
Genome-Wide Identification of MDH Family Genes and Their Association with Salt Tolerance in Rice


Abstract
:1. Introduction
2. Results
2.1. Identification and Characterization of MDH Genes in Rice
2.2. Phylogenetic Analysis of MDH Proteins
2.3. Putative Cis-Regulatory Elements Analysis in the Promoter of OsMDH Genes
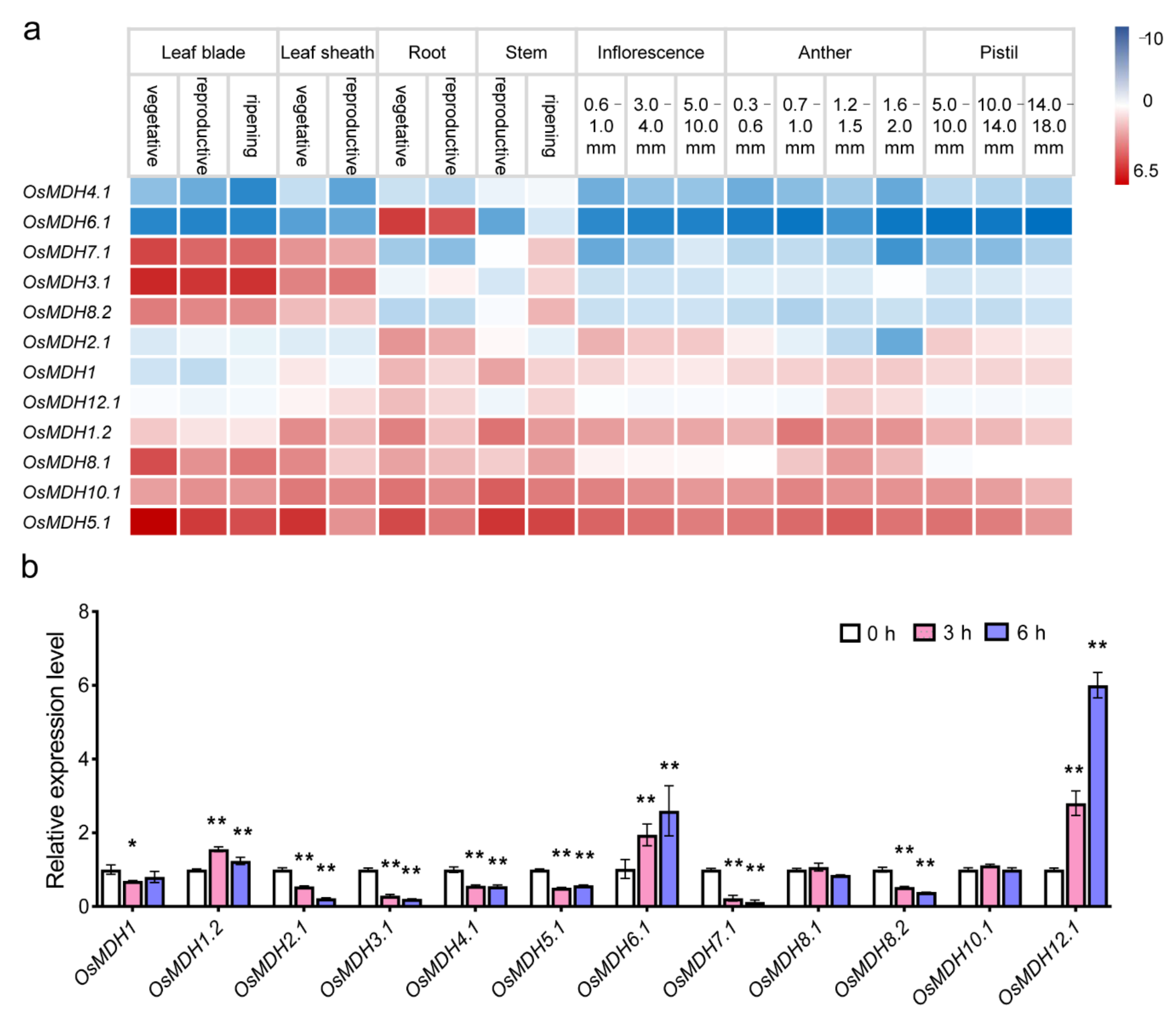
2.4. Expression Pattern of MDH Genes in Rice
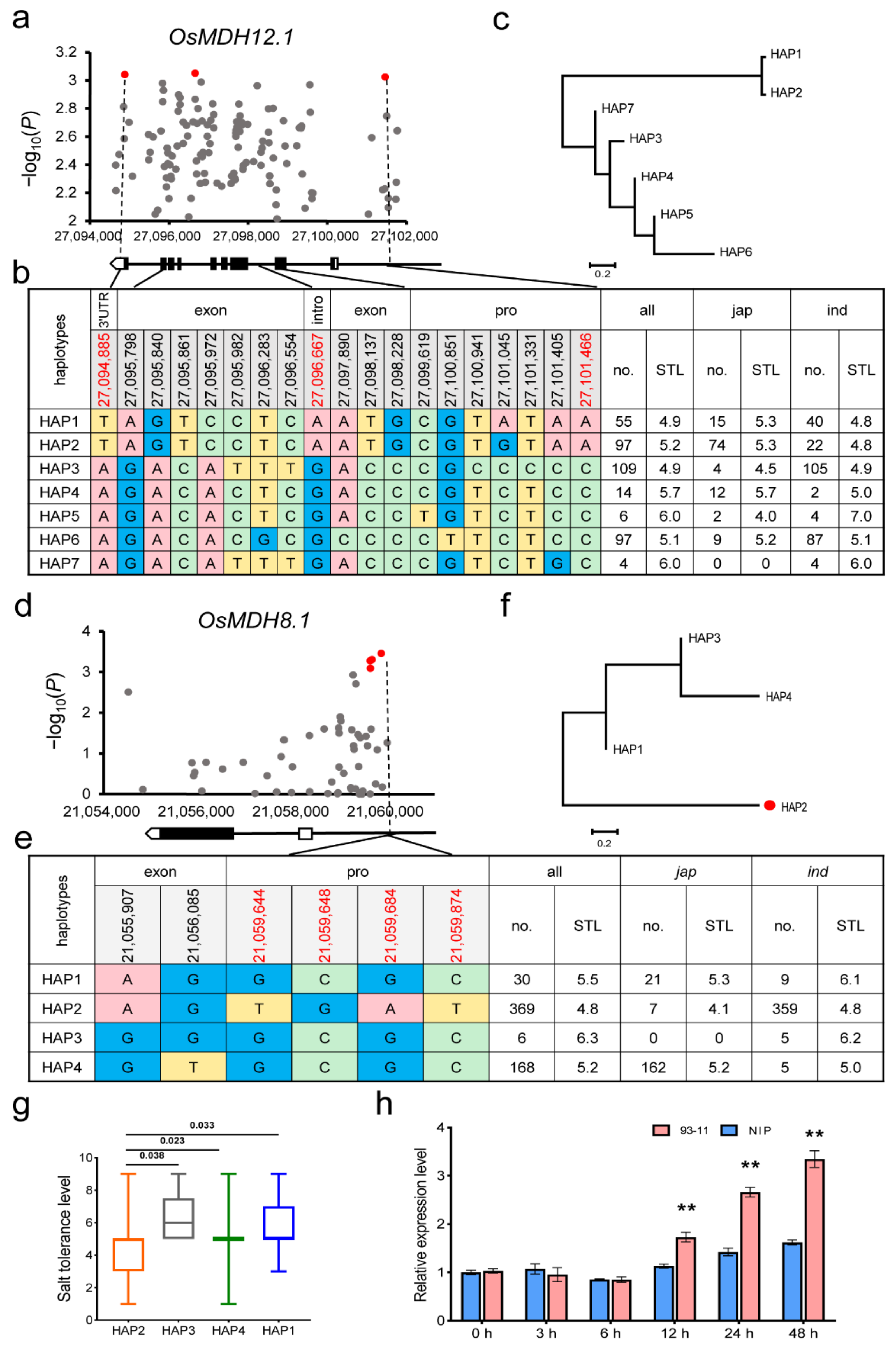
2.5. Gene-Based Association and Haplotype Analysis of OsMDH Genes
3. Discussion
4. Materials and Methods
4.1. Identification of the MDH Gene Family in Rice
4.2. Construction of Phylogenetic Tree of the MDH Gene Family
4.3. Structure and Motif Analysis of the MDH Gene Family in Rice
4.4. Duplication Analysis of OsMDH
4.5. Plant Materials and Salt Stress Treatment
4.6. Transcriptomic Analysis, RNA Extraction and qRT-PCR Analysis
4.7. Association Analysis of Candidate OsMDH Genes
4.8. Haplotype Analysis of Candidate Genes
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Scheibe, R. Malate valves to balance cellular energy supply. Physiol. Plant. 2004, 120, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Selinski, J.; König, N.; Wellmeyer, B.; Hanke, G.T.; Linke, V.; Neuhaus, H.E.; Scheibe, R. The plastid-localized NAD-dependent malate dehydrogenase is crucial for energy homeostasis in developing Arabidopsis thaliana seeds. Mol. Plant 2014, 7, 170–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomaz, T.; Bagard, M.; Pracharoenwattana, I.; Lindén, P.; Lee, C.P.; Carroll, A.J.; Ströher, E.; Smith, S.M.; Gardeström, P.; Millar, A.H. Mitochondrial malate dehydrogenase lowers leaf respiration and alters photorespiration and plant growth in Arabidopsis. Plant Physiol. 2010, 154, 1143–1157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, Y.; Dong, Q.; Zhai, H.; You, C.; Hao, Y. The functions of an apple cytosolic malate dehydrogenase gene in growth and tolerance to cold and salt stresses. Plant Physiol. Biochem. 2011, 49, 257–264. [Google Scholar] [CrossRef]
- Wang, Z.; Li, Q.; Ge, X.; Yang, C.; Luo, X.; Zhang, A.; Xiao, J.; Tian, Y.; Xia, G.; Chen, X.; et al. The mitochondrial malate dehydrogenase 1 gene GhmMDH1 is involved in plant and root growth under phosphorus deficiency conditions in cotton. Sci. Rep.-UK 2015, 5, 10343. [Google Scholar] [CrossRef]
- Sun, Q.; Yamada, T.; Han, Y.; Takano, T. Influence of salt stress on C4 photosynthesis in Miscanthus sinensis anderss. Plant Biol. 2021, 23, 44–56. [Google Scholar] [CrossRef]
