
Oil Palm Breeding in the Modern Era: Challenges and Opportunities


1
Coconut Research Institute, Chinese Academy of Tropical Agricultural Sciences, Wenchang 571339, China
2
Hainan Key Laboratory of Tropical Oil Crops Biology, Wenchang 571339, China
*
Author to whom correspondence should be addressed.
Plants 2022, 11(11), 1395; https://doi.org/10.3390/plants11111395
Submission received: 27 February 2022
/
Revised: 15 May 2022
/
Accepted: 19 May 2022
/
Published: 24 May 2022
(This article belongs to the Special Issue Oil Palm (Elaeis guineensis) Biology, Productivity, Sustainability: From the Organism to the Ecosystem Scale)
Abstract
:Oil palm, a cross-pollinated crop with long generation time, poses a lot of challenges in achieving sustainable oil palm with high yield and quality. The African oil palm (Elaeis guineensis Jacq.) is the most productive and versatile oil-yielding crop in the world, producing more than any other oil-yielding crop. Despite recent challenges, such as stress tolerance, superior oil quality, disease tolerance, and the need for new market niches, there is a growing need to explore and develop new varieties with high yield potential and the genetic diversity required to maintain oil palm yield stability. Breeding is an indispensable part of producing high-quality planting materials to increase oil palm yield. Biotechnological technologies have transformed conventional plant breeding approaches by introducing novel genotypes for breeding. Innovative pre-breeding and breeding approaches, such as identifying candidate genes in wild or land races using genomics tools, can pave the way for genetic improvement in oil palm. In this review, we highlighted the modern breeding tools, including genomics, marker-assisted breeding, genetic engineering, and genome editing techniques in oil palm crops, and we explored certain concerns connected to the techniques and their applications in practical breeding.
1. Introduction
The genus Elaeis consists of two species, namely, E. guineensis Jacq. and Elaeis oleifera. The center of origin and diversity of E. guineensis is found scattered throughout the tropical rainforests of west and central Africa, whereas E. oleifera, also known as American oil palm, is native to south and central America [1]. The most important palm oil importing countries are China, the Netherlands, and Pakistan. Palm oil makes up a significant portion of the world’s edible oils. Palm oil is extracted from the flesh or pulp of the fruit (the outer part) while palm kernel oil is extracted from the soft part of the seed (the inner part). Based on the fruit structure, oil palm is classified into dura (thick shell; less mesocarp), pisifera (shell-less; embryo rarely formed), and the commercially cultivated tenara, the D × P hybrid (thin shell; more mesocarp) with oil content [2,3]. Conventional crosses between dura and pisifera cultivars have typically improved oil palm productivity, particularly oil yield. Tenera, the F1 hybrid resulting from this cross, exhibits desirable traits from both parents [4]. Additionally, there is a high scope for increasing the oil palm yield to a greater extent. After a three-year immature period, the yield per tree increases progressively with age until it peaks around 20 years. As oil palms are typically propagated by the seed, there is a high degree of heterogeneity in the field, and the availability of sustainable production of genetic traits in traditional breeding is limited, resulting in low palm oil yield. Therefore, it is necessary to develop vegetative propagation to produce a larger number of plants from elite palms. Rapid palm growth using tissue culture is the most promising strategy to produce suitable plant materials (offshoots) and achieve high-quality, high yielding, pest-free varieties. As a result, the conception of micropropagation protocols is essential. Significant efforts have been made to improve micropropagation techniques based on either somatic embryogenesis or organogenesis [5]. Plant micropropagation is currently used for plant breeding to circumvent some of the constraints of conventional breeding, and clonal propagation of oil palm by tissue culture is widely used [6]. Plant regeneration has been achieved using callus or cell suspension cultures, and somatic embryogenesis appears to be adequate for oil palm propagation. Another technique to improve the ideal genetic traits of oil palm for the highest quantity and quality of palm oil is through genetic engineering improvements, which can aid in the introduction of the required new traits from other organisms. The application of genetic transformation paves the way for the use of a wide range of beneficial specific genes [7]. Many genetic transformation methods exist, including Agrobacterium-mediated transformation, microinjection, and particle bombardment, among others [8]. Each method has unique requirements that impact productivity. Although Agrobacterium-mediated transformation has several advantages, such as no requirements for special equipment, techniques, or protoplast culture systems, specific gene transfer to target tissues cannot be assured. For species that are not susceptible to infection by Agrobacterium, microinjection by delivery of specific DNA into plant cells is currently the most useful technique [9]. As a result, this strategy is anticipated to enable the transfer of desirable traits, such as quantity and quality of oil yield, to the new oil palm cultivar. However, an efficient oil palm protoplast culture system and specialized procedure are required. In general, each protoplast can form a new cell wall and later initiate cell division, followed by callus formation. This holds the potential for genetic modification of the oil palm through the use of protoplasts. Therefore, oil palm protoplasts are ideal targets for transformation by microinjection or electroporation and protoplast fusion. Recently, protoplast-based biotechnological approaches have been used to improve conventional breeding of various species, and oil palms can be improved through somatic hybridization and genetic transformation. Genome editing using artificial nucleases appears to be more efficient than conventional plant breeding methods or typical genetic engineering (transgenic or genetically engineered) methods [10]. Genome editing using artificial nucleases appears to be more efficient than conventional plant breeding methods or typical genetic engineering (transgenic or genetically engineered) methods [10]. As the potential and prospects of oil palm have been gradually recognized by all sectors of the population, including farmers, common people (consumers), and entrepreneurs, research and development on various aspects of oil palm are becoming more and more important in the country. This review provides new insights into oil palm through biotechnological tools that enable us to achieve sustainable oil palm production, as well as the ultimate success of its application in breeding improvement.
2. Genetic Diversity of Oil Palm Germplasm
Germplasm collections are genetic treasures that serve as the basis for all plant-related initiatives. Despite the importance of germplasm in agricultural development programs, germplasm collection and conservation are significant in many underdeveloped nations. Crossbreeds between pisifera and dura are used to produce commercial (hybrid) tenera planting materials. For the production of tenera planting materials, the world’s oil palm seed industry has relied on a limited genetic basis. Intensive breeding and selection of Deli dura and Avros pisifera populations from Deli over several generations has resulted in the production of hybrid seeds that are genetically related and/or highly similar to several planting materials used worldwide [11]. The introduction of improved cultivars, land rivalry as a result of population growth, and the adoption of farming practices that are accelerating the destruction of genetic resources of oil palm diversity lead to the need to prevent the rapid loss of oil palm genetic resources through collection, conservation, and utilization for crop development being essential to the survival of the oil palm industry in the country. From the 1950s to the present, the primary missions of many oil palm research organizations have been to prospect, characterize, conserve, and evaluate the natural diversity of oil palm germplasm [12]. According to reports from 29 institutes, 21,103 oil palm accessions have been collected globally. Since the 1970s, the Malaysian Palm Oil Board (MPOB) has gathered 1467 oil palm accessions from wild populations, and these accessions have been conserved [13]. MPOB possesses the world’s largest collection of oil palm germplasm [14]. Therefore, oil palm germplasm accessions were investigated and discovered to contain high levels of genetic variation.
2.1. Oil Palm Germplasm Characterization
Oil palm germplasm characterization is the determination of the performance of materials in a collection. It refers to agronomic, morphological, and molecular fingerprints of the material in the gene bank and leads to the identification of genes encoding resistance to biotic and abiotic stress, precocity, dwarfism, productivity, and good fruit quality traits. Morphological, molecular, and biochemical approaches have been used to study genetic variation between individuals and populations. Oil palm germplasm has frequently used all three techniques, but morphological methods require extensive observation of plants in the field for real-time verification of plant performance over a long period of time [15].
2.2. Morphological Characterization of Oil Palm Germplasm
Morphological characterization will be used to examine genetic variation in oil palm by using vegetative, yield, fruit, and bunch traits. It is also used to assess the agronomic potential of oil palm germplasm and detect duplication. Commercially important characteristics, such as high oil output, superior bunch quality component, low height increment, large kernel, and long stalk, are identified. Agronomic traits, such as plant height, height increment, leaf area, leaf area index, frond dry weight, rachis length, total fronds, total leaf area, number of bunches, bunch weight, average bunch weight, fresh fruit bunch yield, single fruit weight, percent fruit to bunch, percent mesocarp, percent shell, percent kernel, are some of the parameters that are frequently evaluated under morphological techniques. The preponderance of oil palm institutions has employed these physical features to determine the genetic diversity of their oil palm collections [16]. These could be exploited to develop improved dura and pisifera fruit types that will serve as parental palm varieties for producing commercial (tenera) oil palm varieties.
