
Isolation and Characterization of the GmMT-II Gene and Its Role in Response to High Temperature and Humidity Stress in Glycine max

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
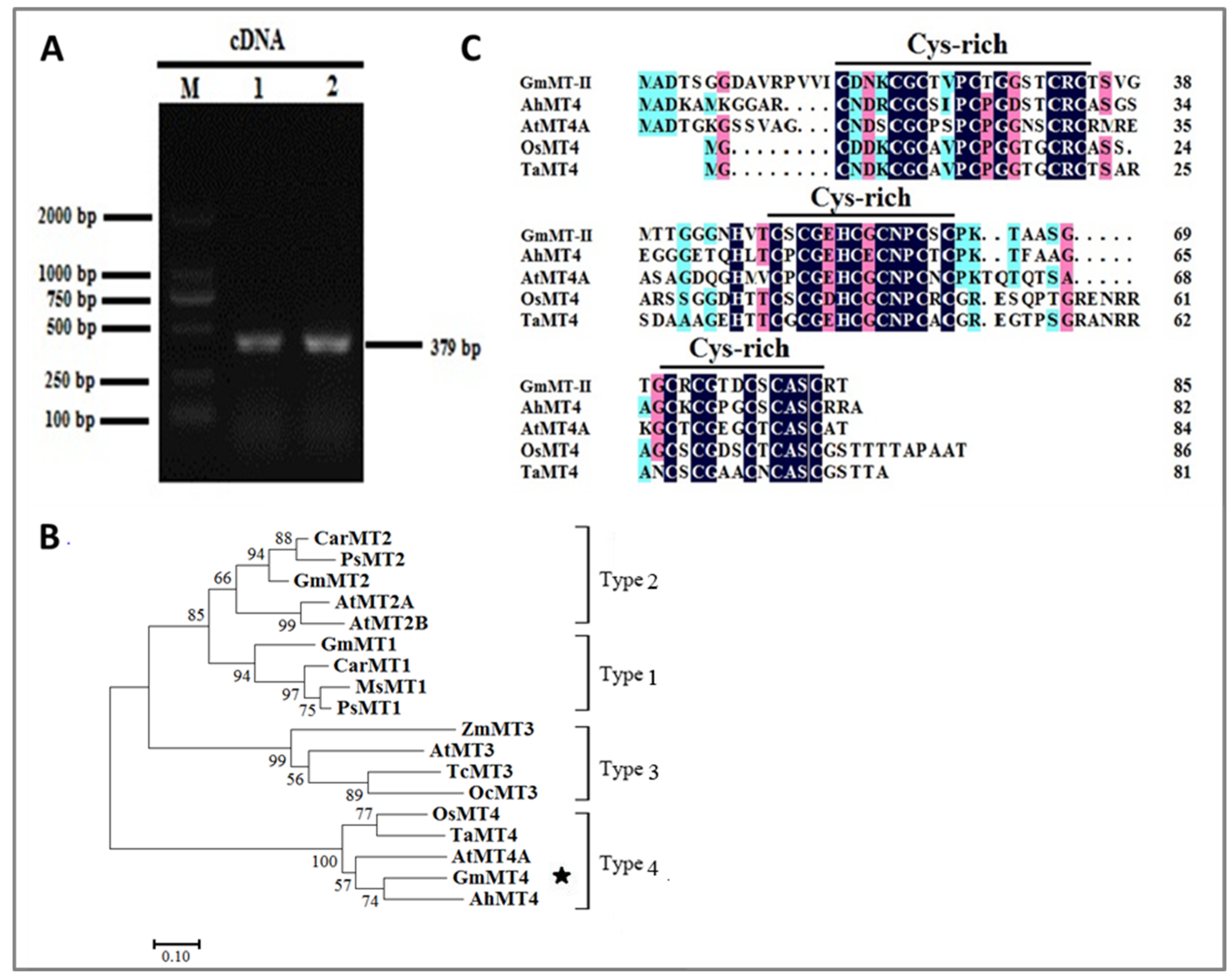
2.1. Sequence Analysis of GmMT-II in Glycine Max
2.2. Expression of the GmMT-II Gene in Glycine max
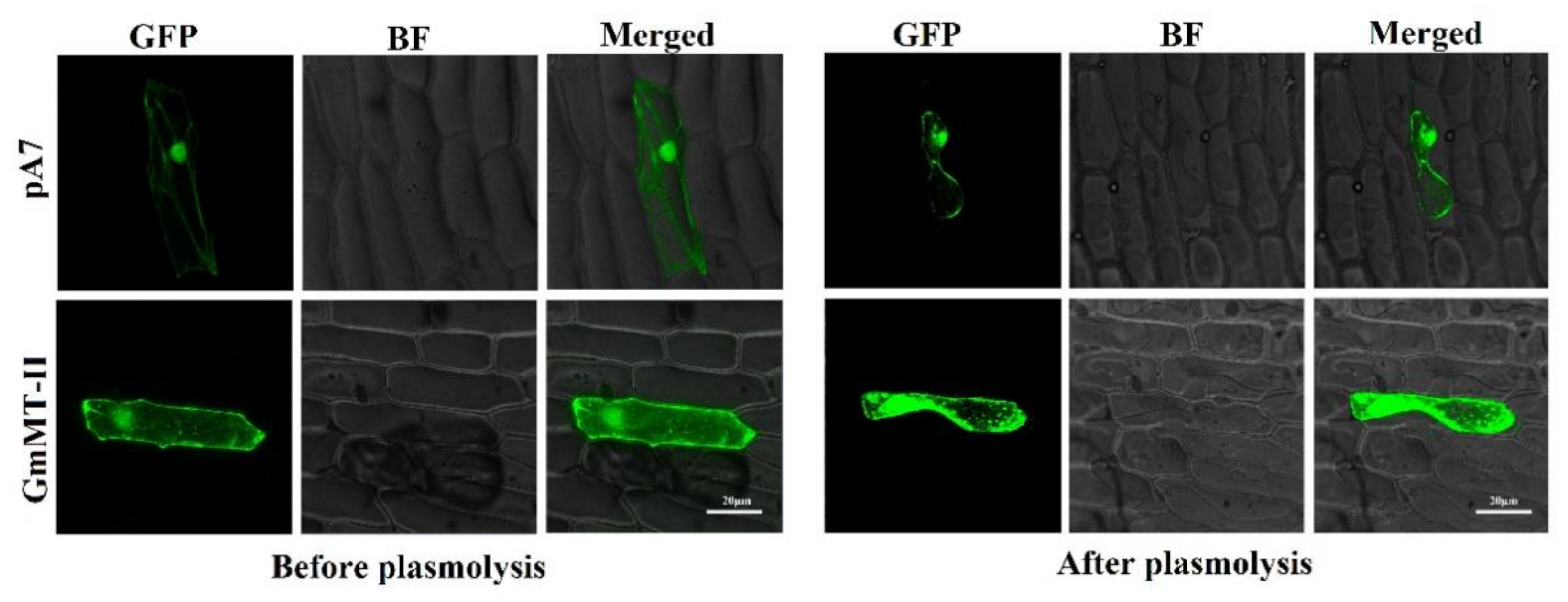
2.3. Subcellular Localization of GmMT-II Protein
2.4. Screening of Gm1-MMP and GmMT-II interaction sites in Yeast
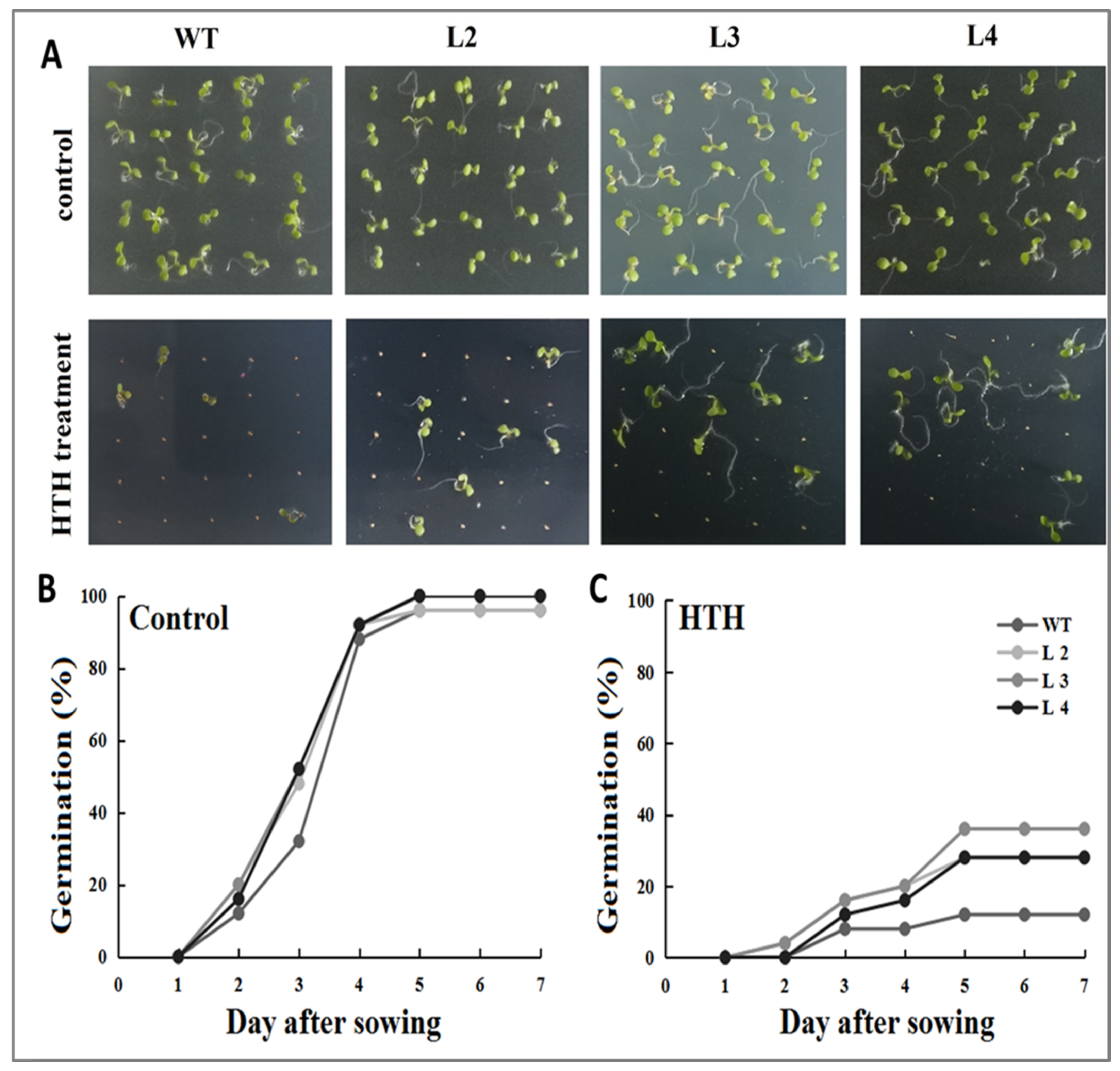
2.5. Response of the GmMT-II Overexpression Transgenic Arabidopsis to HTH Stress
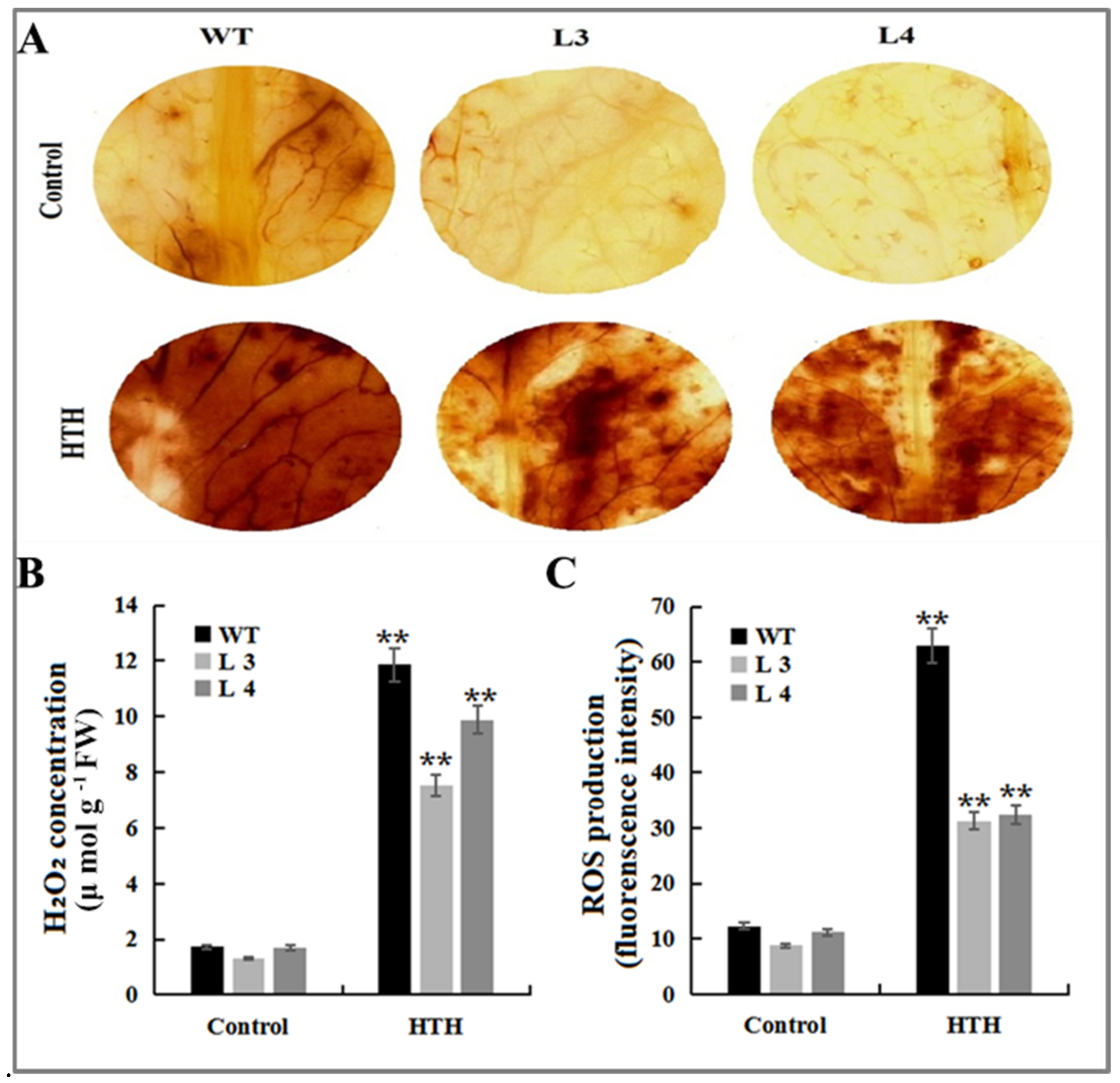
2.6. GmMT-II Overexpression Transgenic Arabidopsis Lines Reduces Oxidative Stress under HTH
2.7. GmMT-II Overexpression Transgenic Arabidopsis Lines Increase Antioxidant Activities under HTH Stress
3. Discussion
4. Materials and Methods
4.1. Sequence Analysis of Metallothioneins in Glycine Max
4.2. Plant Materials and Stress Treatment
4.3. RNA Extraction, cDNA Library and qRT-PCR
4.4. Subcellular Localization and Microscopic Analysis
4.5. Screening of Gm1-MMP and GmMT-II Interaction Sites in Yeast
4.6. Plasmid Construction and Transformation of Arabidopsis
4.7. Measurement of Lipid Peroxidation (MDA)
4.8. Histochemical Analysis of Hydrogen Peroxide (H2O2) and Reactive Oxygen Species (ROS) Analysis
4.9. Determination of H2O2 Content
4.10. Determination of ROS Production
4.11. Analysis of Antioxidants Enzyme Activity
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dong, L.; Fang, C.; Cheng, Q.; Su, T.; Kou, K.; Kong, L.; Zhang, C.; Li, H.; Hou, Z.; Zhang, Y.; et al. Genetic basis and adaptation trajectory of soybean from its temperate origin to tropics. Nat. Commun. 2021, 12, 5445. [Google Scholar] [CrossRef] [PubMed]