- Beeler, S.; Liu, H.; Stadler, M.; Schreier, T.; Eicke, S.; Lue, W.; Truernit, E.; Zeeman, S.C.; Chen, J.; Kötting, O. Plastidial NAD-dependent malate dehydrogenase is critical for embryo development and heterotrophic metabolism in Arabidopsis. Plant Physiol. 2014, 164, 1175–1190. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Fu, Z.; Zhang, H.; Tian, R.; Yang, H.; Sun, C.; Wang, L.; Zhang, W.; Guo, Z.; Zhang, X.; et al. Cytosolic malate dehydrogenase 4 modulates cellular energetics and storage reserve accumulation in maize endosperm. Plant Biotechnol. J. 2020, 18, 2420–2435. [Google Scholar] [CrossRef]
- Teng, X.; Zhong, M.; Zhu, X.; Wang, C.; Ren, Y.; Wang, Y.; Zhang, H.; Jiang, L.; Wang, D.; Hao, Y.; et al. FLOURY ENDOSPERM16 encoding a NAD-dependent cytosolic malate dehydrogenase plays an important role in starch synthesis and seed development in rice. Plant Biotechnol. J. 2019, 17, 1914–1927. [Google Scholar] [CrossRef] [Green Version]
- Tesfaye, M.; Temple, S.J.; Allan, D.L.; Vance, C.P.; Samac, D.A. Overexpression of malate dehydrogenase in transgenic alfalfa enhances organic acid synthesis and confers tolerance to aluminum. Plant Physiol. 2001, 127, 1836–1844. [Google Scholar] [CrossRef]
- Song, J.; Zou, X.; Liu, P.; Cardoso, J.A.; Schultze-Kraft, R.; Liu, G.; Luo, L.; Chen, Z. Differential expressions and enzymatic properties of malate dehydrogenases in response to nutrient and metal stresses in Stylosanthes guianensis. Plant Physiol. Biochem. 2022, 170, 325–337. [Google Scholar] [CrossRef]
- Nan, N.; Wang, J.; Shi, Y.; Qian, Y.; Jiang, L.; Huang, S.; Liu, Y.; Wu, Y.; Liu, B.; Xu, Z.Y. Rice plastidial NAD-dependent malate dehydrogenase 1 negatively regulates salt stress response by reducing the vitamin B6 content. Plant Biotechnol. J. 2019, 18, 172–184. [Google Scholar] [CrossRef]
- Kumar, R.G.; Shah, K.; Dubey, R.S. Salinity induced behavioural changes in malate dehydrogenase and glutamate dehydrogenase activities in rice seedlings of differing salt tolerance. Plant Sci. 2000, 156, 23–34. [Google Scholar] [CrossRef]
- Yang, Y.; Guo, Y. Elucidating the molecular mechanisms mediating plant salt-stress responses. New Phytol. 2018, 217, 523–539. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zhao, H.; Qin, H.; Li, Z.; Liu, H.; Wang, J.; Zhang, H.; Quan, R.; Huang, R.; Zhang, Z. The synthesis of ascorbic acid in rice roots plays an important role in the salt tolerance of rice by scavenging ROS. Int. J. Mol. Sci. 2018, 19, 3347. [Google Scholar] [CrossRef] [Green Version]
- Gulen, H.; Turhan, E.; Eris, A. Changes in peroxidase activities and soluble proteins in strawberry varieties under salt-stress. Acta Physiol. Plant. 2006, 28, 109–116. [Google Scholar] [CrossRef]