2.3. Molecular Characterization of Oil Palm Germplasm
Despite the fact that morphological characterization is promising, it has a number of flaws. Environmental variables may play a role in the differences, which are also time consuming and age dependent. Molecular markers, such as simple sequence repeats (SSR), random amplified polymorphic DNA (RAPD), and amplified fragment length polymorphism (AFLP), have been used to study genetic variation in oil palm [17]. Considerable genetic diversity within the MPOB germplasm collections has been reported using EST-SSR and isozyme analysis [18]. Okoye et al. [11] found high scores for heterozygosity and genetic diversity of 0.69 and 0.70, respectively, while using 10 microsatellite markers to assess genetic variety among palm parental lines. Using molecular markers to evaluate oil palm accessions addresses variance at the DNA level. DNA extraction is required prior to using the molecular marker approach. Molecular approaches are more expensive, particularly when it comes to analyzing large datasets. The study objectives, cost, and intrinsic qualities of the marker are the decisive factors in the selection of a marker approach.
2.4. Characterization of Oil Palm Germplasm Using Biochemical Techniques
Biochemicals, such as fatty acid composition, iodine value, and carotene content, are used to study genetic variability in oil palm germplasm. Noh et al. [19] studied accessions of Angolan oil palm with low lipase levels but high amounts of carotenes, fatty acid compositions, vitamin E, iodine, and oleic acid. Their findings revealed a high level of genetic variation among the accessions. The levels of biochemical markers, such as palmitic acid, were determined using a gas chromatography system and a spectrophotometer.
3. Role of Conventional Breeding in Oil Palm
Traditional plant breeding based on phenotypic selection has historically been used for effective crop improvement and development. Several classical breeding methods have been developed in the last century and are now used all over the world. To respond to global problems, oil palm breeders are constantly searching for new and improved palm cultivars. There are a number of tools and technologies, which, combined with our knowledge of oil palm and elite germplasm base, create two pathways for product development (Figure 1). Both pathways are deeply grounded in genetic approaches, in which we assay for a trait and then discover the underlying genes. The delineation of these genes using mapping/molecular markers identifies genes that are necessary and sufficient for a trait. Today, increasingly, both paths must come together to complete the package before a new product is developed. Conventional breeding refers to the acquisition of a hybrid new variety of oil palm from the crossing of two well-known varieties. The initial cross is followed by the selection of progeny with the wanted characteristics of resistance and for the described scope with the dwarfing phenotype. Crosses between E. guineensis and E. oleifera produce fertile hybrids, indicating the similarity between the two species. Male and female inflorescences are produced separately on the same palm, showing that the species is monoecious. This category of strategies refers to methods of crossing compatible plants, i.e., plants that would naturally interbreed without the help of the breeder. These techniques have been used since the dawn of agriculture. Two fundamental processes belong to this group: cross-pollination (breeding of a plant with another plant) and self-pollination (breeding of the plant with itself) [20]. Oil palm is a cross-pollinated plantation plant that grows gradually. Both Elaeis species are diploid, with n = 16 [21]. The 2C nuclear DNA content of E. guineensis was 3.76–4.32 pg [22], slightly lower than that of E. oleifera with 4.43 pg [22]. Because of their physical resemblance, oil palm chromosomes are difficult to differentiate cytogenetically. Most published research works identifying E. guineensis chromosomes were based on chromosomal length [23]. The whole-genome sequence of E. guineensis has assigned the individual E. guineensis chromosomes according to the size of sequence scaffolds, which correspond to the linkage group in the selected oil palm mapping population [24]. The reported length of the assembled oil palm genome was 1.535 Gb, representing some 85% of the total genome size, although only 43% of this was assembled and assigned to chromosomes. When compared to other oil crops, the E. guineensis genome is greater than soybean [25] and rapeseed [26] but smaller than corn and coconut [27,28].
In general, oil palm breeding programs are utilized to promote both dura (D) and pisifera (P) populations for the production of commercial D × P seeds. Commercial oil palm varieties are typically derived from progeny tests of crosses between selected parents. The pisifera used in hybridization are usually produced by self or crosses (tenera × tenera (T × T)), tenera × pisifera (T × P) hybridization, or pisifera × pisifera (P × P) self or crosses. On the other hand, the dura is selected from the dura × dura (D × D) autologous or cross. In this scenario, the selected dura palm trees offspring can continue to the next cycle [17]. Breeders are focused on developing cultivars with high fresh fruit (FFB) yields and higher mesocarp content to increase palm oil productivity. An optimal GS method should provide the highest possible accuracy, limit overfitting of the training set, and be based on marker QTL LD rather than relatedness. In addition, such approaches must be easy to implement, consistent across a variety of functions and datasets, and computationally efficient. Perennial plants can benefit more from GS than annual plants [29]. Oil palm breeding as we know it uses the selection process to create new varieties of plants with the intention of improving the genetic potential of oil palm species. Commercial variations are developed around the world by pollinating Deli dura mother palms with African pisifera pollen, and a recurrent selection strategy appears to be particularly appropriate for oil palms. Conventional breeding approaches have been highly effective in modifying the FA profiles of edible oil crops to develop commercial oils due to the unique and exotic nature of many industrially relevant fatty acids (FA). Oil palm cultivation takes about 12 years for the first generation and 40 years for the next eight generations. This is accomplished by using a vast planting area due to oil palm’s open pollination behavior. As a result, the development of novel oil palm traits or variants via conventional breeding is significantly delayed. Unlike the common practice with annual plants, it is impossible to produce homozygous parental families because of the length of oil palm reproduction cycle and strong inbreeding depression. Several traits, such as dwarf palms and plants high in vitamin E and oleic acid, have been developed. On the other hand, the oil palm has a serious problem with incomplete dominance inheritance. For example, the tenera fruits displayed all three fruit forms, dura:tenera:pisifera, in a 1:2:1 ratio, indicating the incomplete dominance of dura over pisifera. That is, only 50% of the fruits retained tenera properties. This problem makes oil palm propagation by seed germination unsatisfactory.
4. Importance of Tissue Culture in Oil Palm
Oil palm cannot be propagated by vegetative tissue culture, as it has a single growing apex with no base branches. Oil palm as a monocot grows from a primary shoot meristem and has no apparent morphological basis for vegetative reproduction. Therefore, oil palm tissue culture is unique, undergoing callus and embryogenesis processes and rigorously tested in the 1960s and 1970s. Until recently, no substantial attempt was made to develop tissue culture programs for the propagation of elite palm. Some attempts have been made with seedling or embryo conditions described in regeneration protocols [6]. Therefore, oil palm tissue culture is entirely dependent on somatic embryogenesis [30]. As a result, in vitro regeneration of mature elite palms or tissue culture is the default method to proliferate an elite palm, and hence, the most essential component in oil palm development. The procedure of oil palm tissue culture is separated into many stages: (i) Sampling of explant from selected ortet; (ii) Callus initiation in explants; (iii) Embryogenesis; (iv) Shoot and root regeneration; (v) Transplanting of plants from ex vitro condition. However, the long process of solid culture medium (52–55 months) resulted in a significant proportion of oil palm abnormalities [31], which were mostly attributed to somaclonal variation. In order to improve the frequency of somaclonal variation, embryogenic structures are commonly disseminated to enhance SE efficiency [32]. Under certain circumstances, embryogenic cells and non-embryogenic cells can only be distinguished by morphological traits and a number of potential genes with differential expression between embryogenic callus. These include a putative Aux/IAA gene and Eg707, an unidentified protein implicated in abscisic acid production [33]. Although highly efficient transformation systems have been achieved in oil palm species, most transgenic oil palms are still generated using transformation protocols based on tissue culture methods, which are genotype independent, time consuming, and laborious (Figure 2). Major improvements have been made in many tissue cultures, notably in oil palm suspension culture, where the process has been shortened by at least 35 months. Therefore, oil palm suspension culture has become a valuable starting material for oil palm genetic engineering to accelerate the transformation process [34].
5. Genetic Engineering of Oil Palm
Numerous techniques for transferring genes into plant cells have been developed, and ongoing attempts are made to improve efficiency. Genetic engineering is perhaps the most effective way to address the limitations of conventional breeding, which would be difficult or impossible to achieve with traditional methods. The ultimate focus of these efforts is the development of a transgenic oil palm with a high oleic acid content [35]. MPOB pioneered the use of genetic engineering to grow palm trees with a high oleate content for the industrial commodity and liquid oil industries. When oleic acid content is >65%, the estimated value of high oleate palms is USD 1500/ha/year [36]. Oil quality modification, such as stearic acid increase, value-added oil, such as palmitoleic acid and ricinoleic acid synthesis, and novel products, such as biodegradable polymers, have been studied. The development of a reliable transformation and regeneration mechanism is of crucial importance for genetic engineering. Currently, biolistic and Agrobacterium-mediated transformations are commonly used to introduce useful genes into oil palm (Table 1). The parameters influencing the biolistics for oil palm transformation, such as the appropriate selectable marker [37], promoter selection [38], and ideal physical and biological factors [39], have been determined. This bombarded several hundred embryogenic calli with numerous genes, including those involved in the fatty acid biosynthetic pathway, to enhance oleic and stearic acid [40] and polyhydroxyvalerate (PHBV) genes to produce biodegradable polymers [41].
