
Genome-Wide Identification and Functional Differentiation of Fatty Acid Desaturase Genes in Olea europaea L.

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Genome-Wide Identification of FAD Genes in Olive
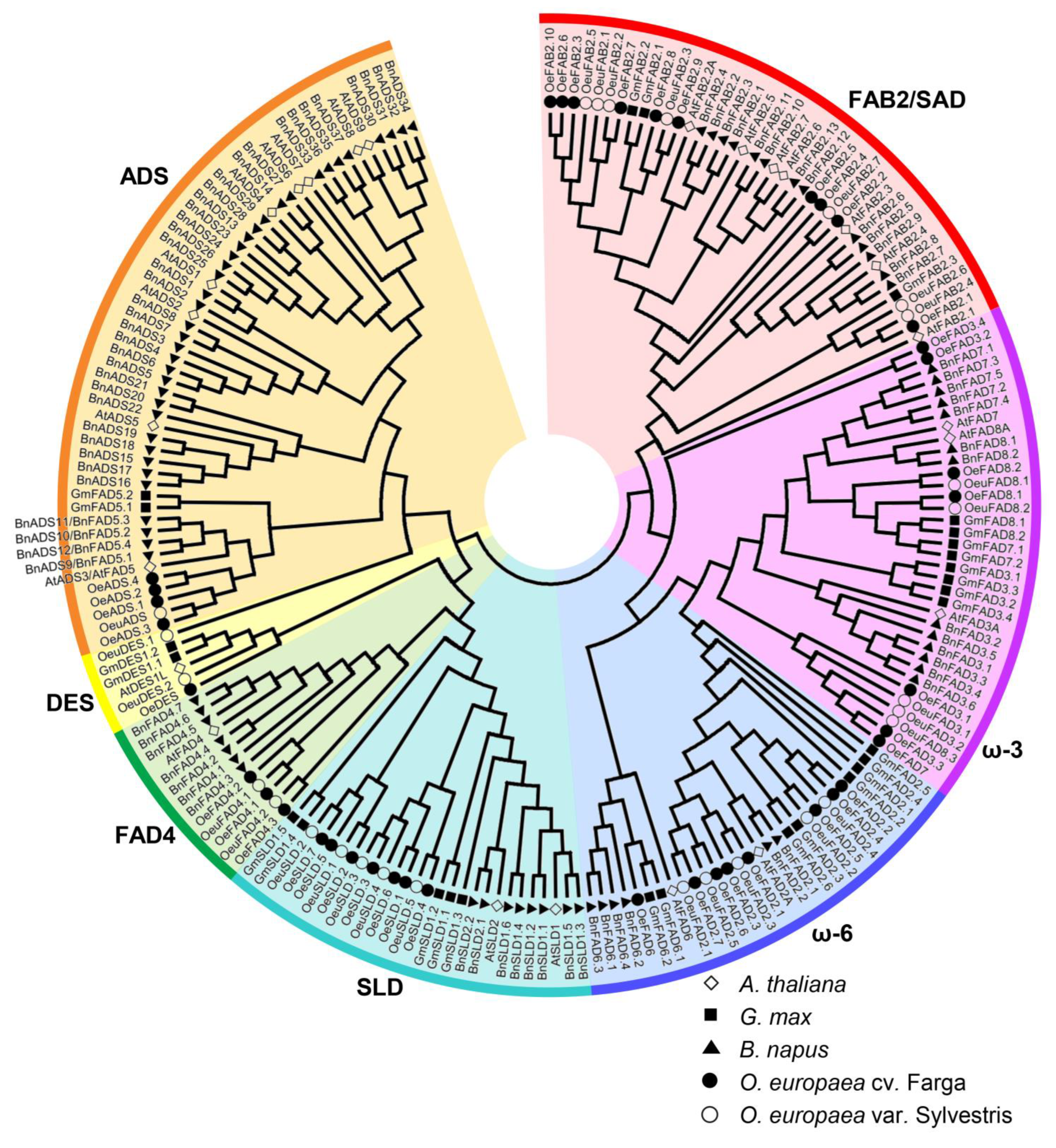
2.2. Phylogenetic Analysis of FAD Genes in Different Species
2.3. Gene Characters and Protein Subcellular Localization
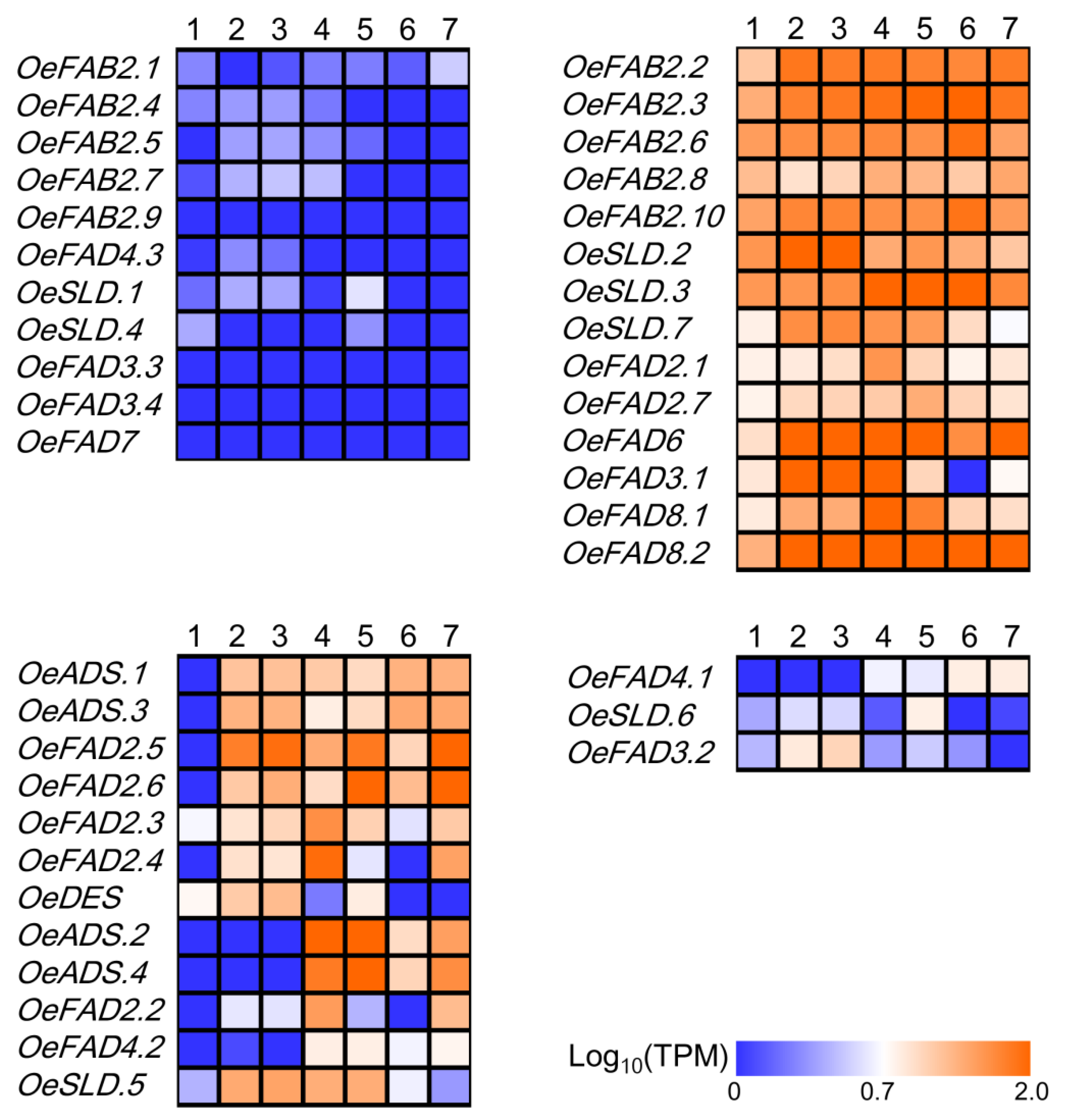
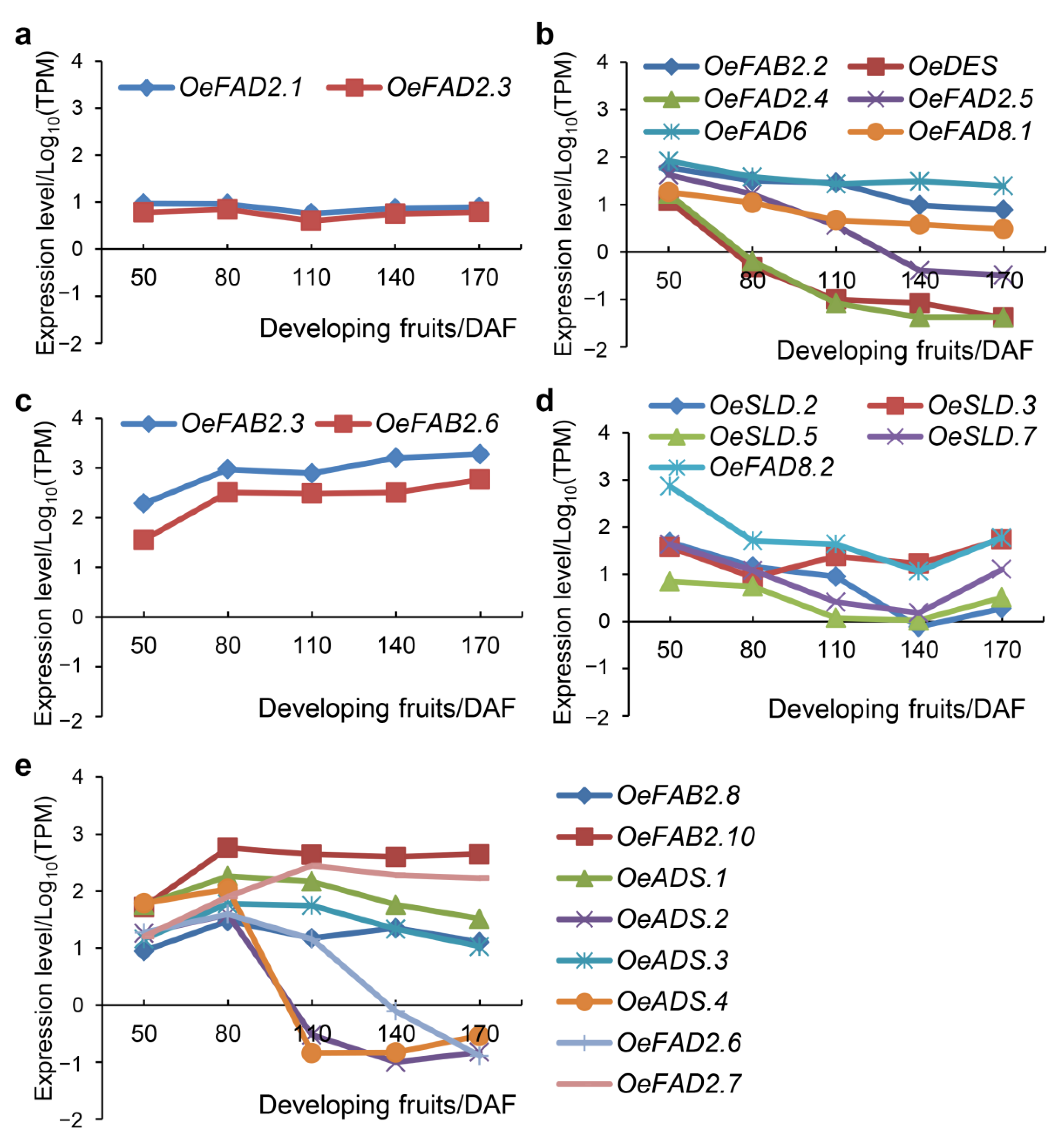
2.4. The Roles of FAD Genes in Olive Morphology
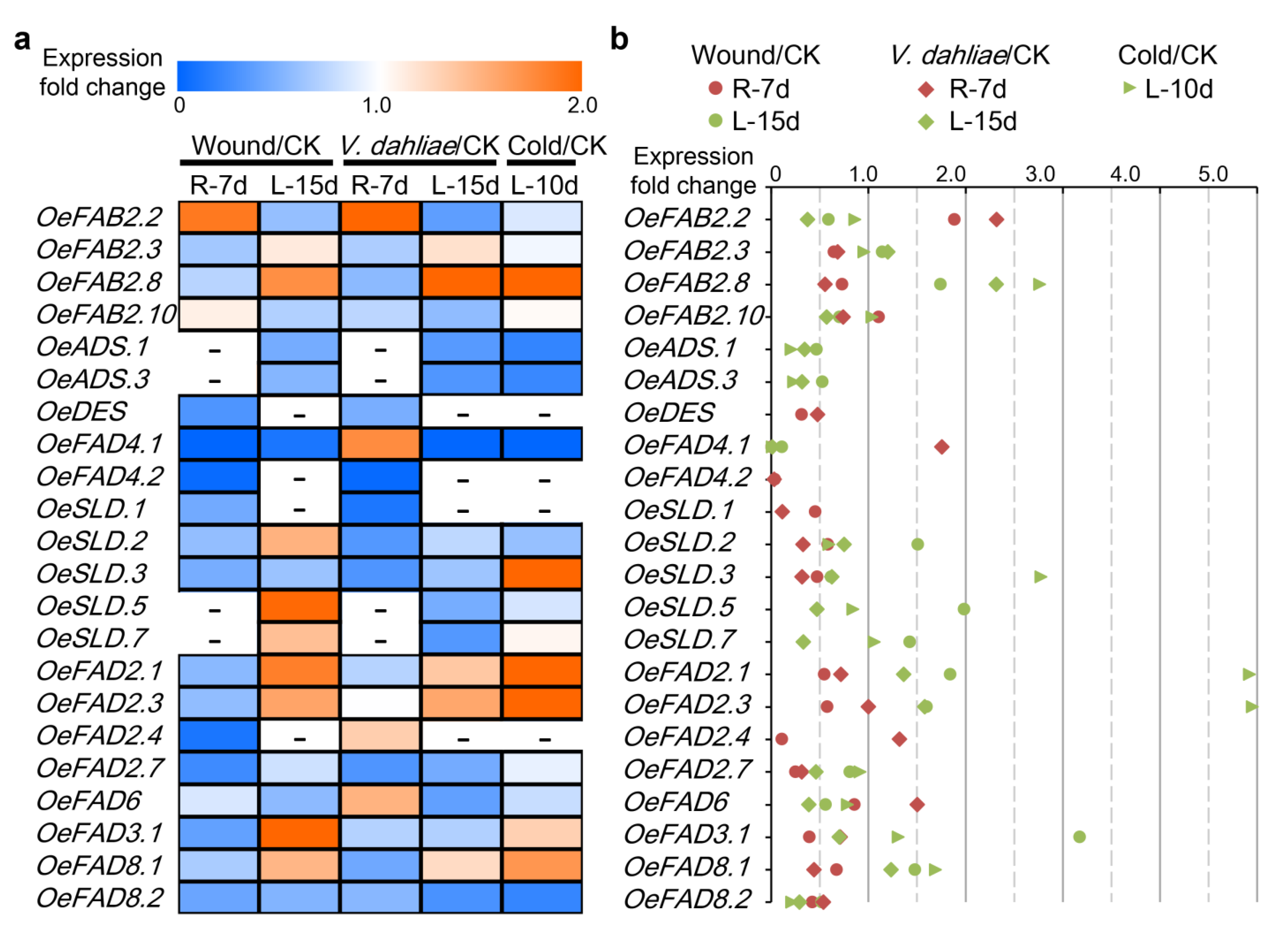
2.5. Stress Response of FAD Genes in Olive
3. Discussion
3.1. FAD Genes in Different Plants
3.2. Functional Differentiation of FAD Genes in Olive
4. Materials and Methods
4.1. Database Search and FAD Genes Retrieval
4.2. Phylogenetic Analysis
4.3. Physicochemical Character and Subcellular Localization Prediction