- Wang, Q.J.; Sun, H.; Dong, Q.L.; Sun, T.Y.; Jin, Z.X.; Hao, Y.J.; Yao, Y.X. The enhancement of tolerance to salt and cold stresses by modifying the redox state and salicylic acid content via thecytosolic malate dehydrogenase gene in transgenic apple plants. Plant Biotechnol. J. 2016, 14, 1986–1997. [Google Scholar] [CrossRef] [Green Version]
- Kandoi, D.; Mohanty, S.; Tripathy, B.C. Overexpression of plastidic maize NADP-malate dehydrogenase (ZmNADP-MDH) in Arabidopsis thaliana confers tolerance to salt stress. Protoplasma 2018, 255, 547–563. [Google Scholar] [CrossRef]
- Schreier, T.B.; Cléry, A.; Schläfli, M.; Galbier, F.; Stadler, M.; Demarsy, E.; Albertini, D.; Maier, B.A.; Kessler, F.; Hörtensteiner, S.; et al. Plastidial NAD-dependent malate dehydrogenase: A moonlighting protein involved in early chloroplast development through its interaction with an FtsH12-FtsHi protease complex. Plant Cell 2018, 30, 1745–1769. [Google Scholar] [CrossRef] [Green Version]
- Ma, B.; Yuan, Y.; Gao, M.; Xing, L.; Li, C.; Li, M.; Ma, F. Genome-wide Identification, Classification, Molecular Evolution and Expression Analysis of Malate Dehydrogenases in Apple. Int. J. Mol. Sci. 2018, 19, 3312. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Zhang, J.; Zhang, C.; Wang, S.; Yang, M. Genome-wide investigation of malate dehydrogenase gene family in poplar (Populus trichocarpa) and their expression analysis under salt stress. Acta Physiol. Plant. 2021, 43, 28. [Google Scholar] [CrossRef]
- Imran, M.; Tang, K.; Liu, J. Comparative Genome-wide analysis of the malate dehydrogenase gene families in cotton. PLoS ONE 2016, 11, e166341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, J.; Warburton, M.; Crouch, J. Association mapping for enhancing maize (Zea mays L.) genetic improvement. Crop Sci. 2011, 51, 433–449. [Google Scholar] [CrossRef]
- Ren, Z.; Gao, J.; Li, L.; Cai, X.; Huang, W.; Chao, D.; Zhu, M.; Wang, Z.; Luan, S.; Lin, H. A rice quantitative trait locus for salt tolerance encodes a sodium transporter. Nat. Genet. 2005, 37, 1141–1146. [Google Scholar] [CrossRef]
- Cui, L.; Shan, J.; Shi, M.; Gao, J.; Lin, H. DCA1 acts as a transcriptional co-activator of DST and contributes to drought and salt tolerance in rice. PLoS Genet. 2015, 11, e1005617. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Yang, B.; He, Y.; Zhan, C.; Cheng, Y.; Zhang, J.; Zhang, H.; Cheng, J.; Wang, Z. A quantitative trait locus, qSE3, promotes seed germination and seedling establishment under salinity stress in rice. Plant J. 2019, 97, 1089–1104. [Google Scholar] [CrossRef] [Green Version]