Agrobacterium-mediated transformations have also been used to improve the oil palm through genetic engineering. However, oil palm is not a natural host for Agrobacterium-like dicots. Nevertheless, several studies have been conducted to improve this method, including the use of immature embryos (IE) [42] and immature zygotic embryos (IZE) [43] as target tissues, as well as the parameters that affect Agrobacterium-mediated transformation of embryogenic oil palm calli [41]. The Bacillus thuringiensis insecticide protein gene (Bt) [44], the cowpea trypsin inhibitor gene (CpTI) [45], which responds to insect pest, and the PHB gene [46] have all been transmitted by Agrobacterium-mediated oil palm transformation. The successful regeneration of transgenic oil palm relied on selectable markers. Genes encoding hpt (hygromycin phosphotransferase), pat or bar (Phosphinothricin acetyltransferase), nptll (neomycinphosphotransferase), and EPSPs were the most often adopted selective markers for crop transformation [47]. These marker genes are implemented efficiently for selection of Agrobacterium-transformed cells when controlled by constitutive promoters, such as the 35S promoter from cauliflower mosic virus and ubiquitin promoters [48]. Using the A. tumefaciens strain LBA4404, the plasmid pBIDOG with the DOGR1 gene under the control of the 35S promoter was converted into oil palm embryogenic calli [49].
A longer selection process for plants developed from callus formation and somatic embryogenesis tends to result in chimeric transgenic plants more frequently. It has been suggested that both methods still require optimization of DNA delivery and selection of transformants to overcome problems associated with escapes. To overcome problems related to biolistics and Agrobacterium-mediated transformation, an alternative transformation method is urgently needed, and transformation of oil palm protoplasts using DNA microinjection is a promising approach. Masani et al. [39] developed unique transformation methods based on polyethylene glycol-mediated transfection and DNA microinjection to show that protoplasts are good targets for genetic manipulation of oil palm. Intense electrical pulses cause transient porosity to develop in the cell membrane, allowing molecules, including DNA, RNA, proteins, antibiotics, and pigments, to be transported into the cell [50]. Although protoplasts were the most common target for electroporation in plant research, multiple investigations showed that the technology could also be utilized to transfer genes directly into plant tissues. Some researchers have reported that utilizing linear plasmid DNA rather than circularized plasmid DNA, as well as infusing spermidine into the incubation buffers, has an effect on transformation efficiency. The incorporation of new traits into established species can reduce the time required for genetic improvement in oil palm. The appearance of a blue spot in the plant tissue as a result of GUS activity toward the X-Gluc substrate may indicate that the transformation technique was effective. The gene has also been used to express transitory genes in various types of oil palm explants, including somatic embryos [51]. Similarly, Chris Darmawan [52] reported that embryogenic oil palm calli were transferred using a vector, plasmid pCAMBIA 1303 driven by the 35S promoter carrying the GUS gene, and that transformation was successful. This system relies heavily on effective plant transformation and regeneration techniques. The main advantage of the approach is that the tissues are kept in the same physiological state after electroporation as before the transformation.

{kind=link}
{kind=link}
Table 1.
Genetic transformation of oil palm through Agrobacterium-mediated transformation and direct DNA transfer methods.
Table 1.
Genetic transformation of oil palm through Agrobacterium-mediated transformation and direct DNA transfer methods.
| Explants |
Transformation/
Strain | Vector | Selection Marker | Promoter Used | Reporter Gene |
Transgene/
Expression | Studied Parameters | References |
|---|---|---|---|---|---|---|---|---|
| Young Leaves | LBA4404 | pUBA | bar | Ubi1 | Gus | Southern Bolt | Study glufosinate-ammonium- resistant transgenic oil palm | [41] |
| Embryonic Calli | LBA4404 | pBIDOG | DOGR1, | CaMV35S | Gus | PCR/ Southern Bolt | Introduction of new selection agent | [49] |
| Immature Embryo | LBA4404 | pCAMBIA 1310 | nptII | CaMV35S | Gus | Gus Assay | In vitro culture of IE for direct plant regeneration | [42] |
| Protoplast, Basidiospore, mycelium | LBA4404 | pCAMBIA 1300 | hyg | gpd fungal promoter | Gus/gfp | PCR | Understanding pathogenicity factor associated with G.boninense | [53] |
| Calli Clump | Electroporation | pCAMBIA 1303 | hptII | CaMV35S | GusA/ mgfp5 | PCR | An efficient electroporation-mediated transformation method for oil palm calli | [52] |
| Embryonic Calli | Biolistic | pMI11, pMI11G, pMI3, pMI3G | pmi gene | Ubi1, CaMV35S | GusA | PCR/ Southern Bolt | To produce transgenic oil palm by using mannose as a selectable agent | [38] |
| Immature Embryo | Biolistic | pBI121 | npt II | CaMV35S | Gus | Gus Assay | In vitro culture of IE for indirect plant regeneration | [42] |
| Embryonic Calli | Biolistic | pBINPLUS | hpt | 35S (2XCaMV35S) | GFP | PCR/ Southern Bolt | Increase the level of transgene expression and transformation efficiencies in oil palm | [54] |
| Immature Embryo | Biolistic | pPSP’AP-VF6 | HYG | CaMV35S, MSP | - | PCR | Successful integration of antiPATE driven by MSP in American oil palm plantlets | [43] |
6. Association Mapping for QTL Identification
In the MAS approach, molecular markers are used for indirect selection of challenging qualities, such as traits that are not phenotypically visible at the seedling stage [55]. Molecular markers are DNA strings that are distributed across the plant genome and may be tracked as they segregate through generations. Because known genes of interest are linked together, the presence of the marker ensures the presence of the gene of interest and allows the breeder to track its segregation throughout breeding. Because breeders can recognize the markers in seedlings and do not need to acquire adult plants for selection, MAS speeds up the process of conventional plant breeding. Furthermore, MAS facilitates the enhancement of traits that cannot be easily determined using conventional methods [55]. Depending on the markers, these procedures range from gel electrophoresis, PCR, through contemporary high-throughput genotyping techniques. MAS has become a standard phase in the breeding of most crops and provides several benefits, including the increasing knowledge about molecular markers, genes of interest, and their localization. Biological traits are often governed by either a large number of genes with minimal effects (polygenic inheritance), termed as quantitative traits, or by single genes with significant effects (monogenic inheritance), referred to as qualitative traits. Quantitative traits in plants have been targets of crop improvement for yield, quality, and pest resistance. Quantitative traits are associated with QTL loci, containing genes on a chromosome. Genes that regulate quantitative traits interact with one another, and the expression of these traits is often influenced by environmental conditions. To study polygenic characteristics, quantitative trait locus analysis is utilized. Multiple potential parameters, including fruit weight, petiole cross-section, axial length, and shell:fruit, mesocarp:fruit, and kernel:fruit ratios have been empirically proved. MPOB also executed QTL research on oil palm oil quality, discovering 11 QTLs in four distinct linkage groups with iodine value C14:0, C16:0, C16:1, C18:0, C18:1, and C18:2, using a frame diagram comprising AFLP, RFLP, and SSR markers [56]. Using an interspecies pseudo-backcross of E. guineensis and E. oleifera revealed 19 QTLs pertaining to palm oil fatty acid content [2]. MPOB is Malaysia’s leading oil palm research and development center. MPOB has carried out extensive genetic diversity studies that can quantify the genetic distance between different breeding materials, assisting in the identification of new elite material suited for introgression into breeding programs. Using progenies derived from a self-pollinated E. guineensis palm (Palm T128), they constructed a linkage map and identified an RFLP marker for Sh gene [56]. Four distinct SNP markers were found to be mapped on either side of the Sh gene, with the closest marker (SNPM00310) at a distance of 2.2 cM. Gan et al. [57] used high-density SNP and DArT markers, with SSR markers as anchor loci, to construct genetic linkage maps of two closely related oil palm populations, which enable the discovery of markers strongly associated with the ‘shell thickness’ (Sh) gene. When the population is biased by the natural population and other important criteria (such as high-resolution QTL mapping, more alleles, less time, and a larger reference population), association mapping provides numerous advantages. However, association mapping in oil palm is uncommon. Using 200K SNP to perform GWAS on the oil-to-dry mesocarp content of 2045 genotyped oil palms, 80 loci were determined to be substantially correlated with oil-to-dry mesocarp yield (P104), and three important signals were determined [58]. They described the most comprehensive use of high-density SNP genotyping and GWAS methodologies to identify SNP variations correlated with differences in important yield parameters in oil-to-dry mesocarp and further confirmed their involvement in independent crosses.