- Sobko, O.; Stahl, A.; Hahn, V.; Zikeli, S.; Claupein, W.; Gruber, S. Environmental Effects on Soybean (Glycine Max (L.) Merr) Production in Central and South Germany. Agronomy 2020, 10, 1847. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.B.; Zulfiqar, F.; Raza, A.; Mohsin, S.M.; Al Mahmud, J.; Fujita, M.; Fotopoulos, V. Reactive Oxygen Species and Antioxidant Defense in Plants under Abiotic Stress: Revisiting the Crucial Role of a Universal Defense Regulator. Antioxidants 2020, 9, 681. [Google Scholar] [CrossRef] [PubMed]
- Deshmukh, R.; Sonah, H.; Patil, G.; Chen, W.; Prince, S.; Mutava, R.; Vuong, T.; Valliyodan, B.; Nguyen, H.T. Integrating omic approaches for abiotic stress tolerance in soybean. Front. Plant Sci. 2014, 5, 244. [Google Scholar] [CrossRef]
- Alsajri, F.A.; Singh, B.; Wijewardana, C.; Irby, J.T.; Gao, W.; Reddy, K.R. Evaluating Soybean Cultivars for Low- and High-Temperature Tolerance During the Seedling Growth Stage. Agronomy 2019, 9, 13. [Google Scholar] [CrossRef] [Green Version]
- Jumrani, K.; Bhatia, V.S. Impact of combined stress of high temperature and water deficit on growth and seed yield of soybean. Physiol. Mol. Biol. Plants 2017, 24, 37–50. [Google Scholar] [CrossRef]
- Miransari, M. Soybean, Protein, and Oil Production Under Stress. In Environmental Stresses in Soybean Production; Miransari, M., Ed.; Academic Press: San Diego, CA, USA, 2016; pp. 157–176. ISBN 9780128015353. [Google Scholar]