- Mishra, S.; Singh, B.; Misra, P.; Rai, V.; Singh, N.K. Haplotype distribution and association of candidate genes with salt tolerance in Indian wild rice germplasm. Plant Cell Rep. 2016, 35, 2295–2308. [Google Scholar] [CrossRef]
- Mishra, S.; Singh, B.; Panda, K.; Singh, B.P.; Singh, N.; Misra, P.; Rai, V.; Singh, N.K. Association of SNP haplotypes of HKT family genes with salt tolerance in indian wild rice germplasm. Rice 2016, 9, 15. [Google Scholar] [CrossRef] [Green Version]
- Yuan, J.; Wang, X.; Zhao, Y.; Khan, N.U.; Zhao, Z.; Zhang, Y.; Wen, X.; Tang, F.; Wang, F.; Li, Z. Genetic basis and identification of candidate genes for salt tolerance in rice by GWAS. Sci. Rep.-UK 2020, 10, 9958. [Google Scholar] [CrossRef]
- Li, B. Identification of genes conferring plant salt tolerance using GWAS: Current success and perspectives. Plant Cell Physiol. 2020, 61, 1419–1426. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Liu, H.J.; Yan, J.; Tian, F. Natural Variation in Crops: Realized understanding, continuing promise. Annu. Rev. Plant Biol. 2021, 72, 357–385. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, A.K.; Desai, P.V.; Pradhan, A.; Khan, S.I.; Avery, M.A.; Walker, L.A.; Tekwani, B.L. An alpha-proteobacterial type malate dehydrogenase may complement LDH function in Plasmodium falciparum. Cloning and biochemical characterization of the enzyme. Eur. J. Biochem. 2004, 274, 3488–3502. [Google Scholar] [CrossRef] [PubMed]
- Moore, R.C.; Purugganan, M.D. The early stages of duplicate gene evolution. Proc. Natl. Acad. Sci. USA 2003, 100, 15682–15687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stein, J.C.; Yu, Y.; Copetti, D.; Zwickl, D.J.; Zhang, L.; Zhang, C.; Chougule, K.; Gao, D.; Iwata, A.; Goicoechea, J.L.; et al. Genomes of 13 domesticated and wild rice relatives highlight genetic conservation, turnover and innovation across the genus Oryza. Nat. Genet. 2018, 50, 285–296. [Google Scholar] [CrossRef]
- Qin, P.; Lu, H.; Du, H.; Wang, H.; Chen, W.; Chen, Z.; He, Q.; Ou, S.; Zhang, H.; Li, X.; et al. Pan-genome analysis of 33 genetically diverse rice accessions reveals hidden genomic variations. Cell 2021, 184, 3542–3558. [Google Scholar] [CrossRef]
- Todeschini, A.; Georges, A.; Veitia, R.A. Transcription factors: Specific DNA binding and specific gene regulation. Trends Genet. 2014, 30, 211–219. [Google Scholar] [CrossRef]
- Singh, B.P.; Jayaswal, P.K.; Singh, B.; Singh, P.K.; Kumar, V.; Mishra, S.; Singh, N.; Panda, K.; Singh, N.K. Natural allelic diversity in OsDREB1F gene in the Indian wild rice germplasm led to ascertain its association with drought tolerance. Plant Cell Rep. 2015, 34, 993–1004. [Google Scholar] [CrossRef]