4.4. Expression Analysis of FAD Genes in Olive
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kaniewski, D.; Van Campo, E.; Boiy, T.; Terral, J.F.; Khadari, B.; Besnard, G. Primary domestication and early uses of the emblematic olive tree: Palaeobotanical, historical and molecular evidences from the middle east. Biol. Rev. 2012, 87, 885–899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Green, P.S.; Wickens, G.E. The Olea Europaea Complex; Edinburg University Press: Edinburg, UK, 1989. [Google Scholar]
- Green, P.S. A revision of Olea L (Oleaceae). Kew Bull. 2002, 57, 91–140. [Google Scholar] [CrossRef]
- Dyer, J.M.; Stymne, S.; Green, A.G.; Carlsson, A.S. High value oils from plants. Plant J. 2008, 54, 640–655. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Yin, X.; Li, X.; Yang, S.; Ferguson, I.B.; Chen, K. Lipoxygenase gene expression in ripening kiwifruit in relation to ethylene and aroma production. J. Agric. Food Chem. 2009, 57, 2875–2881. [Google Scholar] [CrossRef]
- Pirtle, I.L.; Kongcharoensuntorn, W.; Nampaisansuk, M.; Knesek, J.E.; Chapman, K.D.; Pirtle, R.M. Molecular cloning and functional expression of the gene for a cotton Δ-12 fatty acid desaturase (FAD2). Biochim. Biophys. Acta 2001, 1522, 122–129. [Google Scholar] [CrossRef]
- Venugopal, S.C.; Chanda, B.; Vaillancourt, L.; Kachroo, A.; Kachroo, P. The common metabolite glycerol-3-phosphate is a novel regulator of plant defense signaling. Plant Signal. Behav. 2009, 4, 746–749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slabas, A.R.; Fawcett, T. The biochemistry and molecular biology of plant lipid biosynthesis. Plant Mol. Biol. 1992, 19, 169–191. [Google Scholar] [CrossRef]
- Wang, X.; Long, Y.; Yin, Y.; Zhang, C.; Gan, L.; Liu, L.; Yu, L.; Meng, J.; Li, M. New insights into the genetic networks affecting seed fatty acid concentrations in Brassica napus. BMC Plant Biol. 2015, 15, 91. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, Y.; Nagano, Y. Plant acetyl-CoA carboxylase: Structure, biosynthesis, regulation, and gene manipulation for plant breeding. Biosci. Biotechnol. Biochem. 2004, 68, 1175–1184. [Google Scholar] [CrossRef]
- Du, C.; Chen, Y.; Wang, K.; Yang, Z.; Zhao, C.; Jia, Q.; Taylor, D.; Zhang, M. Strong co-suppression impedes an increase in polyunsaturated fatty acids in seeds overexpressing. FAD2. J. Exp. Bot. 2019, 70, 985–994. [Google Scholar] [CrossRef]
- The Arabidopsis Information Resource. Available online: http://www.arabidopsis.org (accessed on 25 April 2022).