- Nakagawa, A.C.S.; Ario, N.; Tomita, Y.; Tanaka, S.; Murayama, N.; Mizuta, C.; Iwaya-Inoue, M.; Ishibashi, Y. High temperature during soybean seed development differentially alters lipid and protein metabolism. Plant Prod. Sci. 2020, 23, 504–512. [Google Scholar] [CrossRef] [Green Version]
- Chennupati, P.; Seguin, P.; Chamoun, R.; Jabaji, S. Effects of High-Temperature Stress on Soybean Isoflavone Concentration and Expression of Key Genes Involved in Isoflavone Synthesis. J. Agric. Food Chem. 2012, 60, 12421–12427. [Google Scholar] [CrossRef]
- Liu, S.; Jia, Y.; Zhu, Y.; Zhou, Y.; Shen, Y.; Wei, J.; Liu, X.; Liu, Y.; Gu, W.; Ma, H. Soybean Matrix Metalloproteinase Gm2-MMP Relates to Growth and Development and Confers Enhanced Tolerance to High Temperature and Humidity Stress in Transgenic Arabidopsis. Plant Mol. Biol. Report. 2018, 36, 94–106. [Google Scholar] [CrossRef]
- Allen, L.H.; Zhang, L.; Boote, K.J.; Hauser, B.A. Elevated temperature intensity, timing, and duration of exposure affect soybean internode elongation, mainstem node number, and pod number per plant. Crop J. 2018, 6, 148–161. [Google Scholar] [CrossRef]
- Chowdhury, M.; Kiraga, S.; Islam, N.; Ali, M.; Reza, N.; Lee, W.-H.; Chung, S.-O. Effects of Temperature, Relative Humidity, and Carbon Dioxide Concentration on Growth and Glucosinolate Content of Kale Grown in a Plant Factory. Foods 2021, 10, 1524. [Google Scholar] [CrossRef] [PubMed]
- Shu, Y.; Tao, Y.; Wang, S.; Huang, L.; Yu, X.; Wang, Z.; Chen, M.; Gu, W.; Ma, H. GmSBH1, a homeobox transcription factor gene, relates to growth and development and involves in response to high temperature and humidity stress in soybean. Plant Cell Rep. 2015, 34, 1927–1937. [Google Scholar] [CrossRef] [PubMed]
