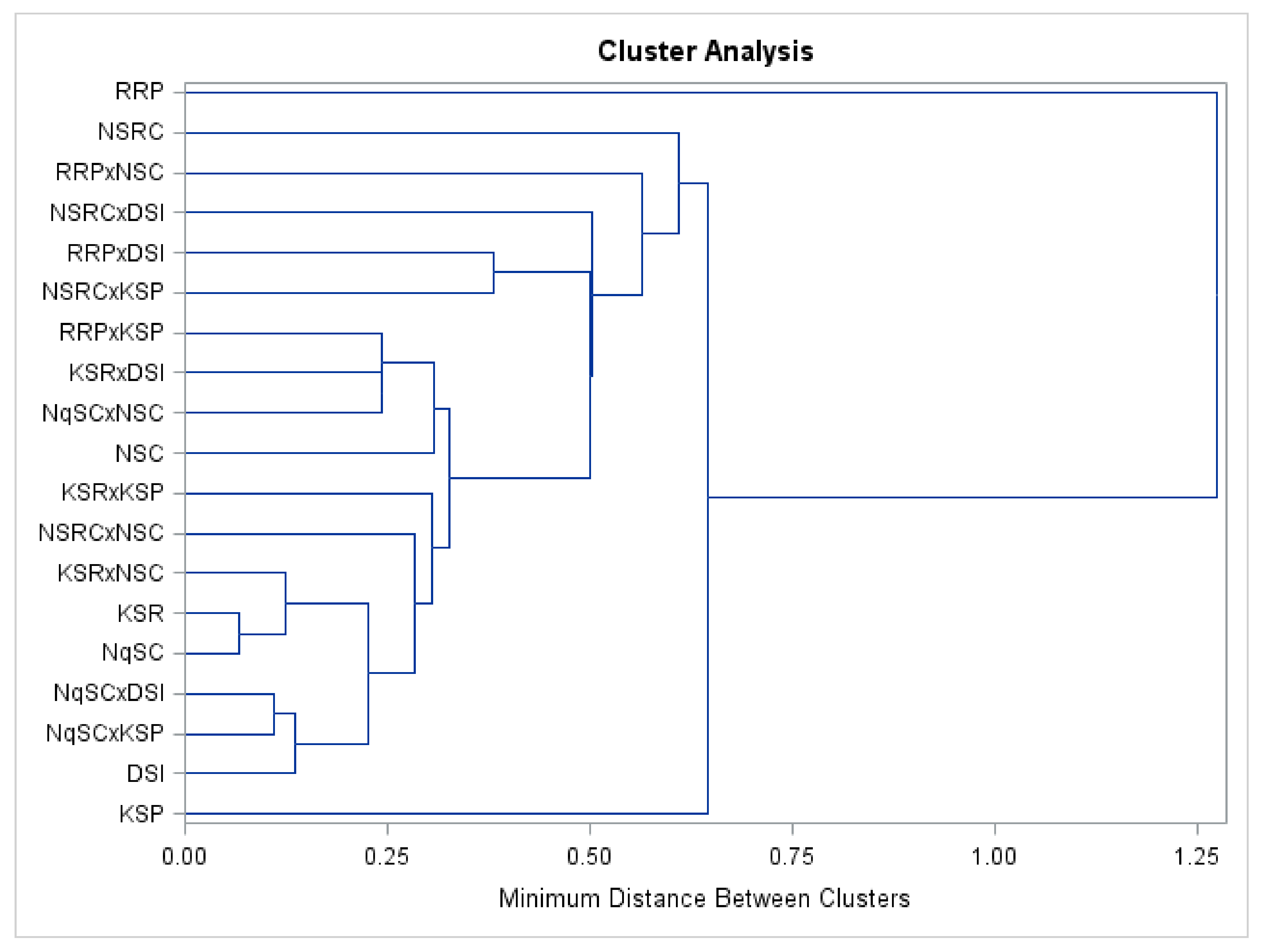
Clustering of genotypes according to fruit texture agreed with a previous study conducted on
Lagenaria siceraria landraces by Buthelezi et al. [
9]. It was reported that genotypes with analogous morphology belong to one cluster [
8] and that genotypes with identical fruit shape, fruit length, fruit mass, and fruit circumference grouped together [
2]. Cluster analysis conducted on quantitative traits grouped the evaluated genotypes into five clusters showing an adequate heritable variation that could permit rational selection.
The genotypic coefficient of variation was generally lower than the phenotypic coefficient of variability signifying a strong impact of the environment concerning the expressiveness of genes in the phenotypic display (
Table 7). Supportive results were obtained in a study conducted on the genotypes of
Pisum sativum, where the genotypic coefficient was generally lower than the phenotypic coefficient of variability [
16]. A higher coefficient of variation was obtained with leaf growth percentage in the current study, but pod bearing length exhibited a higher value in the study conducted on field pea by Meena et al. [
16]. Higher heritability estimates were recorded for fruit length, vine length, seed width, fruit weight, rind thickness, leaf area, the thickness of seed, number of seeds per plant, and fruit circumference in another study [
2]. In most of the traits evaluated in the current study, the genetic advance was lower than the one obtained in a study conducted on
Corchorus accessions by Dube et al. [
17].










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}