
Improving the Phenolic Content of Tempranillo Grapes by Sustainable Strategies in the Vineyard

 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Results and Discussion
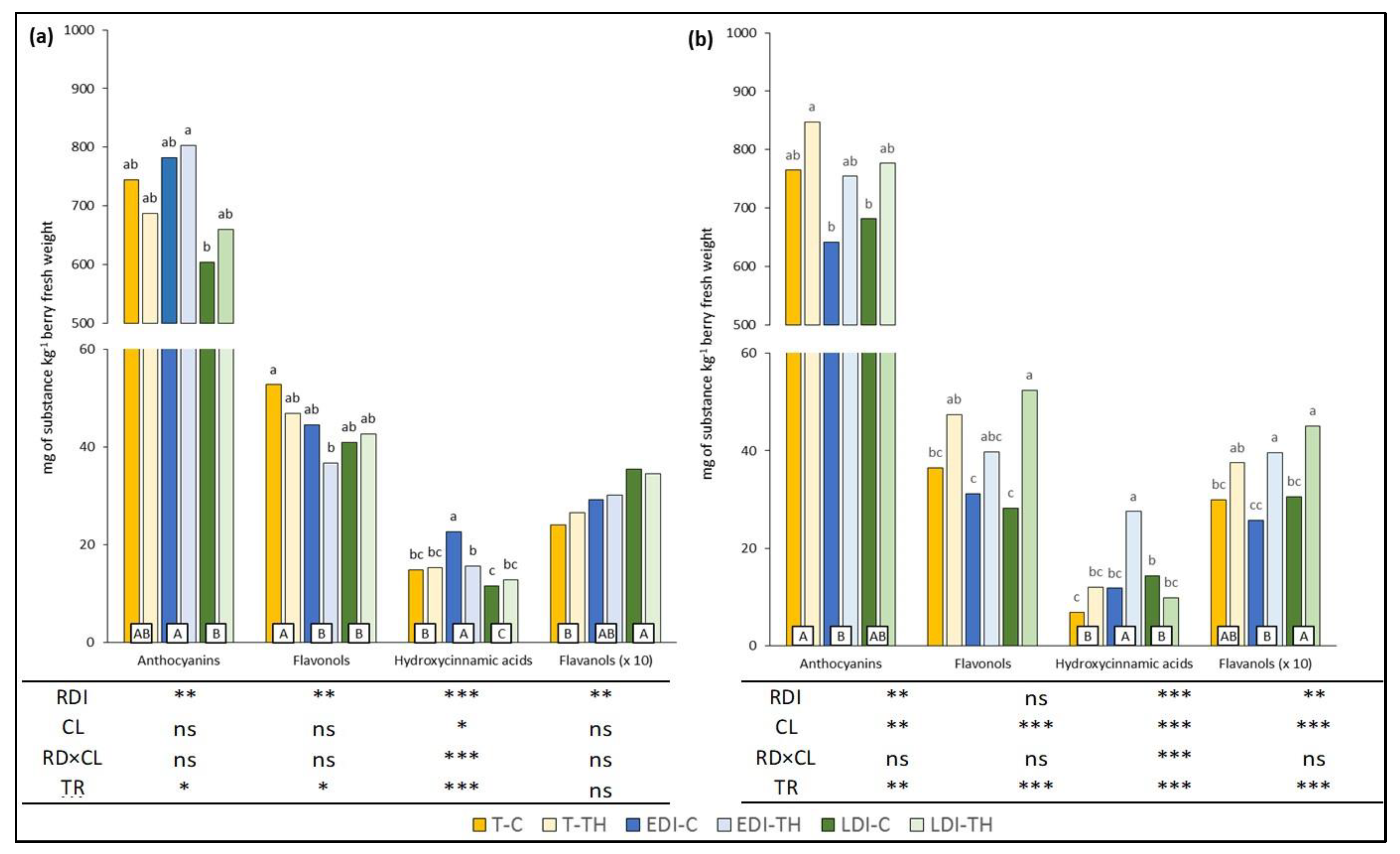
2.1. Grape Skin Total Phenolic Composition
2.2. Grape Skin Individual Phenolic Composition
2.2.1. Anthocyanin Profile
2.2.2. Flavonol Profile
2.2.3. Flavanols
2.2.4. Hydroxycinnamic Compounds and Stilbene
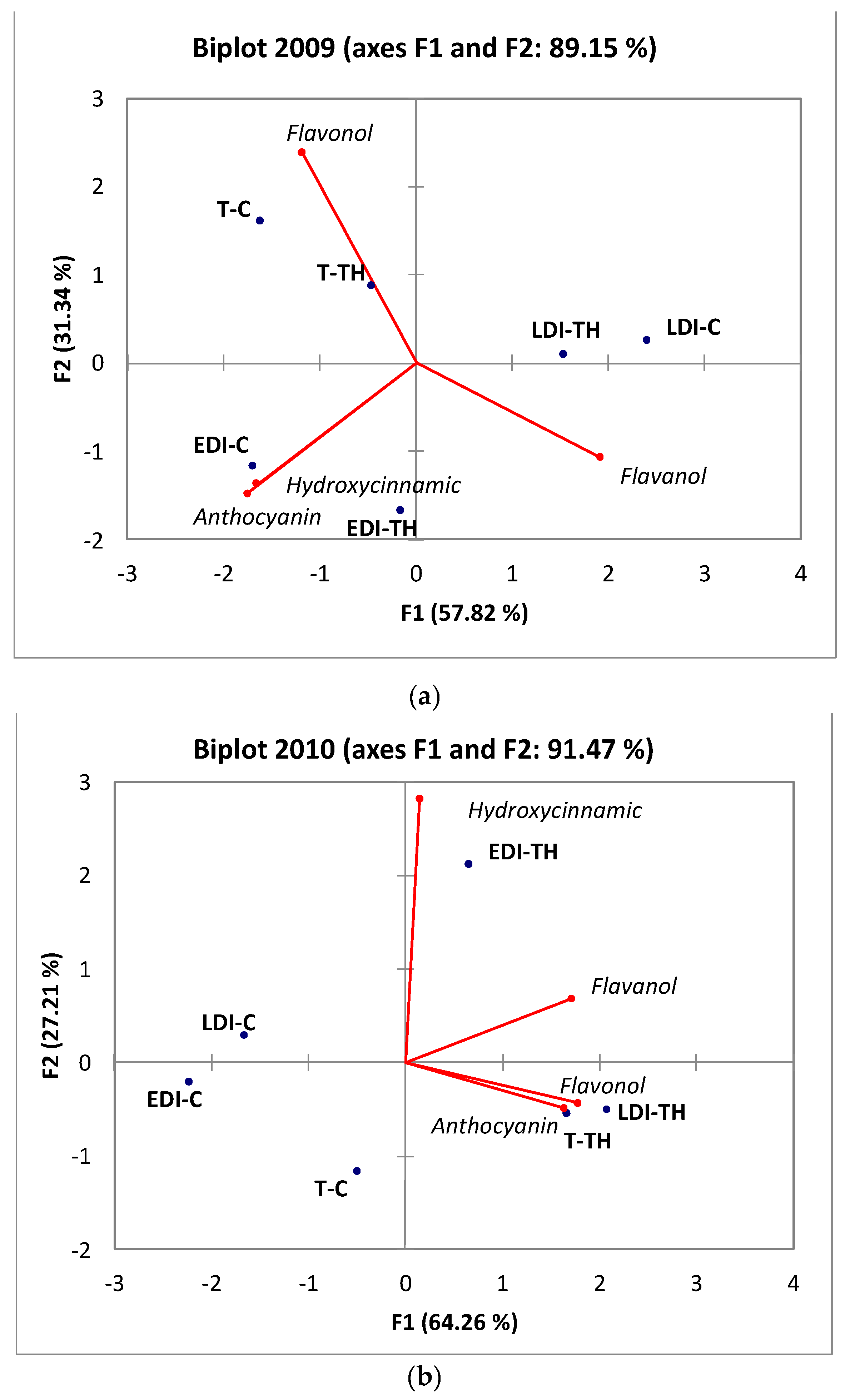
2.2.5. Principal Component Analysis (PCA)
3. Materials and Methods
3.1. Plant Material
3.2. Experimental Design
3.3. Climate Conditions
3.4. Leaf Area
3.5. Phenolic Compounds Analysis
3.6. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Salón, J.L.; Chirivella, C.; Castel, J.R. Response of cv. Bobal to timing of deficit irrigation in Requena, Spain: Water relations, yield, and wine quality. Am. J. Enol. Vitic. 2005, 56, 1–8. [Google Scholar]
- Romero, P.; Gil-Munoz, R.; Amor, F.M.; Valdes, E.; Fernandez, J.I.; Martinez-Cutillas, A. Regulated deficit irrigation based upon optimum water status improves phenolic composition in Monastrell grapes and wines. Agric. Water Manag. 2013, 121, 85–101. [Google Scholar] [CrossRef]
- Zarrouk, O.; Francisco, R.; Pinto-Marijuan, M.; Brossa, R.; Santos, R.R.; Pinheiro, C.; Costa, J.M.; Lopes, C.; Chaves, M.M. Impact of irrigation regime on berry development and flavonoids composition in Aragonez (Syn: Tempranillo) grapevine. Agric. Water Manag. 2012, 114, 18–29. [Google Scholar] [CrossRef]
- Kriedemann, P.E.; Goodwin, I. Regulated Deficit Irrigation and Partial Rootzone Drying: An Overview of Principles and Applications; Land and Water Australia: Canberra, ACT, Australia, 2003. [Google Scholar]
- Basile, B.; Marsal, J.; Mata, M.; Vallverdú, X.; Bellvert, J.; Girona, J. Phenological sensitivity of cabernet sauvignon to water stress: Vine physiology and berry composition. Am. J. Enol. Vitic. 2011, 62, 453–461. [Google Scholar] [CrossRef] [Green Version]
- Intrigliolo, D.S.; Castel, J.R. Response of grapevine cv. “Tempranillo” to timing and amount of irrigation: Water relations, vine growth, yield and berry and wine composition. Irrig. Sci. 2009, 28, 113–125. [Google Scholar] [CrossRef]
- Girona, J.; Marsal, J.; Mata, M.; Campo, J.D.; Basile, B. Phenological sensitivity of berry growth and composition of Tempranillo grapevines (Vitis vinifera L.) to water stress. Aust. J. Grape Wine Res. 2009, 15, 268–277. [Google Scholar] [CrossRef]