- Shu, Y.; Zhou, Y.; Mu, K.; Hu, H.; Chen, M.; He, Q.; Huang, S.; Ma, H.; Yu, X. A transcriptomic analysis reveals soybean seed pre-harvest deterioration resistance pathways under high temperature and humidity stress. Genome 2020, 63, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Ma, H.; Song, L.; Shu, Y.; Gu, W. Comparative proteomics analysis reveals the mechanism of pre-harvest seed deterioration of soybean under high temperature and humidity stress. J. Proteom. 2012, 75, 2109–2127. [Google Scholar] [CrossRef] [PubMed]
- Capdevila, M.; Atrian, S. Metallothionein protein evolution: A miniassay. JBIC J. Biol. Inorg. Chem. 2011, 16, 977–989. [Google Scholar] [CrossRef]
- Jin, S.; Xu, C.; Li, G.; Sun, D.; Li, Y.; Wang, X.; Liu, S. Functional characterization of a type 2 metallothionein gene, SsMT2, from alkaline-tolerant Suaeda salsa. Sci. Rep. 2017, 7, 17914. [Google Scholar] [CrossRef] [Green Version]
- Patankar, H.V.; Al-Harrasi, I.; Al Kharusi, L.; Jana, G.A.; Al-Yahyai, R.; Sunkar, R.; Yaish, M.W. Overexpression of a Metallothionein 2A Gene from Date Palm Confers Abiotic Stress Tolerance to Yeast and Arabidopsis thaliana. Int. J. Mol. Sci. 2019, 20, 2871. [Google Scholar] [CrossRef] [Green Version]
- Hasan, K.; Cheng, Y.; Kanwar, M.K.; Chu, X.-Y.; Ahammed, G.J.; Qi, Z.-Y. Responses of Plant Proteins to Heavy Metal Stress—A Review. Front. Plant Sci. 2017, 8, 1492. [Google Scholar] [CrossRef] [Green Version]
- Mir, G.; Domènech, J.; Huguet, G.; Guo, W.-J.; Goldsbrough, P.; Atrian, S.; Molinas, M. A plant type 2 metallothionein (MT) from cork tissue responds to oxidative stress. J. Exp. Bot. 2004, 55, 2483–2493. [Google Scholar] [CrossRef] [Green Version]