- Zhu, N.; Cheng, S.; Liu, X.; Du, H.; Dai, M.; Zhou, D.; Yang, W.; Zhao, Y. The R2R3-type MYB gene OsMYB91 has a function in coordinating plant growth and salt stress tolerance in rice. Plant Sci. 2015, 236, 146–156. [Google Scholar] [CrossRef]
- Li, J.; Han, G.; Sun, C.; Sui, N. Research advances of MYB transcription factors in plant stress resistance and breeding. Plant Signal. Behav. 2019, 14, 1613131. [Google Scholar] [CrossRef]
- Wang, X.; Niu, Y.; Zheng, Y. Multiple functions of MYB transcription factors in abiotic stress responses. Int. J. Mol. Sci. 2021, 22, 6125. [Google Scholar] [CrossRef]
- Zelm, E.; Zhang, Y.; Testerink, C. Salt tolerance mechanisms of plants. Annu. Rev. Plant Biol. 2020, 71, 403–433. [Google Scholar] [CrossRef] [Green Version]
- Ponce, K.S.; Guo, L.; Leng, Y.; Meng, L.; Ye, G. Advances in sensing, response and regulation mechanism of salt tolerance in rice. Int. J. Mol. Sci. 2021, 22, 2254. [Google Scholar] [CrossRef]
- Kamran, M.; Parveen, A.; Ahmar, S.; Malik, Z.; Hussain, S.; Chattha, M.S.; Saleem, M.H.; Adil, M.; Heidari, P.; Chen, J. An Overview of hazardous impacts of soil salinity in crops, tolerance mechanisms, and amelioration through selenium supplementation. Int. J. Mol. Sci. 2020, 21, 148. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Mao, B.; Yuan, D.; Chu, C.; Duan, M. Salt tolerance in rice: Physiological responses and molecular mechanisms. Crop J. 2022, 10, 13–25. [Google Scholar] [CrossRef]
- Yıldız, S.; Okay, A.; Büyük, İ. Defining the roles of PvMDH genes in response to salt stress and detailed characterization of the gene family. J. Plant Biochem. Biot. 2022, 31, 380–393. [Google Scholar] [CrossRef]
- Conant, G.C.; Wolfe, K.H. Turning a hobby into a job: How duplicated genes find new functions. Nat. Rev. Genet. 2008, 9, 938–950. [Google Scholar] [CrossRef]
- Liao, Y.; Zou, H.; Wei, W.; Hao, Y.; Tian, A.; Huang, J.; Liu, Y.; Zhang, J.; Chen, S. Soybean GmbZIP44, GmbZIP62 and GmbZIP78 genes function as negative regulator of ABA signaling and confer salt and freezing tolerance in transgenic Arabidopsis. Planta 2008, 228, 225–240. [Google Scholar] [CrossRef]
- Cai, S.G.; Chen, G.; Wang, Y.Y.; Huang, Y.Q.; Marchant, D.B.; Wang, Y.Z.; Yang, Q.; Dai, F.; Hills, A.A.; Franks, P.J.; et al. Evolutionary conservation of ABA signaling for stomatal closure. Plant Physiol. 2017, 174, 732–747. [Google Scholar] [CrossRef]
- Belda-Palazón, B.; Adamo, M.; Valerio, C.; Ferreira, L.J.; Confraria, A.; Reis-Barata, D.; Rodrigues, A.; Meyer, C.; Rodriguez, P.L.; Baena-González, E. A dual function of SnRK2 kinases in the regulation of SnRK1 and plant growth. Nat. Plants 2020, 6, 1345–1353. [Google Scholar] [CrossRef]