- Calderon-Orellana, A.; Mercenaro, L.; Shacke, K.A.; Willits, N.; Matthews, M.A. Responses of fruit uniformity to deficit irrigation and cluster thinning in commercial winegrape production. Am. J. Enol. Vitic. 2014, 65, 354–362. [Google Scholar] [CrossRef]
- Valdés, M.E.; Moreno, D.; Gamero, E.; Uriarte, D.; Prieto, M.D.H.; Manzano, R.; Intrigliolo, D.S. Effects of Cluster Thinning and Irrigation Amount on Water Relations, Growth, Yield and fruit and wine composition of Tempranillo grapes in Extremadura (Spain). J. Int. Sci. Vigne Du Vin. 2009, 43, 67–76. [Google Scholar]
- Pena-Neira, A.; Caceres, A.; Pastenes, C. Low Molecular Weight Phenolic and Anthocyanin Composition of Grape Skins from cv. Syrah (Vitis vinifera L.) in the Maipo Valley (Chile): Effect of Clusters Thinning and Vineyard Yield. Food Sci. Technol. Int. 2007, 13, 153–158. [Google Scholar] [CrossRef]
- Ough, C.S.; Nagaoka, R. Effect of Cluster Thinning and Vineyard Yields on Grape and Wine Composition and Wine Quality of Cabernet Sauvignon. Am. J. Enol. Vitic. 1984, 35, 30–34. [Google Scholar]
- Keller, M.; Mills, L.J.; Wample, R.L.; Spayd, S.E. Cluster Thinning Effects on Three Deficit-Irrigated Vitis vinifera Cultivars. Am. J. Enol. Vitic. 2005, 56, 91–103. [Google Scholar]
- King, P.D.; Smart, R.E.; Mcclellan, D.J. Timing of Crop Removal Has Limited Effect on Merlot Grape and Wine Composition. J. Agric. Sci. 2015, 6, 456–465. [Google Scholar] [CrossRef] [Green Version]
- Diago, M.P.; Vilanova, M.; Tardaguila, J. Effects of Timing of Manual and Mechanical Early Defoliation on the Aroma of Vitis vinifera L. Tempranillo Wine. Am. J. Enol. Vitic. 2010, 3, 382–391. [Google Scholar]
- Fanzone, M.; Zamora, F.; Jofré, V.; Assof, M.; Peña-Neira, Á. Phenolic composition of Malbec grape skins and seeds from Valle de Uco (Mendoza, Argentina) during ripening. Effect of cluster thinning. J. Agric. Food Chem. 2011, 59, 6120–6136. [Google Scholar] [CrossRef]
- Santesteban, L.; Miranda, C.; Royo, J. Thinning intensity and water regime affect the impact cluster thinning has on grape quality. Vitis 2011, 50, 159–165. [Google Scholar]
- Bowen, P.; Bogdanoff, C.; Usher, K.; Estergaard, B.; Watson, M. Effects of Irrigation and Crop Load on Leaf Gas Exchange and Fruit Composition in Red Winegrapes Grown on a Loamy Sand. Am. J. Enol. Vitic. 2011, 62, 9–22. [Google Scholar] [CrossRef]