- Kawka, B.; Kwiecień, I.; Ekiert, H. Influence of Culture Medium Composition and Light Conditions on the Accumulation of Bioactive Compounds in Shoot Cultures of Scutellaria lateriflora L. (American Skullcap) Grown In Vitro. Appl. Biochem. Biotechnol. 2017, 183, 1414–1425. [Google Scholar] [CrossRef] [Green Version]
- Freisinger, E. Structural features specific to plant metallothioneins. JBIC J. Biol. Inorg. Chem. 2011, 16, 1035–1045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cobbett, C.; Goldsbrough, P. Phytochelatins and metallothioneins: Roles in Heavy Metal Detoxification and Homeostasis. Annu. Rev. Plant Biol. 2002, 53, 159–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, J.; Chen, D.; Ren, Y.; Zhang, X.; Zhao, J. Characteristic and Expression Analysis of a Metallothionein Gene, OsMT2b, Down-Regulated by Cytokinin Suggests Functions in Root Development and Seed Embryo Germination of Rice. Plant Physiol. 2008, 146, 1637–1650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, W.; Bundithya, W.; Goldsbrough, P.B. Characterization of the Arabidopsis metallothionein gene family: Tissue-specific expression and induction during senescence and in response to copper. New Phytol. 2003, 159, 369–381. [Google Scholar] [CrossRef]
- Chen, Y.-Y.; Li, M.-Y.; Li, Y.; Wang, F.; Xu, Z.-S.; Xiong, A.-S. Isolation and characterization of the Agmt2 gene and its response to abiotic and metalstress in Apium graveolens. Sci. Hortic. 2015, 186, 1–6. [Google Scholar] [CrossRef]