- Chi, X.; Yang, Q.; Lu, Y.; Wang, J.; Zhang, Q.; Pan, L.; Chen, M.; He, Y.; Yu, S. Genome-wide analysis of fatty acid desaturases in soybean (Glycine max). Plant Mol. Biol. Rep. 2011, 29, 769–783. [Google Scholar] [CrossRef] [Green Version]
- Xue, Y.; Chen, B.; Wang, R.; Win, A.N.; Li, J.; Chai, Y. Genome-wide survey and characterization of fatty acid desaturase gene family in Brassica napus and its parental species. Appl. Biochem. Biotechnol. 2018, 184, 582–598. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.; Chai, C.; Chen, B.; Shi, X.; Chai, Y. Whole-genome mining and in silico analysis of fad gene family in Brassica juncea. J. Plant Biochem Biotechnol. 2020, 29, 149–154. [Google Scholar] [CrossRef]
- Liu, K.; Zhao, S.; Wang, S.; Wang, H.; Zhang, Z. Identification and analysis of the FAD gene family in walnuts (Juglans regia L.) based on transcriptome data. BMC Genom. 2020, 21, 299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Jin, X.; Liu, Z.; Zhang, J.; Liu, W. Genome-wide identification of fad gene family and functional analysis of MsFAD3.1 involved in the accumulation of α-linolenic acid in alfalfa. Crop Sci. 2020, 61, 566–579. [Google Scholar] [CrossRef]
- Wallis, J.G.; Browse, J. Mutants of Arabidopsis reveal many roles for membrane lipids. Prog. Lipid Res. 2002, 41, 254–278. [Google Scholar] [CrossRef]
- Smith, M.A.; DaUk, M.; Ramadan, H.; Yang, H.; Seamons, L.E.; Haslam, R.P.; Beaudoin, F.; Ramirez-Erosa, I.; Forseille, L. Involvement of Arabidopsis Acyl-coenzyme a desaturase-like2 (At2g31360) in the biosynthesis of the very-long-chain monounsaturated fatty acid components of membrane lipids. Plant Physiol. 2013, 161, 81–96. [Google Scholar] [CrossRef] [Green Version]
- Ohlrogge, J.B.; Jaworski, J.G. Regulation of fatty acid synthesis. Annu. Rev. Plant Physiol. 1997, 48, 109–136. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Singh, S.P.; Green, A.G. High-stearic and high-oleic cottonseed oils produced by hairpin RNA-mediated post-transcriptional gene silencing. Plant Physiol. 2002, 129, 1732–1743. [Google Scholar] [CrossRef] [Green Version]
- Okuzaki, A.; Ogawa, T.; Koizuka, C.; Kaneko, K.; Inaba, M.; Imamura, J.; Koizuka, N. CRISPR/Cas9-mediated genome editing of the fatty acid desaturase 2 gene in Brassica napus. Plant Physiol. Biochem. 2018, 131, 63–69. [Google Scholar] [CrossRef]
- Kargiotidou, A.; Deli, D.; Galanopoulou, D.; Tsaftaris, A.; Farmaki, T. Low temperature and light regulate delta 12 fatty acid desaturases (FAD2) at a transcriptional level in cotton (Gossypium hirsutum). J. Exp. Bot. 2008, 59, 2043–2056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domínguez, T.; Hernández, M.L.; Pennycooke, J.C.; Jiménez, P.; Martinez-Rivas, J.M.; Sanz, C.; Stockinger, E.; SánchezSerrano, J.J.; Sanmartín, M. Increasing ω-3 desaturase expression in tomato results in altered aroma profile and enhanced resistance to cold stress. Plant Physiol. 2010, 153, 655–665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Z.; Wang, M.; Zhao, S.; Hu, J.; Lu, M. Changes in freezing tolerance in hybrid poplar caused by up- and down-regulation of PtFAD2 gene expression. Transgenic Res. 2010, 19, 647–654. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, H.; Sun, J.; Li, B.; Zhu, Q.; Chen, S.; Zhang, H. Arabidopsis fatty acid desaturase FAD2 is required for salt tolerance during seed germination and early seedling growth. PLoS ONE 2012, 7, e30355. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Barg, R.; Yin, M.; Gueta-Dahan, Y.; Leikin-Frenkel, A.; Salts, Y.; Shabtai, S.; Ben-Hayyim, G. Modulated fatty acid desaturation via overexpression of two distinct x-3 desaturases differentially alters tolerance to various abiotic stresses in transgenic tobacco cells and plants. Plant J. 2005, 44, 361–371. [Google Scholar] [CrossRef]