- Intrigliolo, D.S.; Castel, J.R. Interactive effects of deficit irrigation and shoot and cluster thinning on grapevine cv. Tempranillo. Water relations, vine performance and berry and wine composition. Irrig. Sci. 2011, 29, 443–454. [Google Scholar] [CrossRef]
- Gatti, M.; Bernizzoni, F.; Civardi, S.; Poni, S. Effects of Cluster Thinning and Preflowering Leaf Removal on Growth and Grape Composition in cv. Sangiovese. Am. J. Enol. Vitic. 2012, 63, 325–332. [Google Scholar] [CrossRef]
- Gamero, E.; Moreno, D.; Talaverano, I.; Prieto, M.H.; Guerra, M.T.; Valdés, M.E. Effects of Irrigation and Cluster Thinning on Tempranillo Grape and Wine Composition. S. Afr. J. Enol. Vitic. 2014, 35, 196–204. [Google Scholar] [CrossRef] [Green Version]
- Gamero, E.; Moreno, D.; Vilanova, M.; Uriarte, D.; Prieto, M.H.; Valdés, M.E. Effect of bunch thinning and water stress on chemical and sensory characteristics of Tempranillo wines. Aust. J. Grape Wine Res. 2014, 20, 394–400. [Google Scholar] [CrossRef]
- Uriarte, D.; Intrigliolo, D.S.; Mancha, L.A.; Picon-Toro, J.; Valdes, E.; Prieto, M.H. Interactive Effects of Irrigation and Crop Level on Tempranillo Vines in a Semi-Arid Climate. Am. J. Enol. Vitic. 2015, 66, 101–111. [Google Scholar] [CrossRef]
- Vilanova, M.; Rodríguez, I.; Canosa, P.; Otero, I.; Gamero, E.; Moreno, D.; Talaverano, M.I.; Valdés, E. Variability in chemical composition of Vitis vinifera cv Mencía from different geographic areas and vintages in Ribeira Sacra (NW Spain). Food Chem. 2015, 169, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez Montealegre, R.; Romero Peces, R.; Chacón Vozmediano, J.L.; Martínez Gascueña, J.; García Romero, E. Phenolic compounds in skins and seeds of ten grape Vitis vinifera varieties grown in a warm climate. J. Food Compos. Anal. 2006, 19, 687–693. [Google Scholar] [CrossRef]
- Kennedy, J.A.; Matthews, M.A.; Waterhouse, A.L. Effect of Maturity and Vine Water Status on Grape Skin and Wine Flavonoids. Am. J. Enol. Vitic. 2002, 53, 268–274. [Google Scholar]
- Cadot, Y.; Chevalier, M.; Barbeau, G. Evolution of the localization and composition of phenolics in grape skin between veraison and maturity in relation to water availability and some climatic conditions. J. Sci. Food Agric. 2011, 91, 1963–1976. [Google Scholar] [CrossRef]
- Guidoni, S.; Allara, P.; Schubert, A. Effect of cluster thinning on berry skin anthocyanin composition of Vitis vinifera cv. Nebbiolo. Am. J. Enol. Vitic. 2002, 53, 224–226. [Google Scholar]