7. Functional Markers and Their Application
Molecular markers have been found to be an effective technique for assessing genetic diversity and elucidating genetic relationships among species. Molecular markers, such as RAPD, AFLP, and SSR, have been utilized to simulate genetic diversity coupling and association mapping, as well as the construction of linkage maps. Two important monogenic hereditary traits of oil palm are the gene for fruit color (Vir) and the gene for shell thickness (Sh). Two RFLP markers associated with the fruit color gene were identified in the MPOB linkage map: MET16 (3 cM) and KT3 (4 cM) [59]. It was discovered that the markers are able to distinguish not only the Nigrescens and Virescens fruits but also the homozygous and heterozygous variants of the Vir fruit. Due to the lack of carotenoids in the exocarp, Vir fruits are green when immature and bright orange when ripe. Many molecular approaches have been employed to study the shell thickness gene (Sh) and/or marker(s). Mayes et al. [60] developed an oil palm RFLP linkage genetic map using a population derived from a self-pollinated tenera palm, which segregated by shell thickness, allowing the discovery and mapping of an RFLP marker (pOPgSP1282), which is distantly connected to Sh by 9.8 cM. This marker is quite far from Sh to be used to identify crop species, as the probability of recombination between the marker allele and the gene is still high. RAPD markers have the potential to be utilized to genotype both parents and hybrids [61]. Singh et al. [62] identified two mutations in the MADS box transcription factor of the SHELL gene, which is homologous to the Arabidopsis thaliana gene, SEEDSTICK (STK), which controls ovule identity through homozygosity mapping. They also detected an SNP in the 28 or 30 codon that prevents shell (Sh) from binding to DNA normally, resulting in a shell-less phenotype. RAPD markers have been used to evaluate the genetic diversity of natural palm oil species, such as E.oleifera species [63]. In another study, RAPD work by Moretzsohn et al. [64] on linkage mapping of the shell thickness locus also showed two different markers, R11-1282 and T19-1046, which were 17.5 cM and 23.9 cM, respectively, on either side of the sh+ locus. Although the two markers were even further removed from the Sh gene, the authors claimed that using a flanking marker-based assay would allow tenera and pisifera palms resulting from a D × P cross with an error rate of only 4% (0.175 × 0.239 = 0.042). Therefore, with these two markers, a more accurate and faster identification of the fruit shape is possible. Dou et al. [65] recently performed genetic variation analyses on a panel of twenty oil palm accessions to find single-nucleotide polymorphisms (SNPs) associated with vitamin E content. The BLASTP analysis revealed 37 similar genes in oil palm corresponding to the vitamin E biosynthetic pathway, with lengths ranging from 426 to 25,717 bp (average 7089 bp). Within the Arecaceae family, the utility of transcribed libraries and cDNA tag archives has resulted in the development of EST-SSR in oil palm [66]. Other investigations on Cocos nucifera and P. dactylifera found similar findings [67]. Although most of these methods are unfeasible due to the high cost of implementation, they have become obsolete as the utility of simple sequence repeats/microsatellites has evolved. Billotte et al. [68] discovered a high-density microsatellite-based backtracking map of oil palms. It is the first linkage map with 16 independent linkage groups corresponding to the 16 homologous chromosome pairs of oil palm. Using 944 SSR and AFLP markers, this integrated map spanned 1743 cM of the plant genome. It was discovered that E-AGG/M-CAA132, an AFLP marker, maps at 4.7 cM from the Sh locus and is positioned near the terminus of LG4. Recently, Babu et al. [69] discovered a cleaved amplified polymorphic site (CAPS) marker for oil palm fruit type differentiation. The CAPS identifier will enable the selection and timely delivery of attractive high-yielding variants. For oil palm, Bai et al. [70] identified two potential QTL that are related to foliar surface, and this trait is strongly related to oil yields. This would allow breeders to select high-oil-yielding trees indirectly through MAS by identifying trees with a large leaf area and reduce the laborious and destructive phenotyping required to measure oil efficiency.
Genomic Selection of Oil Palm
For quantitative traits, the efficiency of QTL-based marker-assisted selection (MAS) is limited because it overestimates the effect of strong QTLs and fails to exploit weak QTLs [71]. To overcome these issues, genomic selection was developed. Oil palm is a diploid, monoecious, allogamous plant with a high GS potential. Reciprocal recurrent selection (RRS) is the most used method for improving oil palm genetics [72]. The RRS is based on two populations: the Deli and Group B (African origin). In each of the two populations, full-sib families were sampled, and progenies were examined in Deli × Group B crosses. Genomic selection has the potential to rapidly evolve heterotic pools in plants where they are not well established, such as oil palm. When selecting single crosses, genomic prediction could reduce the need for test crosses and field evaluations [73]. RRGS programs have been initiated in oil palm, which has a long generation interval and high phenotyping costs [74]. The genomic feature modeling approaches were designed to improve the prediction of complex traits, provided the groups of selected markers were enriched for causal variants. The potential of genomic models to forecast the genetic value of unevaluated selection candidates was exhibited using SNPs and without optimizing the training and validation populations, with prediction accuracies ranging from 0.14 to 0.73 for various yield components [74]. The GS model predicted the performance of unevaluated hybrid crossings better than a control model utilizing pedigree data instead of markers for five yield components (FFB, O/M, BN, BW, and M/F) [74]. This emphasized GS’s ability to capture genetic variations within full-sib families, as well as genetic differences across families, allowing for the selection of the best individuals from the best families, as is now performed with phenotypic breeding schemes.
8. Genome Editing Technology
The utilization of the CRISPR/Cas9 system in genome editing has found wide application in various fields, such as medicine and agriculture [75]. The versatility of this system has been applied from knockout or edits of a gene, regulating gene expression until single nucleotide polymorphisms arise [76]. In plant genome editing, two currently used delivery methods of the CRISPR/Cas9 system are particle bombardment and Agrobacterium transformation. The CRISPR/Cas9 module is simpler than ZFNs and TALENs and is regarded as a breakthrough technology for genome editing. The module is based on Cas9 nuclease and a modified RNA single guide (sgRNA) for editing target nucleotide sequences [75]. This could be efficiently exploited in the oil palm to modify desirable genes by targeting specific DNA sequences with tailored nucleases containing sequence-specific DNA-binding domains. Recently, a novel technique based on the CRISPR-Cas9 system, termed ‘base editing’, has been developed. Most recently, Yarra et al. [77] proposed the notion of using base editing techniques in oil palm. It is HDR independent and edits single DNA bases at precise places in the genome without causing DSBs or adding a foreign DNA template. Although base editing has been effective in improving rice and wheat yields, its application to other crops is still in its early stages. In oil palm, the CRISPR/Cas9 transgenes can be delivered via young leaves, and stably transformed plants can be regenerated via tissue culture. Bohari et al. used rice as a model system in order to develop a genome editing system for oil palm. The genomic sequences of rice and oil palm were discovered to have considerable similarities [78]. This might be utilized efficiently in oil palm to modify desirable genes by targeting specific DNA sequences with tailored nucleases containing sequence-specific DNA-binding domains. Transcription activator-like effector nucleases were utilized to downregulate two fatty acid desaturase 2 genes (FAD2-1A and FAD2-1B) in soybean, resulting in an increase in oleic acid content due to a decrease in linoleic acid content [79]. Even though it is a great approach with a multitude of benefits, it could be used in oil palm to more effectively regulate specific genes or to replace an undesirable gene. With the advent of oil palm genome sequencing, this has now become feasible.
9. Conclusions
Unlike other oil crops, palm oil also has the most sustainability certification schemes. In 2004, a non-profit organization called the Roundtable on Sustainable Palm Oil (RSPO) was formed. The RSPO has become a leading certifier, developing a set of environmental and social criteria for the production of certified sustainable palm oil (CSPO). In other words, the level of rigor around the sustainability of palm oil is unprecedented. Because of this, the expansion of cultivation through deforestation is now banned in many countries, and fertile land is being reduced by urban development. Therefore, many research organizations had initiated the efforts to improve oil yield on the existing land bank to meet increasing demand. On the other hand, selective breeding for high-yielding, stress-tolerant materials is also equally essential in the long run; however, the breeding cycle of perennial palm oil is about 12 years per generation. Therefore, improving oil palm yield through conventional breeding is time consuming, while increasing production through simply expanding acreage is unsustainable. The application of the diagnostic and analytical power in modern biotechnology would be a great advantage in oil palm breeding programs. There is a heavy commitment in terms of labor for traits maintenance and recording. All these factors result in high costs of breeding programs. It is clear that biotechnology offers unprecedented opportunities to improve plant breeding. Biotechnological application offers researchers new insights and tools that improve their efficiency and effectiveness. This stabilizes the broad phenotypic variability of the heterozygous hybrids, resulting in recognized palm cultivars with consistent traits. The use of molecular tools and transgenic crops will allow us to meet our food needs in a sustainable manner with limited availability of resources. Therefore, the development of biotechnological tools to maximize the progression of genomic resources in oil palm is essential for the continuous improvement in oil yield and the overall quality of oil produced, as well as the production of market-responsive varieties, which are essential for long-term agricultural sustainability.