- Contreras, C.; Mariotti, R.; Mousavi, S.; Baldoni, L.; Guerrero, C.; Roka, L.; Cultrera, N.; Pierantozzi, P.; Maestri, D.; ·Gentili, L.; et al. Characterization and validation of olive FAD and SAD gene families: Expression analysis in different tissues and during fruit development. Mol. Biol. Rep. 2020, 47, 4345–4355. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Guo, L.; Zhang, J.; Xue, L.; Luo, Y.; Rao, G. Integrated analysis of fatty acid metabolism and transcriptome involved in olive fruit development to improve oil composition. Forests 2021, 12, 1773. [Google Scholar] [CrossRef]
- Niu, E.; Hu, W.; Ding, J.; Wang, W.; Romero, A.; Shen, G.; Zhu, S. GC-MS/LC-MS and transcriptome analyses revealed the metabolisms of fatty acid and flavonoid in olive fruits (Olea europaea L.). Sci. Hortic. 2022, 299, 111017. [Google Scholar] [CrossRef]
- Ayed, R.B.; Moreau, F.; Hlima, H.B.; Rebai, A.; Ercisli, S.; Kadoo, N.; Hanana, M.; Assouguem, A.; Ullah, R.; Ali, E.A. SNP discovery and structural insights into OeFAD2 unravelling high oleic/linoleic ratio in olive oil. Comput. Struct. Biotechnol. J. 2022, 20, 1229–1243. [Google Scholar] [CrossRef]
- Cruz, F.; Julca, I.; Gómez-Garrido, J.; Loska, D.; Marcet-Houben, M.; Cano, E.; Galán, B.; Frias, L.; Ribeca, P.; Derdak, S.; et al. Genome sequence of the olive tree, Olea europaea. Gigascience 2016, 5, 29. [Google Scholar] [CrossRef]
- Unver, T.; Wu, Z.; Sterck, L.; Turktas, M.; Lohaus, R.; Li, Z.; Yang, M.; He, L.; Deng, T.; Escalante, F.J.; et al. Genome of wild olive and the evolution of oil biosynthesis. Proc. Natl. Acad. Sci. USA 2017, 114, E9413–E9422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, G.; Zhang, J.; Liu, X.; Lin, C.; Xin, H.; Xue, L.; Wang, C. De novo assembly of a new Olea europaea genome accession using nanopore sequencing. Hortic Res. 2021, 8, 64. [Google Scholar] [CrossRef] [PubMed]
- Mistry, J.; Chuguransky, S.; Williams, L.; Qureshi, M.; Salazar, G.A.; Sonnhammer, E.L.L.; Tosatto, S.C.E.; Paladin, L.; Raj, S.; Richardson, L.J.; et al. Pfam: The protein families database in 2021. Nucleic Acids Res. 2021, 49, D412–D419. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Clements, J.; Eddy, S.R. HMMER web server: Interactive sequence similarity searching. Nucleic Acids Res. 2011, 39, 29–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niu, E.; Jiang, C.; Wang, W.; Zhang, Y.; Zhu, S. Chloroplast genome variation and evolutionary analysis of Olea europaea L. Genes 2020, 11, 879. [Google Scholar] [CrossRef] [PubMed]
- Lumaret, R.; Ouazzani, N. Plant genetics. Ancient wild olives in Mediterranean forests. Nature 2001, 413, 700. [Google Scholar] [CrossRef]
- Diez, C.M.; Trujillo, I.; Martinez-Urdiroz, N.; Barranco, D.; Rallo, L.; Marfil, P.; Gaut, B.S. Olive domestication and diversification in the Mediterranean Basin. New Phytol. 2015, 206, 436–447. [Google Scholar] [CrossRef]
- Julca, I.; Marcet-Houben, M.; Cruz, F.; Gómez-Garrido, J.; Gaut, B.S.; Díez, C.M.; Gut, I.G.; Alioto, T.S.; Vargas, P.; Gabaldón, T. Genomic evidence for recurrent genetic admixture during the domestication of Mediterranean olive trees (Olea europaea L.). BMC Biol. 2020, 18, 148. [Google Scholar] [CrossRef]