- Boulton, R. The Copigmentation of Anthocyanins and Its Role in the Color of Red Wine: A Critical Review. Am. J. Enol. Vitic. 2001, 52, 67–87. [Google Scholar]
- Teixeira, A.; Eiras-Dias, J.; Castellarín, S.; Gerós, H. Berry Phenolics of grapevine under challenging environments. Int. J. Mol. Sci. 2013, 14, 18711–18739. [Google Scholar] [CrossRef] [Green Version]
- Gamero, E.; Espinosa, F.; Moreno, D.; Uriarte, D.; Prieto, M.H.; Garrido, I.; Valdés, M.E. Convenience of Applying of Viticulture Technique as a Function of the Water Status of the Vine-Stock. In Grapes and Wines-Advances in Production, Processing, Analysis and Valorization; Jordao, A.M., Cosme, F., Eds.; InTechOpen: Rijeka, Croatia, 2018; pp. 91–115. [Google Scholar]
- Moreno, D.; Vilanova, M.; Gamero, E.; Intrigliolo, D.S.; Talaverano, M.I.; Uriarte, D.; Esperanza Valdés, M. Effects of preflowering leaf removal on phenolic composition of tempranillo in the semiarid terroir of Western Spain. Am. J. Enol. Vitic. 2015, 66, 204–211. [Google Scholar] [CrossRef] [Green Version]
- Intrigliolo, D.S.; Pérez, D.; Risco, D.; Yeves, A.; Castel, J.R. Yield components and grape composition responses to seasonal water deficits in Tempranillo grapevies. Irrig. Sci. 2012, 30, 339–349. [Google Scholar] [CrossRef]
- Deloire, A.; Carbonneau, A.; Wang, Z.; Ojeda, H. Vine and water: A short review. J. Int. Sci. Vigne Du Vin. 2004, 38, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Ojeda, H.; Andary, C.; Kraeva, E.; Carbonneau, A.; Deloire, A. Influence of pre- and postveraison water deficit on synthesis and concentration of skin phenolic compounds during berry growth of Vitis vinifera cv. Shiraz. Am. J. Enol. Vitic. 2002, 53, 261–267. [Google Scholar]
- Valdés, M.E.; Talaverano, M.I.; Moreno, D.; Prieto, M.H.; Mancha, L.A.; Uriarte, D.; Vilanova, M. Effect of the timing of water deficit on the must amino acid profile of Tempranillo grapes grown under the semiarid conditions of SW Spain. Food Chem. 2019, 292, 24–31. [Google Scholar] [CrossRef]
- Downey, M.O.; Harvey, J.S.; Robinson, S.P. Synthesis of flavonols and expression of flavonol synthase genes in the developing grape berries of Shiraz and Chardonnay (Vitis vinifera L) Aust. J. Grape Wine Res. 2003, 9, 110–121. [Google Scholar] [CrossRef]