- Mishra, M.K.; Tiwari, S.; Misra, P. Overexpression of WssgtL3.1 gene from Withania somnifera confers salt stress tolerance in Arabidopsis. Plant Cell Rep. 2021, 40, 2191–2204. [Google Scholar] [CrossRef]
- Chaturvedi, A.K.; Patel, M.K.; Mishra, A.; Tiwari, V.; Jha, B. The SbMT-2 Gene from a Halophyte Confers Abiotic Stress Tolerance and Modulates ROS Scavenging in Transgenic Tobacco. PLoS ONE 2014, 9, e111379. [Google Scholar] [CrossRef]
- Xue, T.; Li, X.; Zhu, W.; Wu, C.; Yang, G.; Zheng, C. Cotton metallothionein GhMT3a, a reactive oxygen species scavenger, increased tolerance against abiotic stress in transgenic tobacco and yeast. J. Exp. Bot. 2008, 60, 339–349. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Rahman, A.; Mahmud, J.A.; Hossain, M.S.; Fujita, M. Soybean Production and Environmental Stresses. In Environmental Stresses in Soybean Production; Miransari, M., Ed.; Academic Press: San Diego, CA, USA, 2016; pp. 61–102. ISBN 9780128015353. [Google Scholar]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Brun, T.; Rabuske, J.E.; Todero, I.; Almeida, T.C.; Junior, J.J.D.; Ariotti, G.; Confortin, T.; Arnemann, J.A.; Kuhn, R.C.; Guedes, J.V.C.; et al. Production of bioherbicide by Phoma sp. in a stirred-tank bioreactor. 3 Biotech 2016, 6, 230. [Google Scholar] [CrossRef] [Green Version]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Henriques, R.; Lin, S.-S.; Niu, Q.-W.; Chua, N.-H. Agrobacterium-mediated transformation of Arabidopsis thaliana using the floral dip method. Nat. Protoc. 2006, 1, 641–646. [Google Scholar] [CrossRef] [PubMed]