Author Contributions
Conceptualization: H.C.; writing—original draft preparation: J.J.J.M., R.Y., H.C., and L.W.; supervision: H.C.; funding acquisition: H.C. and L.W.; project administration: H.C. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Central public-interest scientific institution basal research fund for the Chinese Academy of Tropical Agricultural Sciences (No.1630152021008) and Protection of species resource (crop) of Ministry of Agriculture and Rural Affairs (No.2021NWB048), Protection of species and variety resources on tropical palm of Ministry of Agriculture and Rural Affairs (No. 18200039).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Montúfar, R.; Louise, C.; Tranbarger, T. Elaeis oleifera (Kunth) Cortés: A neglected palm from the Ecuadorian Amazon. Rev. Ecuat. De Med. Y Cienc. Biol. 2018, 39, 11–18. [Google Scholar] [CrossRef]
- Montoya, C.; Lopes, R.; Flori, A.; Cros, D.; Cuellar, T.; Summo, M.; Espeout, S.; Rivallan, R.; Risterucci, A.M.; Bittencourt, D.; et al. Quantitative trait loci (QTLs) analysis of palm oil fatty acid composition in an interspecific pseudo-backcross from Elaeis oleifera (H.B.K.) Cortés and oil palm (Elaeis guineensis Jacq.). Tree Genet. Genomes 2013, 9, 1207–1225. [Google Scholar] [CrossRef] [Green Version]
- Astorkia, M.; Hernández, M.; Bocs, S.; Ponce, K.; León, O.; Morales, S.; Quezada, N.; Orellana, F.; Wendra, F.; Sembiring, Z.; et al. Detection of significant SNP associated with production and oil quality traits in interspecific oil palm hybrids using RARSeq. Plant Sci. 2020, 291, 110366. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Sun, Y.; Qu, J.; Syah, R.; Lim, C.H.; Alfiko, Y.; Rahman, N.E.B.; Suwanto, A.; Yue, G.; Wong, L.; et al. Transcriptome and functional analysis reveals hybrid vigor for oil biosynthesis in oil palm. Sci. Rep. 2017, 7, 439. [Google Scholar] [CrossRef] [Green Version]
- Tisserat, B. Clonal propagation of palms. In Plant Tissue Culture Manual: Supplement 7; Lindsey, K., Ed.; Springer: Dordrecht, The Netherlands, 1997; pp. 473–486. [Google Scholar]
- Rajesh, M.K.; Radha, E.; Karun, A.; Parthasarathy, V.A. Plant regeneration from embryo-derived callus of oil palm—The effect of exogenous polyamines. Plant Cell Tissue Organ Cult. 2003, 75, 41–47. [Google Scholar] [CrossRef]
- Borisjuk, N.; Kishchenko, O.; Eliby, S.; Schramm, C.; Anderson, P.; Jatayev, S.; Kurishbayev, A.; Shavrukov, Y. Genetic Modification for Wheat Improvement: From Transgenesis to Genome Editing. BioMed Res. Int. 2019, 2019, 6216304. [Google Scholar] [CrossRef] [PubMed]
- Ortiz-Matamoros, M.F.; Villanueva, M.A.; Islas-Flores, T. Genetic transformation of cell-walled plant and algae cells: Delivering DNA through the cell wall. Brief. Funct. Genom. 2018, 17, 26–33. [Google Scholar] [CrossRef]
- Yadava, P.; Abhishek, A.; Singh, R.; Singh, I.; Kaul, T.; Pattanayak, A.; Agrawal, P.K. Advances in Maize Transformation Technologies and Development of Transgenic Maize. Front. Plant Sci. 2017, 7, 1949. [Google Scholar] [CrossRef] [Green Version]
- Khatodia, S.; Bhatotia, K.; Passricha, N.; Khurana, S.M.P.; Tuteja, N. The CRISPR/Cas Genome-Editing Tool: Application in Improvement of Crops. Front. Plant Sci. 2016, 7, 506. [Google Scholar] [CrossRef] [Green Version]
- Okoye, M.; Uguru, M.; Bakoumé, C.; Singh, R.; Okwuagwu, C. Assessment of Genetic Diversity of NIFOR Oil Palm Main Breeding Parent Genotypes Using Microsatellite Markers. Am. J. Plant Sci. 2016, 7, 218–237. [Google Scholar] [CrossRef] [Green Version]
- Barcelos, E.; Rios, S.D.A.; Cunha, R.N.V.; Lopes, R.; Motoike, S.Y.; Babiychuk, E.; Skirycz, A.; Kushnir, S. Oil palm natural diversity and the potential for yield improvement. Front. Plant Sci. 2015, 6, 190. [Google Scholar] [CrossRef] [PubMed]
- Rajanaidu, N. Institiut Penyelidikan Minyak Kelapa Sawit M. PORIM Oil Palm Genebank: Collection, Evaluation, Utilization and Conservation of Oil Palm Genetic Resources; Palm Oil Research Institute of Malaysia: Kuala Lumpur, Malaysia, 1994. [Google Scholar]
- Okwuagwu, C.O.; Ataga, C.D.; Okoye, M.; Okolo, E.C. Germplasm Collection of Highland Palms of Afikpo in Eastern Nigeria. Bayero J. Pure Appl. Sci. 2011, 4, 112–114. [Google Scholar] [CrossRef]
- Bakoumé, C.; Wickneswari, R.; Siju, S.; Rajanaidu, N.; Kushairi, A.; Billotte, N. Genetic diversity of the world’s largest oil palm (Elaeis guineensis Jacq.) field genebank accessions using microsatellite markers. Genet. Resour. Crop Evol. 2015, 62, 349–360. [Google Scholar] [CrossRef]
- Okwuagwu, C.O.; Okoye, M.; Okolo, E.C.; Ataga, C.D.; Uguru, M. Genetic variability of fresh fruit bunch yield in Deli/dura×tenera breeding populations of oil palm (Elaeis guineensis Jacq.) in Nigeria. J. Trop. Agric. 2008, 46, 40–45. [Google Scholar]
- Rajanaidu, N.; Kushairi, A.; Rafii, M.; Mohd Din, A.; Maizura, I.; Jalani, B.S. Oil palm breeding and genetic resources. In Advances in Oil Palm Research; Malaysian Palm Oil Board: Kuala Lumpur, Malaysia, 2000; pp. 171–237. [Google Scholar]
- Hayati, A.; Wickneswari, R.; Ithnin, M.; Rajanaidu, N. Genetic diversity of oil palm (Elaeis guineensis Jacq.) germplasm collections from Africa: Implications for improvement and conservation of genetic resources. TAG Theor. Appl. Genet. Theor. Und Angew. Genet. 2004, 108, 1274–1284. [Google Scholar] [CrossRef]
- Noh, A.; Rafii, M.Y.; Saleh, G.; Kushairi, A.; Latif, M.A. Genetic Performance and General Combining Ability of Oil Palm Deli dura x AVROS pisifera Tested on Inland Soils. Sci. World J. 2012, 2012, 792601. [Google Scholar] [CrossRef] [Green Version]