- Zohary, D.; Spiegel-Roy, P. Beginnings of fruit growing in the Old World. Science 1975, 187, 319–327. [Google Scholar] [CrossRef]
- Madi, L.; Wang, X.; Kobiler, I.; Lichter, A.; Prusky, D. Stress on avocado fruits regulates Δ9-stearoyl ACP desaturase expression, fatty acid composition, antifungal diene level and resistance to Colletotrichum gloeosporioides attack. Physiol. Mol. Plant Pathol. 2003, 62, 277–283. [Google Scholar] [CrossRef]
- Teixeira, M.C.; Carvalho, I.S.; Brodelius, M. ω-3 fatty acid desaturase genes isolated from purslane (Portulaca oleracea L.): Expression in different tissues and response to cold and wound stress. J. Agric. Food Chem. 2010, 58, 1870–1877. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivated/Wild Olive | Groups | Gene Name | Gene ID | Chr./Scaffold | Protein Length/a.a. | Molecular Weight/Da | Isoelectric Point |
|---|---|---|---|---|---|---|---|
| O. europaea cv. Farga | FAB2/SAD | OeFAB2.1 | OE6A002165 | s09619 | 386 | 43,985.23 | 6.2 |
| OeFAB2.2 | OE6A012975 | s05995 | 396 | 45,549.82 | 6.08 | ||
| OeFAB2.3 | OE6A020845 | s05960 | 366 | 41,893.6 | 5.44 | ||
| OeFAB2.4 | OE6A024078 | s03906 | 94 | 10,932.6 | 6.83 | ||
| OeFAB2.5 | OE6A033129 | s03906 | 159 | 18,196.6 | 5.96 | ||
| OeFAB2.6 | OE6A048475 | s02255 | 390 | 44,535.91 | 6.11 | ||
| OeFAB2.7 | OE6A060676 | s03906 | 208 | 23,921.35 | 8.45 | ||
| OeFAB2.8 | OE6A089828 | s09854 | 399 | 45,584.11 | 6.58 | ||
| OeFAB2.9 | OE6A108617 | s03845 | 291 | 33,386.07 | 5.38 | ||
| OeFAB2.10 | OE6A118450 | s09608 | 283 | 32,301.89 | 5.68 | ||
| ADS | OeADS.1 | OE6A005264 | s08189 | 384 | 43,921.63 | 9.36 | |
| OeADS.2 | OE6A080903 | s00065 | 375 | 43,343.08 | 9.28 | ||
| OeADS.3 | OE6A090227 | s00065 | 384 | 43,987.69 | 9.36 | ||
| OeADS.4 | OE6A120700 | s08189 | 151 | 17,260.67 | 6.64 | ||
| DES | OeDES | OE6A026807 | s03867 | 332 | 38,815.97 | 7.88 | |
| FAD4 | OeFAD4.1 | OE6A064387 | s09642 | 305 | 34,001.84 | 8.19 | |
| OeFAD4.2 | OE6A070143 | s09790 | 305 | 33,954.76 | 8.19 | ||
| OeFAD4.3 | OE6A082617 | s07944 | 237 | 26,292.52 | 8.85 | ||
| SLD | OeSLD.1 | OE6A013963 | s00013 | 447 | 51,607.52 | 8.68 | |
| OeSLD.2 | OE6A023328 | s07941 | 447 | 51,536.29 | 7.93 | ||
| OeSLD.3 | OE6A037200 | s07705 | 447 | 51,568.46 | 8.68 | ||
| OeSLD.4 | OE6A037564 | s00022 | 448 | 51,531.57 | 8.76 | ||
| OeSLD.5 | OE6A087532 | s09955 | 447 | 51,649.44 | 7.63 | ||
| OeSLD.6 | OE6A107382 | s05784 | 447 | 51,823.75 | 8.7 | ||
| OeSLD.7 | OE6A114237 | s01961 | 371 | 43,394.75 | 8.6 | ||
| ω-3 | OeFAD3.1 | OE6A024624 | s02233 | 395 | 45,696.39 | 7.17 | |
| OeFAD3.2 | OE6A086562 | s09646 | 164 | 18,813.28 | 7.77 | ||
| OeFAD3.3 | OE6A109248 | s06024 | 158 | 17,919.2 | 6.19 | ||
| OeFAD3.4 | OE6A113752 | s00046 | 172 | 19,727.44 | 6.7 | ||
| OeFAD7 | OE6A117679 | s04425 | 64 | 7269.06 | 6.08 | ||
| OeFAD8.1 | OE6A074266 | s07757 | 461 | 52,736.2 | 7.79 | ||
| OeFAD8.2 | OE6A075849 | s03965 | 436 | 49,806.14 | 9.18 | ||
| ω-6 | OeFAD2.1 | OE6A011870 | s00121 | 383 | 43,916.5 | 8.73 | |
| OeFAD2.2 | OE6A019200 | s05069 | 381 | 44,216.96 | 8.48 | ||
| OeFAD2.3 | OE6A051290 | s00121 | 383 | 43,907.49 | 8.73 | ||
| OeFAD2.4 | OE6A067271 | s07908 | 381 | 44,167.93 | 8.62 | ||
| OeFAD2.5 | OE6A069627 | s01964 | 381 | 44,182.9 | 8.46 | ||
| OeFAD2.6 | OE6A085290 | s05804 | 380 | 43,729.63 | 8.62 | ||