- Ito, Y.; Katsura, K.; Maruyama, K.; Taji, T.; Kobayashi, M.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Functional analysis of rice DREB1/CBF-type transcription factors involved in cold-responsive gene expression in transgenic rice. Plant Cell Physiol. 2006, 47, 141–153. [Google Scholar] [CrossRef] [Green Version]
- Jin, X.F.; Xiong, A.S.; Peng, R.H.; Liu, J.G.; Gao, F.; Chen, J.M.; Yao, Q.H. OsAREB1, an ABRE-binding protein responding to ABA and glucose, has multiple functions in Arabidopsis. BMB Rep. 2010, 43, 34–39. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Wang, X.; Wang, H.; Xin, H.; Yang, X.; Yan, J.; Li, J.; Tran, L.S.; Shinozaki, K.; Yamaguchi-Shinozaki, K.; et al. Genome-wide analysis of ZmDREB genes and their association with natural variation in drought tolerance at seedling stage of Zea mays L. PLoS Genet. 2013, 9, e1003790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, W.; Ye, J.; Yao, X.; Zhao, P.; Xuan, W.; Tian, Y.; Zhang, Y.; Xu, S.; An, H.; Chen, G.; et al. Genome-wide associated study identifies NAC42-activated nitrate transporter conferring high nitrogen use efficiency in rice. Nat. Commun. 2019, 10, 5279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, H.; Li, S.; Chen, B.; Jian, C.; Mei, F.; Zhang, Y.; Li, F.; Chen, N.; Li, T.; Du, L.; et al. Variation in cis-regulation of a NAC transcription factor contributes to drought tolerance in wheat. Mol. Plant 2022, 15, 276–292. [Google Scholar] [CrossRef]
- Jiang, H.; Hong, J.; Jiang, Y.; Yu, S.; Zhang, Y.; Shi, J.; Lin, W. Genome-wide association analysis identifies candidate genes regulating seed number per silique in Arabidopsis thaliana. Plants 2020, 9, 585. [Google Scholar] [CrossRef]
- Liu, X.; Gu, X.; Lu, H.; Liu, P.; Miao, H.; Bai, Y.; Zhang, S. Identification of novel loci and candidate genes for resistance to powdery mildew in a resequenced cucumber germplasm. Genes 2021, 12, 584. [Google Scholar] [CrossRef]
- Abbai, R.; Singh, V.K.; Nachimuthu, V.V.; Sinha, P.; Selvaraj, R.; Vipparla, A.K.; Singh, A.K.; Singh, U.M.; Varshney, R.K.; Kumar, A. Haplotype analysis of key genes governing grain yield and quality traits across 3KRG panel reveals scope for the development of tailor-made rice with enhanced genetic gains. Plant Biotechnol. J. 2019, 17, 1612–1622. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Kumar, S.; Singh, K.; Prasad, M.; Thakur, J.K. Designing a mini-core collection effectively representing 3004 diverse rice accessions. Plant Commun. 2020, 1, 100049. [Google Scholar] [CrossRef]