- van de Wiel, C.; Schaart, J.; Niks, R.; Visser, R. Traditional Plant Breeding Methods; Wageningen UR Plant Breeding: Wageningen, The Netherlands, 2010. [Google Scholar]
- Madon, M. Short communication: Genomic constitution of oil palm interspecific hybrid crosses monitored by genomic in situ hybridisation (gish). J. Oil Palm Res. 2018, 30, 338–344. [Google Scholar] [CrossRef] [Green Version]
- Camillo, J.; Leão, A.P.; Alves, A.A.; Formighieri, E.F.; Azevedo, A.L.; Nunes, J.D.; de Capdeville, G.; de A Mattos, J.K.; Souza, M.T., Jr. Reassessment of the Genome Size in Elaeis guineensis and Elaeis oleifera, and Its Interspecific Hybrid. Genom. Insights 2014, 7, 13–22. [Google Scholar] [CrossRef] [Green Version]
- Maria, M.; Clyde, M.M.; Cheah, S.C. Cytological analysis of Elaeis guineensis (tenera) chromosomes. Elaeis 1995, 7, 122–131. [Google Scholar]
- Singh, R.; Ong-Abdullah, M.; Low, E.-T.L.; Manaf, M.A.A.; Rosli, R.; Nookiah, R.; Ooi, L.C.-L.; Ooi, S.-E.; Chan, K.-L.; Halim, M.A.; et al. Oil palm genome sequence reveals divergence of interfertile species in Old and New worlds. Nature 2013, 500, 335–339. [Google Scholar] [CrossRef] [Green Version]
- Schmutz, J.; Cannon, S.B.; Schlueter, J.; Ma, J.; Mitros, T.; Nelson, W.; Hyten, D.L.; Song, Q.; Thelen, J.J.; Cheng, J.; et al. Genome sequence of the palaeopolyploid soybean. Nature 2010, 463, 178–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chalhoub, B.; Denoeud, F.; Liu, S.; Parkin, I.A.; Tang, H.; Wang, X.; Chiquet, J.; Belcram, H.; Tong, C.; Samans, B.; et al. Plant genetics. Early allopolyploid evolution in the post-Neolithic Brassica napus oilseed genome. Science 2014, 345, 950–953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schnable, P.S.; Ware, D.; Fulton, R.S.; Stein, J.C.; Wei, F.; Pasternak, S.; Liang, C.; Zhang, J.; Fulton, L.; Graves, T.A.; et al. The B73 maize genome: Complexity, diversity, and dynamics. Science 2009, 326, 1112–1115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, Y.; Xu, P.; Fan, H.; Baudouin, L.; Xia, W.; Bocs, S.; Xu, J.; Li, Q.; Guo, A.; Zhou, L.; et al. The genome draft of coconut (Cocos nucifera). Gigascience 2017, 6, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Denis, M.; Bouvet, J.-M. Efficiency of genomic selection with models including dominance effect in the context of Eucalyptus breeding. Tree Genet. Genomes 2013, 9, 37–51. [Google Scholar] [CrossRef]
- Rival, A.; Bertrand, L.; Beulé, T.; Combes, M.C.; Trouslot, P.; Lashermes, P. Suitability of RAPD analysis for the detection of somaclonal variants in oil palm (Elaeis guineensis Jacq). Plant Breed. 1998, 117, 73–76. [Google Scholar] [CrossRef]
- Rohani, P.; Wearing, H.J.; Cameron, T.; Sait, S.M. Natural enemy specialization and the period of population cycles. Ecol. Lett. 2003, 6, 381–384. [Google Scholar] [CrossRef] [Green Version]
- Rival, A.; Ilbert, P.; Labeyrie, A.; Torres, E.; Doulbeau, S.; Personne, A.; Dussert, S.; Beulé, T.; Durand-Gasselin, T.; Tregear, J.W.; et al. Variations in genomic DNA methylation during the long-term in vitro proliferation of oil palm embryogenic suspension cultures. Plant Cell Rep. 2013, 32, 359–368. [Google Scholar] [CrossRef]
- Ooi, S.-E.; Choo, C.-N.; Ishak, Z.; Ong-Abdullah, M. A candidate auxin-responsive expression marker gene, EgIAA9, for somatic embryogenesis in oil palm (Elaeis guineensis Jacq.). Plant Cell. Tissue Organ Cult. 2012, 110, 201–212. [Google Scholar] [CrossRef]
- Yunus, A.M.M.; Fischer, R. Development of a Protoplast based Transformation System for Genetic Engineering of Oil Palm; Publikationsserver der RWTH Aachen University: Aachen, Germany, 2013. [Google Scholar]
- Bahariah, B.; Mat Yunus, A.M.; Rasid, O.; Ghulam Kadir, A.P. Multiplex CRISPR/Cas9-mediated genome editing of the FAD2 gene in rice: A model genome editing system for oil palm. J. Genet. Eng. Biotechnol. 2021, 19, 86. [Google Scholar] [CrossRef]
- Sambanthamurthi, R.; Abdullah, S.; Ghulam Kadir, A.P. Genetic manipulation of the oil palm—Challenges and prospects. Planter 2002, 78, 547–562. [Google Scholar]
- Ghulam Kadir, A.P.; Rasid, O.; Hashim, A.; Ishak, Z.; Rosli, S.; Sambanthamurthi, R. Tissue Culture and Genetic Engineering of Oil Palm. In Palm Oil: Production, Processing, Characterization, and Uses; Science Direct; AOCS Press: Urbana, IL, USA, 2012; pp. 87–135. [Google Scholar] [CrossRef]
- Bahariah, B.; Ghulam Kadir, A.P.; Mat Yunus, A.M.; Subhi, S.; Khalid, N.; Othman, D.R. Biolistic transformation of oil palm using the phosphomannose isomerase (pmi) gene as a positive selectable marker. Biocatal. Agric. Biotechnol. 2013, 2, 295–304. [Google Scholar] [CrossRef]
- Masani, M.Y.A.; Noll, G.A.; Parveez, G.K.A.; Sambanthamurthi, R.; Prüfer, D. Efficient Transformation of Oil Palm Protoplasts by PEG-Mediated Transfection and DNA Microinjection. PLoS ONE 2014, 9, e96831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mat Yunus, A.M.; Ghulam Kadir, A.P. Development of transformation vectors for the production of potentially high oleate transgenic oil palm. Electron. J. Biotechnol. 2008, 11, 23–31. [Google Scholar] [CrossRef]
- Izawati, A.M.; Parveez, G.K.; Masani, M.Y. Transformation of oil palm using Agrobacterium tumefaciens. In Transgenic Plants; Methods in Molecular Biology; Dunwell, J., Wetten, A., Eds.; Humana Press: Totowa, NJ, USA, 2012; Volume 847, pp. 177–188. [Google Scholar] [CrossRef]
- Abdullah, R.; Zainal, A.; Heng, W.; Li, L.; Beng, Y.; Phing, L.; Abdullah Sirajuddin, S.; Yap, W.; Joseph, J.; Jusoh, S.A.; et al. Immature embryo: A useful tool for oil palm (Elaeis guineensis Jacq.) genetic transformation studies. Electron. J. Biotechnol. 2005, 8, 8. [Google Scholar] [CrossRef] [Green Version]