| OeFAD2.7 | OE6A098403 | s07710 | 383 | 44,064.09 | 9.01 | ||
| OeFAD6 | OE6A116067 | s07955 | 443 | 51,281.56 | 9.21 | ||
| O. europaea var. Sylvestris | FAB2/SAD | OeuFAB2.1 | Oeu009943 | scaffold13793 | 366 | 41,906.65 | 5.53 |
| OeuFAB2.2 | Oeu015714 | scaffold1625 | 208 | 23,865.24 | 7.02 | ||
| OeuFAB2.3 | Oeu024466 | scaffold2105 | 187 | 21,435.4 | 6.53 | ||
| OeuFAB2.4 | Oeu025716 | scaffold2199 | 386 | 43,746.94 | 6.03 | ||
| OeuFAB2.5 | Oeu040504 | scaffold369 | 390 | 44,622.01 | 6.11 | ||
| OeuFAB2.6 | Oeu048444 | scaffold5181 | 106 | 12,242.2 | 8.88 | ||
| OeuFAB2.7 | Oeu050331 | scaffold564 | 407 | 46,693.26 | 6.03 | ||
| ADS | OeuADS | Oeu027810 | scaffold2325 | 314 | 36,183.62 | 8.71 | |
| DES | OeuDES.1 | Oeu027812 | scaffold2325 | 110 | 12,813.52 | 6.96 | |
| OeuDES.2 | Oeu053967 | scaffold655 | 331 | 38,746.7 | 7.31 | ||
| FAD4 | OeuFAD4.1 | Oeu042929 | scaffold41 | 305 | 34,047.89 | 8.24 | |
| OeuFAD4.2 | Oeu062594 | chr7 | 237 | 26,199.36 | 8.62 | ||
| SLD | OeuSLD.1 | Oeu003987 | scaffold1142 | 447 | 51,536.29 | 7.93 | |
| OeuSLD.2 | Oeu009350 | scaffold1349 | 447 | 51,753.64 | 8.33 | ||
| OeuSLD.3 | Oeu036586 | scaffold314 | 450 | 51,825.81 | 8.65 | ||
| OeuSLD.4 | Oeu036587 | scaffold314 | 447 | 51,588.45 | 8.68 | ||
| OeuSLD.5 | Oeu063672 | scaffold949 | 448 | 51,531.57 | 8.76 | ||
| ω-3 | OeuFAD3.1 | Oeu004670 | scaffold1172 | 373 | 43,368.95 | 8.35 | |
| OeuFAD3.2 | Oeu015599 | scaffold162 | 373 | 43,253.83 | 8.61 | ||
| OeuFAD8.1 | Oeu004694 | scaffold1173 | 436 | 49,840.16 | 9.18 | ||
| OeuFAD8.2 | Oeu033588 | scaffold281 | 473 | 54,190.85 | 8.14 | ||
| OeuFAD8.3 | Oeu050958 | scaffold58 | 63 | 7167.97 | 6.47 | ||
| ω-6 | OeuFAD2.1 | Oeu007766 | scaffold1284 | 383 | 44,071.13 | 9.09 | |
| OeuFAD2.2 | Oeu013924 | scaffold1547 | 381 | 44,182.9 | 8.46 | ||
| OeuFAD2.3 | Oeu033739 | scaffold283 | 383 | 43,948.5 | 8.73 | ||
| OeuFAD2.4 | Oeu058547 | scaffold782 | 381 | 44,185.95 | 8.62 | ||
| OeuFAD2.5 | Oeu061755 | scaffold885 | 380 | 43,685.55 | 8.45 |
| Groups | A. thaliana | O. sativa | G. max | B. rapa | B. oleracea | J. regia | O. europaea cv. Farga | O. europaea var. Sylvestris |
|---|---|---|---|---|---|---|---|---|
| FAB2/SAD | 7 | 8 | 5 | 7 | 6 | 9 | 10 | 7 |
| Total soluble FAD | 7 | 8 | 5 | 7 | 6 | 9 | 10 | 7 |
| ADS | 9 | 0 | 2 | 20 | 18 | 1 | 4 | 1 |
| DES | 1 | 1 | 2 | 1 | 1 | 2 | 1 | 2 |
| FAD4 | 1 | 1 | 0 | 4 | 4 | 1 | 3 | 2 |
| SLD | 2 | 1 | 6 | 4 | 5 | 3 | 7 | 5 |
| ω-3 | 3 | 4 | 9 | 6 | 7 | 8 | 7 | 5 |
| ω-6 | 2 | 4 | 17 | 3 | 3 | 6 | 8 | 5 |
| Total membrane-bound FAD | 18 | 11 | 36 | 38 | 38 | 21 | 30 | 20 |
| Total FAD | 25 | 19 | 41 | 45 | 44 | 30 | 40 | 27 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niu, E.; Gao, S.; Hu, W.; Zhang, C.; Liu, D.; Shen, G.; Zhu, S. Genome-Wide Identification and Functional Differentiation of Fatty Acid Desaturase Genes in Olea europaea L. Plants 2022, 11, 1415. https://doi.org/10.3390/plants11111415
Niu E, Gao S, Hu W, Zhang C, Liu D, Shen G, Zhu S. Genome-Wide Identification and Functional Differentiation of Fatty Acid Desaturase Genes in Olea europaea L. Plants. 2022; 11(11):1415. https://doi.org/10.3390/plants11111415
Chicago/Turabian StyleNiu, Erli, Song Gao, Wenjun Hu, Chengcheng Zhang, Daqun Liu, Guoxin Shen, and Shenlong Zhu. 2022. "Genome-Wide Identification and Functional Differentiation of Fatty Acid Desaturase Genes in Olea europaea L." Plants 11, no. 11: 1415. https://doi.org/10.3390/plants11111415