- Park, C.; Chen, S.; Shirsekar, G.; Zhou, B.; Khang, C.H.; Songkumarn, P.; Afzal, A.J.; Ning, Y.; Wang, R.; Bellizzi, M.; et al. The magnaporthe oryzae effector AvrPiz-t targets the RING E3 ubiquitin ligase APIP6 to suppress pathogen-associated molecular pattern-triggered immunity in rice. Plant Cell 2012, 24, 4748–4762. [Google Scholar] [CrossRef] [Green Version]
- Bradbury, P.J.; Zhang, Z.; Kroon, D.E.; Casstevens, T.M.; Ramdoss, Y.; Buckler, E.S. TASSEL: Software for association mapping of complex traits in diverse samples. Bioinformatics 2007, 23, 2633–2635. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Gene Name | Chr | Start | End | Genomic Sequence Length (bp) | CDS (bp) | Protein Length (aa) | MW (kDa) | Isoelectric Point (PI) | Subcellular Localization |
|---|---|---|---|---|---|---|---|---|---|---|
| LOC_Os01g61380 | OsMDH1 [12] | 1 | 35499017 | 35501765 | 2749 | 1191 | 396 | 41.79 | 7.90 | cytoplasmic |
| LOC_Os01g46070 | OsMDH1.2 | 1 | 26190752 | 26194517 | 3766 | 1023 | 340 | 35.46 | 8.74 | cytoplasmic |
| LOC_Os02g01510 | OsMDH2.1 | 2 | 295302 | 299174 | 3873 | 1179 | 392 | 42.72 | 7.29 | endoplasmic reticulum |
| LOC_Os03g56280 | OsMDH3.1 | 3 | 32086001 | 32089685 | 3685 | 1065 | 354 | 37.02 | 8.06 | cytoplasmic |
| LOC_Os04g46560 | OsMDH4.1 | 4 | 27605166 | 27608347 | 3182 | 1059 | 352 | 38.30 | 7.22 | endoplasmic reticulum |
| LOC_Os05g49880 | OsMDH5.1 | 5 | 28617595 | 28621585 | 3991 | 1023 | 340 | 35.44 | 8.30 | nuclear |
| LOC_Os06g01590 | OsMDH6.1 | 6 | 346985 | 348516 | 1532 | 1083 | 360 | 38.72 | 8.46 | nuclear |
| LOC_Os07g43700 | OsMDH7.1 | 7 | 26153825 | 26156006 | 2182 | 1215 | 404 | 42.22 | 9.03 | nuclear |
| LOC_Os08g33720 | OsMDH8.1 | 8 | 21054659 | 21057561 | 2903 | 1194 | 397 | 41.54 | 7.54 | cytoplasmic |
| LOC_Os08g44810 | OsMDH8.2 | 8 | 28141042 | 28146270 | 5229 | 1302 | 433 | 47.01 | 7.34 | endoplasmic reticulum |
| LOC_Os10g33800 | OsMDH10.1(FLO16) [9] | 10 | 17913818 | 17917850 | 4033 | 999 | 332 | 35.57 | 5.97 | endoplasmic reticulum |
| LOC_Os12g43630 | OsMDH12.1 | 12 | 27094647 | 27099351 | 4705 | 1071 | 356 | 37.39 | 7.99 | cytoplasmic |
| Gene ID | Gene Name | Polymorphic Number | GLM (p ≤ 0.01) | GLM (p ≤ 0.001) | CMLM (p ≤ 0.01) | CMLM (p ≤ 0.001) |
|---|---|---|---|---|---|---|
| LOC_Os01g61380 | OsMDH1 | 9 | 0 | 0 | 0 | 0 |
| LOC_Os01g46070 | OsMDH1.2 | 9 | 0 | 0 | 0 | 0 |
| LOC_Os02g01510 | OsMDH2.1 | 19 | 0 | 0 | 0 | 0 |
| LOC_Os03g56280 | OsMDH3.1 | 40 | 0 | 0 | 0 | 0 |
| LOC_Os04g46560 | OsMDH4.1 | 63 | 6 | 0 | 0 | 0 |
| LOC_Os05g49880 | OsMDH5.1 | 29 | 0 | 0 | 0 | 0 |
| LOC_Os06g01590 | OsMDH6.1 | 49 | 14 | 0 | 0 | 0 |