- Bhore, S.; Shah, F.H. Genetic Transformation of the American Oil Palm (Elaeis oleifera) Immature Zygotic Embryos with Antisense Palmitoyl-Acyl Carrier Protein Thioesterase (PATE) Gene. World Appl. Sci. J. 2012, 16, 362–369. [Google Scholar]
- Lee, M.-P.; Yeun, L.-H.; Abdullah, R. Expression of Bacillus thuringiensis insecticidal protein gene in transgenic oil palm. Electron. J. Biotechnol. 2006, 9, 117–126. [Google Scholar] [CrossRef] [Green Version]
- Ismail, I.; Fong, S.L.; Abdullah, R.; Fei, C.; Zainal, Z.; Sidik, N. Molecular and Expression Analysis of Cowpea Trypsin Inhibitor (CpTI) Gene in Transgenic Elaeis guineensis Jacq Leaves. Aust. J. Crop Sci. 2010, 4, 37–48. [Google Scholar]
- Ariffin, N.; Abdullah, R.; Muad, M.; Lourdes, J.; Emran, N.; Ismail, M.; Ismail, I.; Fadzil, M.; Kong, L.; Siddiqui, Y.; et al. Construction of expression vectors of polyhydroxybutyrate-co-hydroxyvalerate (PHBV) and transient expression of transgenes in immature oil palm embryos. Plasmid 2011, 66, 136–143. [Google Scholar] [CrossRef]
- Bahariah, B.; Rasid, O.; Rahmah, S.; Ghulam Kadir, A.P.; Mat Yunus, A.M. Genetic transformation of oil palm based on selection with hygromycin. J. Oil Palm Res. 2020, 33, 577–587. [Google Scholar]
- Yarra, R.; Jin, L.; Zhao, Z.; Cao, H. Progress in Tissue Culture and Genetic Transformation of Oil Palm: An Overview. Int. J. Mol. Sci. 2019, 20, 5353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Izawati, A.M.D.; Masani, M.Y.A.; Ismanizan, I.; Parveez, G.K.A. Evaluation on the effectiveness of 2-deoxyglucose-6-phosphate phosphatase (DOGR1) gene as a selectable marker for oil palm (Elaeis guineensis Jacq.) embryogenic calli transformation mediated by Agrobacterium tumefaciens. Front. Plant Sci. 2015, 6, 727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, P.; Nagarajan, A.; Uchil, P.D. Electroporation. Cold Spring Harb. Protoc. 2019, 2019, 519–525. [Google Scholar] [CrossRef] [Green Version]
- Mariani, T.S.; Ermavitalini, D.; Mitsutaka, T.; Chia, T.F.; Miyake, H. GUS gene expression in somatic embryo of oil palm (Elaeis guineensis Jacq.). Asian J. Appl. Sci. 2015, 3, 649–650. [Google Scholar]
- Darmawan, C.; Wiendi, N.; Utomo, C.; Liwang, T. Electroporation-mediated genetic transformation of oil palm (Elaeis guineensis). Biodiversitas J. Biol. Divers. 2020, 21, 3720–3726. [Google Scholar] [CrossRef]
- Govender, N.; Wong, M.-Y. Detection of Oil Palm Root Penetration by Agrobacterium-Mediated Transformed Ganoderma boninense, Expressing Green Fluorescent Protein. Phytopathology 2016, 107, 483–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bahariah, B.; Mat Yunus, A.M.; Rasid, O.; Ghulam Kadir, A.P. Construction of a vector containing hygromycin (HPT) gene driven by double 35S (2XCaMV35S) promoter for oil palm transformation. J. Oil Palm Res. 2017, 29, 180–188. [Google Scholar] [CrossRef] [Green Version]
- Varshney, R.K.; Graner, A.; Sorrells, M.E. Genic microsatellite markers in plants: Features and applications. Trends Biotechnol. 2005, 23, 48–55. [Google Scholar] [CrossRef]
- Singh, R.; Tan, S.G.; Panandam, J.M.; Rahman, R.A.; Ooi, L.C.; Low, E.T.; Cheah, S.C. Mapping quantitative trait loci (qtls) for fatty acid composition in an interspecific cross of oil palm. BMC Plant Biol. 2009, 9, 114. [Google Scholar] [CrossRef] [Green Version]
- Gan, Y.L.; Fu, J.Y.; Lai, O.M.; Chew, B.H.; Yuen, K.H.; Teng, K.T.; Nesaretnam, K.; Selvaduray, K.R.; Meganathan, P. Effect of palm-based tocotrienols and tocopherol mixture supplementation on platelet aggregation in subjects with metabolic syndrome: A randomized controlled trial. Sci. Rep. 2017, 7, 11542. [Google Scholar] [CrossRef] [Green Version]
- Teh, C.K.; Ong, A.L.; Kwong, Q.B.; Apparow, S.; Chew, F.T.; Mayes, S.; Mohamed, M.; Appleton, D.; Kulaveerasingam, H. Genome-wide association study identifies three key loci for high mesocarp oil content in perennial crop oil palm. Sci. Rep. 2016, 6, 19075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sambanthamurthi, R.; Singh, R.; Kadir, A.P.G.; Abdullah, M.O.; Kushairi, A. Opportunities for the Oil Palm via Breeding and Biotechnology. In Breeding Plantation Tree Crops: Tropical Species; Jain, S.M., Priyadarshan, P.M., Eds.; Springer: New York, NY, USA, 2009; pp. 377–421. [Google Scholar] [CrossRef]
- Mayes, S.; Jack, P.; Corley, R. The use of molecular markers to investigate the genetic structure of an oil palm breeding programme. Heredity 2000, 85 Pt 3, 288–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sathish, D.; Mohankuma, R.C. RAPD markers for identifying oil palm (Elaeis guineensis Jacq.) parental varieties (dura & pisifera) and the hybrid tenera. Indian J. Biotechnol. 2007, 6, 354–358. [Google Scholar]
- Singh, R.; Low, E.T.L.; Ooi, L.C.L.; Ong-Abdullah, M.; Ting, N.C.; Nagappan, J.; Nookiah, R.; Amiruddin, M.D.; Rosli, R.; Manaf, M.A.A.; et al. The oil palm SHELL gene controls oil yield and encodes a homologue of SEEDSTICK. Nature 2013, 500, 340–344. [Google Scholar] [CrossRef] [Green Version]
- Moretzsohn, M.C.; Ferreira, M.A.; Amaral, Z.; Coelho, P.; Grattapaglia, D.; Ferreira, M.E. Genetic diversity of brazilian oil palm (elaeis oleifera hbk) germplasm collected in the amazon forest. Euphytica 2002, 124, 35–45. [Google Scholar] [CrossRef]
- Moretzsohn, M.; Nunes, C.; Ferreira, M.; Grattapaglia, D. RAPD linkage mapping of the shell thickness locus in oil palm ( Elaeis guineensis Jacq.). Theor. Appl. Genet. 2000, 100, 63–70. [Google Scholar] [CrossRef]