- Price, S.F.; Breen, P.J.; Valladao, M.; Watson, B.T. Cluster Sun Exposure and Quercetin in Pinot noir Grapes and Wine. Am. J. Enol. Vitic. 1995, 46, 187–194. [Google Scholar]
- Downey, M.O.; Harvey, J.S.; Robinson, S.P. The effect of bunch shading on berry development and flavonoid accumulation in Shiraz grapes. Aust. J. Grape Wine Res. 2004, 10, 55–73. [Google Scholar] [CrossRef]
- Lee, C.Y.; Jaworski, A. Phenolic Compounds in White Grapes Grown in New York. Am. J. Enol. Vitic. 1987, 38, 277–281. [Google Scholar]
- Revilla, E.; García-Beneytez, E.; Cabello, F. Anthocyanin fingerprint of clones of Tempranillo grapes and wines made with them. Aust. J. Grape Wine Res. 2009, 15, 70–78. [Google Scholar] [CrossRef]
- Moreno, D.; Intrigliolo, D.S.; Vilanova, M.; Castel, J.R.; Gamero, E.; Valdés, E. Phenolic profile of grapevine cv. Tempranillo skins is affected by timing and severity of early defoliation. Span. J. Agric. Res. 2021, 19, e0905. [Google Scholar] [CrossRef]
- Koundouras, S.; Marinos, V.; Gkoulioti, A.; Kotseridis, Y.; Van Leeuwen, C. Influence of vineyard location and vine water status on fruit maturation of nonirrigated cv. Agiorgitiko (Vitis vinifera L.). Effects on wine phenolic and aroma components. J. Agric. Food Chem. 2006, 54, 5077–5086. [Google Scholar] [CrossRef]
- Castellarin, S.D.; Pfeiffer, A.; Sivilotti, P.; Degan, M.; Peterlunger, E.; Di Gaspero, G. Transcriptional regulation of anthocyanin biosynthesis in ripening fruits of grapevine under seasonal water deficit. Plant Cell Environ. 2007, 30, 1381–1399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boss, P.K.; Davies, C.; Robinson, S.P. Analysis of the Expression of Anthocyanin Pathway Genes. Plant Physiol. 1996, 111, 1059–1066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coombe, B.G.; McCarthy, M.G. Dynamics of grape berry growth and physiology of ripening. Aust. J. Grape Wine Res. 2000, 6, 131–135. [Google Scholar] [CrossRef]
- Matus, J.T.; Loyola, R.; Vega, A.; Peña-Neira, A.; Bordeu, E.; Arce-Johnson, P.; Alcalde, J.A. Post-veraison sunlight exposure induces MYB-mediated transcriptional regulation of anthocyanin and flavonol synthesis in berry skins of Vitis vinifera. J. Exp. Bot. 2009, 60, 853–867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres, N.; Hilbert, G.; Luquin, J.; Goicoechea, N.; Antolín, M.C. Flavonoid and amino acid profiling on Vitis vinifera L. cv Tempranillo subjected to deficit irrigation under elevated temperatures. J. Food Compos. Anal. 2017, 62, 51–62. [Google Scholar] [CrossRef]