| LOC_Os07g43700 | OsMDH7.1 | 33 | 0 | 0 | 0 | 0 |
| LOC_Os08g33720 | OsMDH8.1 | 56 | 6 | 4 | 2 | 0 |
| LOC_Os08g44810 | OsMDH8.2 | 35 | 4 | 0 | 0 | 0 |
| LOC_Os10g33800 | OsMDH10.1 | 56 | 0 | 0 | 0 | 0 |
| LOC_Os12g43630 | OsMDH12.1 | 159 | 134 | 3 | 0 | 0 |
| Gene ID | Primer Name | Sequence of Forward Primer | Sequence of Reverse Primer | Annotation |
|---|---|---|---|---|
| LOC_Os01g61380 | OsMDH1 | CGAAAGCTGGTGCTGGATCTG | CACGGAGGGATGACTCAACA | OsMDH1 [12] |
| LOC_Os01g46070 | OsMDH1.2 | AACGCCGGCATCGTTAAGAAC | GGGTTGCTGATCATGTTGACAAG | lactate/malate dehydrogenase |
| LOC_Os02g01510 | OsMDH2.1 | CGGCACCAACCTCGACTC | CGTGCTCTCCCACCATGTAC | lactate/malate dehydrogenase |
| LOC_Os03g56280 | OsMDH3.1 | TGGCGTTGTGGAATGTTCA | GGCTCCAGCACGACCTAAC | lactate/malate dehydrogenase |
| LOC_Os04g46560 | OsMDH4.1 | CGAGGCTGAGGCGTTCAAG | GCAGAGGCCTGGGATTTGTAG | lactate/malate dehydrogenase |
| LOC_Os05g49880 | OsMDH5.1 | GCCAGCTTTCCGAGTTTGAGAAG | GTTCGCGTGAGCAAACTTGATG | lactate/malate dehydrogenase |
| LOC_Os06g01590 | OsMDH6.1 | AGCGCGTACGAGGTGATCAAG | GATGCTGGCGACGGAGTAG | lactate/malate dehydrogenase |
| LOC_Os07g43700 | OsMDH7.1 | GCGCTGCACCTGTACGAC | CGTGTTGCAGTGTCCAAGATC | lactate/malate dehydrogenase |
| LOC_Os08g44810 | OsMDH8.1 | GCAGAGGACATCGTGTTCAGTA | CGTCCATTGCCACATCTTTAACTAG | lactate/malate dehydrogenase |
| LOC_Os08g33720 | OsMDH8.2 | GCTGACCTTGAGGGAGTGA | TCGATGCCCTTCTCGATACTG | lactate/malate dehydrogenase |
| LOC_Os10g33800 | OsMDH10.1 | AGCAAACACCAACGCTCTCATC | TGCCCTGTTGTGGTCAAGA | FLO16 [9] |
| LOC_Os12g436301 | OsMDH12.1 | GCCAGCCACAGTTGGAAA | CCCAGGCTTACGAGGAACA | lactate/malate dehydrogenase |
| LOC_Os03g13170 | Ubi | AACCAGCTGAGGCCCAAGA | ACGATTGATTTAACCAGTCCATGA | Ubq (ubiquitin fusion protein) [60] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Wang, Y.; Sun, X.; Yuan, J.; Zhao, Z.; Gao, J.; Wen, X.; Tang, F.; Kang, M.; Abliz, B.; et al. Genome-Wide Identification of MDH Family Genes and Their Association with Salt Tolerance in Rice. Plants 2022, 11, 1498. https://doi.org/10.3390/plants11111498
Zhang Y, Wang Y, Sun X, Yuan J, Zhao Z, Gao J, Wen X, Tang F, Kang M, Abliz B, et al. Genome-Wide Identification of MDH Family Genes and Their Association with Salt Tolerance in Rice. Plants. 2022; 11(11):1498. https://doi.org/10.3390/plants11111498
Chicago/Turabian StyleZhang, Yanhong, Yulong Wang, Xingming Sun, Jie Yuan, Zhiqiang Zhao, Jie Gao, Xiaorong Wen, Fusen Tang, Mintai Kang, Buhaliqem Abliz, and et al. 2022. "Genome-Wide Identification of MDH Family Genes and Their Association with Salt Tolerance in Rice" Plants 11, no. 11: 1498. https://doi.org/10.3390/plants11111498