- Dou, Y.; Xia, W.; Mason, A.S.; Huang, D.; Sun, X.; Fan, H.; Xiao, Y. Developing functional markers for vitamin E biosynthesis in oil palm. PLoS ONE 2021, 16, e0259684. [Google Scholar] [CrossRef]
- Xiao, Y.; Zhou, L.; Xia, W.; Mason, A.S.; Yang, Y.; Ma, Z.; Peng, M. Exploiting transcriptome data for the development and characterization of gene-based ssr markers related to cold tolerance in oil palm (elaeis guineensis). BMC Plant Biol. 2014, 14, 384. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Williams, R.; Prakash, C.S.; He, G. Identification and characterization of gene-based ssr markers in date palm (phoenix dactylifera l). BMC Plant Biol. 2012, 12, 237. [Google Scholar] [CrossRef] [Green Version]
- Billotte, N.; Marseillac, N.; Risterucci, A.M.; Adon, B.; Brottier, P.; Baurens, F.C.; Singh, R.; Herrán, A.; Asmady, H.; Billot, C.; et al. Microsatellite-based high density linkage map in oil palm (Elaeis guineensis Jacq.). Theor. Appl. Genet. 2005, 110, 754–765. [Google Scholar] [CrossRef]
- Babu, B.K.; Mathur, R.K.; Kumar, P.N.; Ramajayam, D.; Ravichandran, G.; Venu, M.V.B.; Babu, S.S. Development, identification and validation of CAPS marker for SHELL trait which governs dura, pisifera and tenera fruit forms in oil palm (Elaeis guineensis Jacq.). PLoS ONE 2017, 12, e0171933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, B.; Wang, L.; Zhang, Y.J.; Lee, M.; Rahmadsyah, R.; Alfiko, Y.; Yue, G.H. Developing genome-wide snps and constructing an ultrahigh-density linkage map in oil palm. Sci. Rep. 2018, 8, 691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grattapaglia, D.; Silva-Junior, O.B.; Resende, R.T.; Cappa, E.P.; Müller, B.S.F.; Tan, B.; Isik, F.; Ratcliffe, B.; El-Kassaby, Y.A. Quantitative Genetics and Genomics Converge to Accelerate Forest Tree Breeding. Front. Plant Sci. 2018, 9, 1693. [Google Scholar] [CrossRef] [PubMed]
- Rafii, M.; Sukaimi, J.; Din, A.; Jalloh, M.; Swaray, S.; Yusuff, O.; Chukwu, S. Genetic Improvement of Oil Palm Through Recurrent Selection. In The Oil Palm Genome; Compendium of Plant Genomes; Springer: Cham, Switzerland, 2020; pp. 35–46. [Google Scholar] [CrossRef]
- Kadam, D.C.; Potts, S.M.; Bohn, M.O.; Lipka, A.E.; Lorenz, A.J. Genomic Prediction of Single Crosses in the Early Stages of a Maize Hybrid Breeding Pipeline. G3 2016, 6, 3443–3453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cros, D.; Bocs, S.; Riou, V.; Ortega-Abboud, E.; Tisné, S.; Argout, X.; Pomiès, V.; Nodichao, L.; Lubis, Z.; Cochard, B.; et al. Genomic preselection with genotyping-by-sequencing increases performance of commercial oil palm hybrid crosses. BMC Genom. 2017, 18, 839. [Google Scholar] [CrossRef] [Green Version]
- El-Mounadi, K.; Morales-Floriano, M.L.; Garcia-Ruiz, H. Principles, Applications, and Biosafety of Plant Genome Editing Using CRISPR-Cas9. Front. Plant Sci. 2020, 11, 56. [Google Scholar] [CrossRef]
- Arora, L.; Narula, A. Gene Editing and Crop Improvement Using CRISPR-Cas9 System. Front. Plant Sci. 2017, 8, 1932. [Google Scholar] [CrossRef] [Green Version]
- Yarra, R.; Cao, H.; Jin, L.; Mengdi, Y.; Zhou, L. CRISPR/Cas mediated base editing: A practical approach for genome editing in oil palm. 3 Biotech 2020, 10, 306. [Google Scholar] [CrossRef]
- Kalyana Babu, B.; Mary Rani, K.L.; Sarika Sahu, R.K.; Mathur Naveen Kumar, P.; Ravichandran, G.; Anitha, P.; Bhagya, H.P. Development andvalidation of whole genome-wide and genic microsatellite markers in oil palm (Elaeis guineensis Jacq.): First microsatellite database (OpSatdb). Sci. Rep. 2019, 9, 1899. [Google Scholar]
- Haun, W.; Coffman, A.; Clasen, B.; Demorest, Z.; Lowy, A.; Ray, E.; Retterath, A.; Stoddard, T.; Juillerat, A.; Cedrone, F.; et al. Improved soybean oil quality by targeted mutagenesis of the fatty acid desaturase 2 gene family. Plant Biotechnol. J. 2014, 12, 934–940. [Google Scholar] [CrossRef]
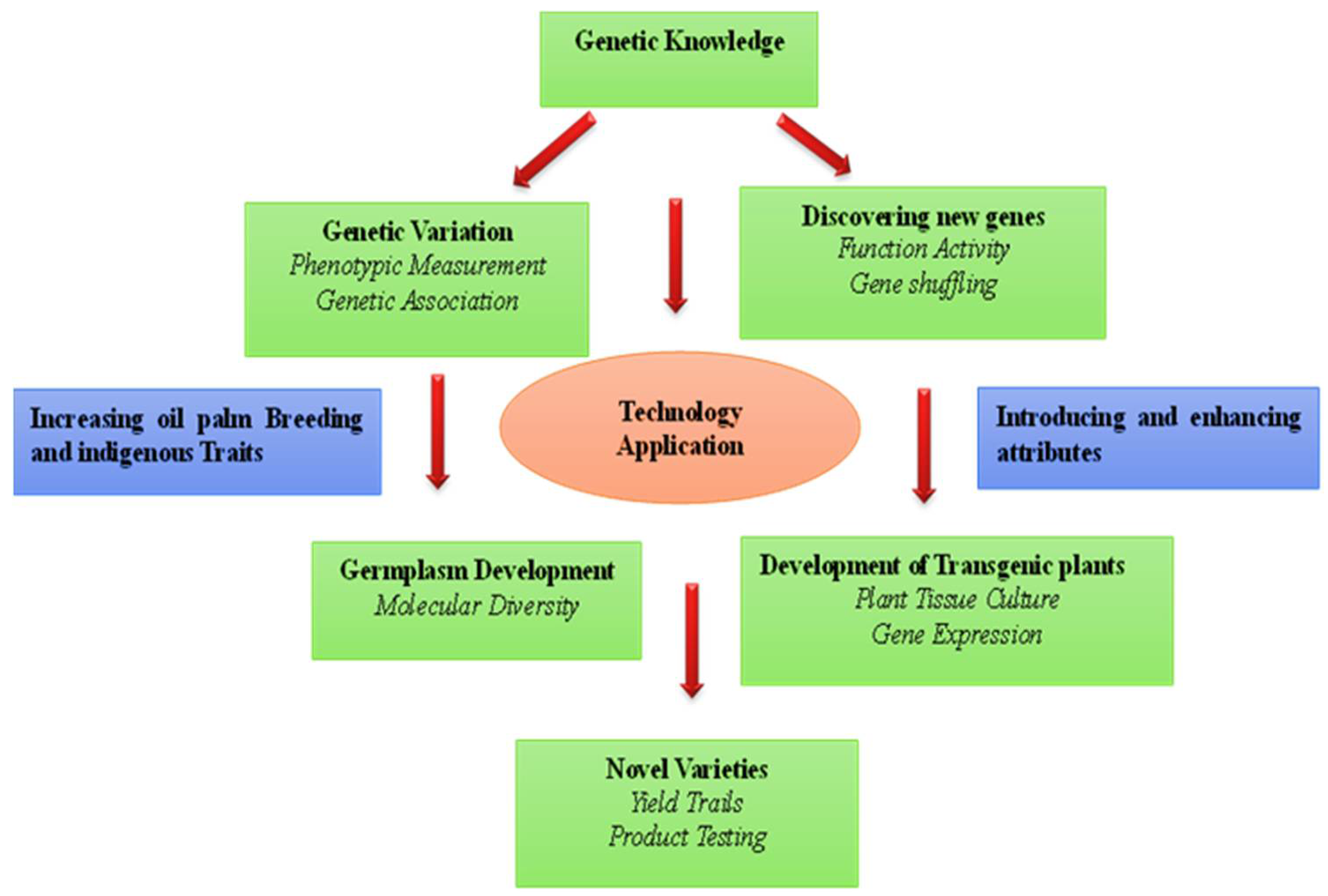
Figure 1.
Tools for oil palm breeding. The left side exhibits the identification of native traits and molecular isolation of genes that reside in the oil palm, although sometimes in wild variants or in low-yielding varieties. The right side depicts discovery of transgenic traits, where we want to change the expression levels, location, or timing of a gene, or to use a gene from a different species.
Figure 1.
Tools for oil palm breeding. The left side exhibits the identification of native traits and molecular isolation of genes that reside in the oil palm, although sometimes in wild variants or in low-yielding varieties. The right side depicts discovery of transgenic traits, where we want to change the expression levels, location, or timing of a gene, or to use a gene from a different species.

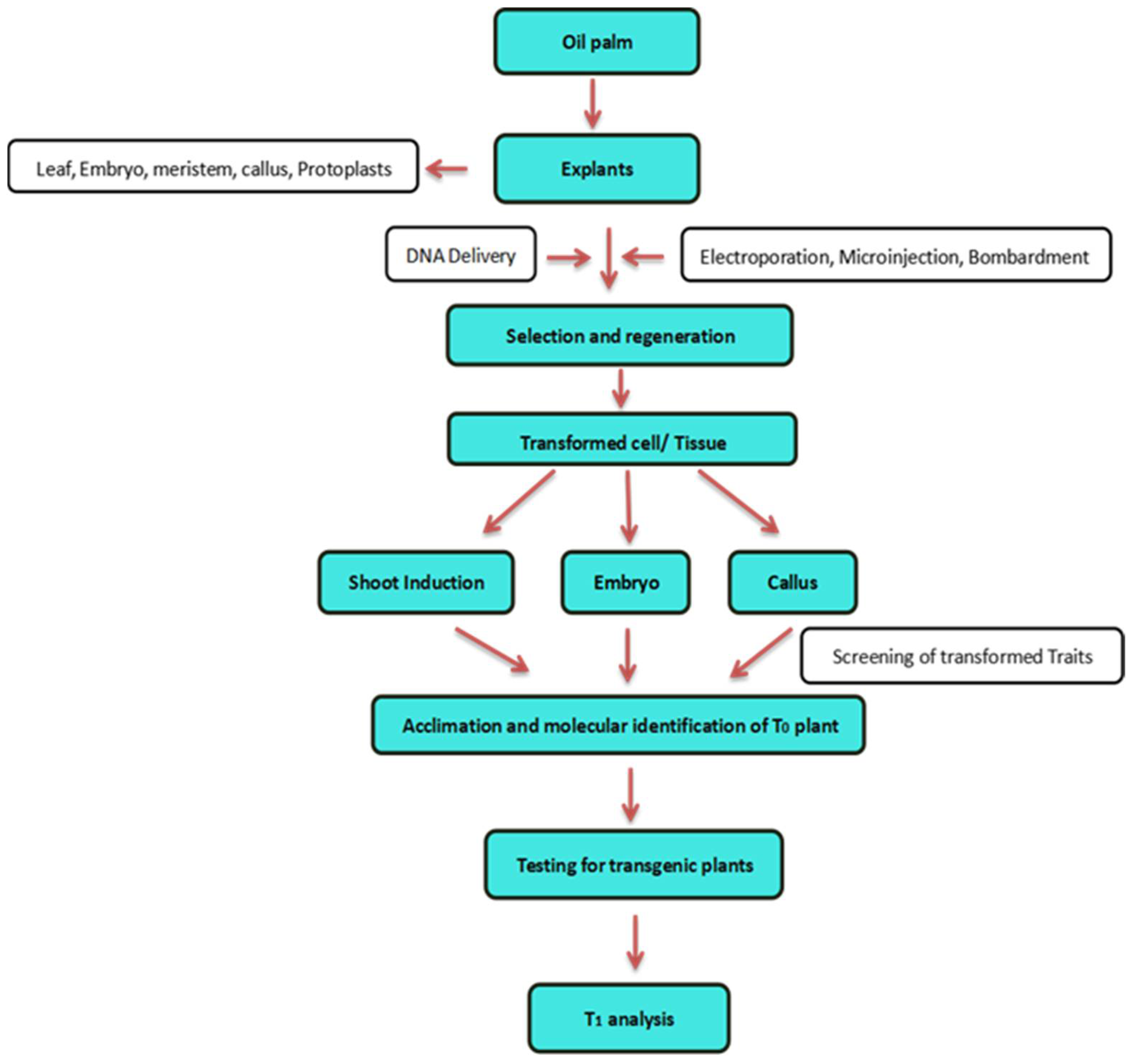
Figure 2.
A summary of the technique for producing transgenic oil palm utilizing the direct DNA delivery method.
Figure 2.
A summary of the technique for producing transgenic oil palm utilizing the direct DNA delivery method.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
John Martin, J.J.; Yarra, R.; Wei, L.; Cao, H. Oil Palm Breeding in the Modern Era: Challenges and Opportunities. Plants 2022, 11, 1395. https://doi.org/10.3390/plants11111395
AMA Style
John Martin JJ, Yarra R, Wei L, Cao H. Oil Palm Breeding in the Modern Era: Challenges and Opportunities. Plants. 2022; 11(11):1395. https://doi.org/10.3390/plants11111395
Chicago/Turabian StyleJohn Martin, Jerome Jeyakumar, Rajesh Yarra, Lu Wei, and Hongxing Cao. 2022. "Oil Palm Breeding in the Modern Era: Challenges and Opportunities" Plants 11, no. 11: 1395. https://doi.org/10.3390/plants11111395
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.