- Jahan, M.S.; Guo, S.; Sun, J.; Shu, S.; Wang, Y.; El-Yazied, A.A.; Alabdallah, N.M.; Hikal, M.; Mohamed, M.H.; Ibrahim, M.F.; et al. Melatonin-mediated photosynthetic performance of tomato seedlings under high-temperature stress. Plant Physiol. Biochem. 2021, 167, 309–320. [Google Scholar] [CrossRef] [PubMed]
- Oparka, M.; Walczak, J.; Malinska, D.; van Oppen, L.M.P.E.; Szczepanowska, J.; Koopman, W.J.H.; Wieckowski, M.R. Quantifying ROS levels using CM-H2DCFDA and HyPer. Methods 2016, 109, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Barja, G. The quantitative measurement of H2O2 generation in isolated mitochondria. J. Bioenerg. Biomembr. 2002, 34, 227–233. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-O.; Kang, H. Comparative expression analysis of genes encoding metallothioneins in response to heavy metals and abiotic stresses in rice (Oryza sativa) and Arabidopsis thaliana. Biosci. Biotechnol. Biochem. 2018, 82, 1656–1665. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, S.; Liu, Y.; Liu, C.; Li, Y.; Zhang, F.; Ma, H. Isolation and Characterization of the GmMT-II Gene and Its Role in Response to High Temperature and Humidity Stress in Glycine max. Plants 2022, 11, 1503. https://doi.org/10.3390/plants11111503
Liu S, Liu Y, Liu C, Li Y, Zhang F, Ma H. Isolation and Characterization of the GmMT-II Gene and Its Role in Response to High Temperature and Humidity Stress in Glycine max. Plants. 2022; 11(11):1503. https://doi.org/10.3390/plants11111503
Chicago/Turabian StyleLiu, Sushuang, Yanmin Liu, Chundong Liu, Yang Li, Feixue Zhang, and Hao Ma. 2022. "Isolation and Characterization of the GmMT-II Gene and Its Role in Response to High Temperature and Humidity Stress in Glycine max" Plants 11, no. 11: 1503. https://doi.org/10.3390/plants11111503