- Ristic, R.; Downey, M.O.; Iland, P.G.; Bindon, K.; Francis, I.L.; Herderich, M.; Robinson, S.P. Exclusion of sunlight from Shiraz grapes alters wine colour, tannin and sensory properties. Aust. J. Grape Wine Res. 2007, 13, 53–65. [Google Scholar] [CrossRef]
- Martínez-Lüscher, J.; Torres, N.; Hilbert, G.; Richard, T.; Sánchez-Díaz, M.; Delrot, S.; Aguirreolea, J.; Pascual, I.; Gomès, E. Ultraviolet-B radiation modifies the quantitative and qualitative profile of flavonoids and amino acids in grape berries. Phytochemistry 2014, 102, 106–114. [Google Scholar] [CrossRef]
- Adams, D.O. Phenolics and ripening in grape berries. Am. J. Enol. Vitic. 2006, 57, 249–256. [Google Scholar]
- Martínez-Lüscher, J.; Sánchez-Díaz, M.; Delrot, S.; Aguirreolea, J.; Pascual, I.; Gomès, E. Ultraviolet-B alleviates the uncoupling effect of elevated CO2 and increased temperature on grape berry (Vitis vinifera cv. Tempranillo) anthocyanin and sugar accumulation. Aust. J. Grape Wine Res. 2016, 22, 87–95. [Google Scholar] [CrossRef]
- Castellarin, S.D.; Matthews, M.A.; Di Gaspero, G.; Gambetta, G.A. Water deficits accelerate ripening and induce changes in gene expression regulating flavonoid biosynthesis in grape berries. Planta 2007, 227, 101–112. [Google Scholar] [CrossRef]
- Ollé, D.; Guiraud, J.L.; Souquet, J.M.; Terrier, N.; Ageorges, A.; Cheynier, V.; Verries, C. Effect of pre- and post-veraison water deficit on proanthocyanidin and anthocyanin accumulation during Shiraz berry development. Aust. J. Grape Wine Res. 2011, 17, 90–100. [Google Scholar] [CrossRef]
- Savoi, S.; Wong, D.C.; Degu, A.; Herrera, J.C.; Bucchetti, B.; Peterlunger, E.; Fait, A.; Mattivi, F.; Castellarin, S.D. Multi-omics and integrated network analyses reveal new insights into the systems relationships between metabolites, structural genes, and transcriptional regulators in developing grape berries (Vitis vinifera L.) exposed to water deficit. Front. Plant Sci. 2017, 8, 1124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casassa, L.; Keller, M.; Harbertson, J. Regulated deficit irrigation alters anthocyanins, tannins and sensory properties of Cabernet Sauvignon grapes and wines. Molecules 2015, 20, 7820–7844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanlin, R.L.; Hrmova, M.; Harbertson, J.F.; Downey, M.O. Review: Condensed tannin and grape cell wall interactions and their impact on tannin extractability into wine. Aust. J. Grape Wine Res. 2010, 16, 173–188. [Google Scholar] [CrossRef]
- Karoglan, M.; Osrečak, M.; Maslov, L.; Kozina, B. Effect of cluster and berry thinning on Merlot and Cabernet Sauvignon wines composition. Czech J. Food Sci. 2014, 32, 470–476. [Google Scholar] [CrossRef] [Green Version]
- Avizcuri-Inac, J.M.; Gonzalo-Diago, A.; Sanz-Asensio, J.; Martínez-Soria, M.T.; López-Alonso, M.; Dizy-Soto, M.; Fernández-Zurbano, P. Effect of cluster thinning and prohexadione calcium applications on phenolic composition and sensory properties of red wines. J. Agric. Food Chem. 2013, 61, 1124–1137. [Google Scholar] [CrossRef]
- Escribano-Bailón, T.; Álvarez-García, M.; Rivas-Gonzalo, J.G.; Heredia, F.J.; Santos-Buelga, C. Color and stability of pigments derived from the acetaldehyde-mediated condensation between malvidin 3-O-glucoside and (+)-catechin. J. Agric. Food Chem. 2001, 49, 1213–1217. [Google Scholar] [CrossRef]
- Gawel, R. Red wine astringency: A review. Aust. J. Grape Wine Res. 1998, 4, 74–95. [Google Scholar] [CrossRef]
- Niculcea, M.; Martinez-Lapuente, L.; Guadalupe, Z.; Sánchez-Díaz, M.; Ayestarán, B.; Antolín, M.C. Characterization of phenolic composition of Vitis vinifera L. “Tempranillo” and “Graciano” subjected to deficit irrigation during berry development. Vitis 2015, 54, 9–16. [Google Scholar]
- Hochberg, U.; Degu, A.; Cramer, G.R.; Rachmilevitch, S.; Fait, A. Cultivar specific metabolic changes in grapevines berry skins in relation to deficit irrigation and hydraulic behavior. Plant Physiol. Biochem. 2015, 88, 42–52. [Google Scholar] [CrossRef]
- Deluc, L.G.; Decendit, A.; Papastamoulis, Y.; Mérillon, J.M.; Cushman, J.C.; Cramer, G.R. Water deficit increases stilbene metabolism in cabernet sauvignon berries. J. Agric. Food Chem. 2011, 59, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Picón-Toro, J.; González-Dugo, V.; Uriarte, D.; Mancha, L.A.; Testi, L. Effects of canopy size and water stress over the crop coefficient of a “Tempranillo” vineyard in south-western Spain. Irrig. Sci. 2012, 30, 419–432. [Google Scholar] [CrossRef] [Green Version]
- Allen, R.G.; Pruitt, W.O.; Wright, J.L.; Howell, T.A.; Ventura, F.; Snyder, R.; Itenfisu, D.; Steduto, P.; Berengena, J.; Yrisarry, J.B.; et al. A recommendation on standardized surface resistance for hourly calculation of reference ETo by the FAO56 Penman-Monteith method. Agric. Water Manag. 2006, 81, 1–22. [Google Scholar] [CrossRef]
- Moral, F.J.; Rebollo, F.J.; Paniagua, L.L.; García, A. Climatic spatial variability in Extremadura (Spain) based on viticultural bioclimatic indices. Int. J. Biometeorol. 2014, 58, 2139–2152. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Alonso, S.; García-Romero, E.; Hermosín-Gutiérrez, I. HPLC analysis of diverse grape and wine phenolics using direct injection and multidetection by DAD and fluorescence. J. Food Compos. Anal. 2007, 20, 618–626. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| DpG | CyG | PtG | PnG | MvG | ∑G | DpA | CyA | PtA | PnA | MvA | ∑Ac | DpC | CyC | PtC | PnC | MvC | ∑Cm | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2009 | ||||||||||||||||||
| T | 85.5 ab | 9.9 | 85.4 ab | 18.4 | 266.4 ab | 476.9 ab | 4.8 a | 1.1 | 6.5 a | 0.9 | 26.6 a | 40.5 a | 3.2 a | 3.8 | 39.0 | 7.4 | 142.8 | 201.1 |
| EDI | 96.4 a | 13.4 | 92.4 a | 22.8 | 304.4 a | 542.1 a | 4.8 a | 1.2 | 6.2 a | 0.9 | 27.0 a | 40.8 a | 2.9 ab | 4.0 | 38.2 | 7.6 | 152.8 | 210.5 |
| LDI | 75.3 b | 11.2 | 71.7 b | 18.3 | 230.7 b | 417.0 b | 3.8 b | 1.0 | 4.8 b | 0.7 | 22.0 b | 32.9 b | 1.6 b | 3.5 | 33.1 | 6.8 | 132.8 | 182.2 |
| C | 83.4 | 11.2 | 81.4 | 20.1 | 268.4 | 475.6 | 4.1 b | 1.0 | 5.4 b | 0.8 | 24.3 | 36.2 | 2.6 | 3.5 | 35.8 | 7.2 | 145.3 | 199.2 |
| TH | 88.1 | 11.8 | 85.0 | 19.6 | 266.0 | 481.8 | 4.8 a | 1.2 | 6.3 a | 0.9 | 26.1 | 39.9 | 2.6 | 4.0 | 37.7 | 7.4 | 140.2 | 196.7 |
| T-C | 87.1 | 11.5 | 87.1 ab | 21.0 | 273.7 ab | 492.0 ab | 4.6 ab | 1.1 | 6.3 ab | 1.0 | 27.4 | 41.1 ab | 3.9 | 3.8 | 40.4 | 7.7 | 151.4 ab | 212.2 |
| T-TH | 83.8 | 8.3 | 83.7 ab | 15.9 | 259.2 ab | 461.8 ab | 5.0 a | 1.1 | 6.6 a | 0.8 | 25.8 | 39.9 ab | 2.6 | 3.9 | 37.6 | 7.1 | 134.3 ab | 190.1 |
| EDI-C | 98.4 | 13.3 | 92.6 a | 23.3 | 308.7 a | 549.2 a | 4.6 ab | 1.1 | 5.6 ab | 0.8 | 24.7 | 37.3 ab | 2.0 | 3.7 | 35.3 | 7.6 | 143.3 ab | 196.6 |
| EDI-TH | 94.4 | 13.5 | 92.2 a | 22.2 | 300.0 a | 535.1 a | 5.1 a | 1.2 | 6.8 a | 1.1 | 29.4 | 44.4 a | 3.7 | 4.3 | 41.0 | 7.6 | 162.3 a | 224.3 |
| LDI-C | 64.6 | 8.8 | 64.3 b | 15.9 | 222.7 b | 385.6 b | 3.0 b | 0.9 | 4.2 b | 0.5 | 21.0 | 30.2 b | 1.8 | 3.1 | 31.7 | 6.2 | 141.4 ab | 188.8 |
| LDI-TH | 86.0 | 13.5 | 79.1 ab | 20.7 | 238.7 ab | 448.5 ab | 4.5 ab | 1.2 | 5.4 ab | 1.0 | 23.0 | 35.6 ab | 1.4 | 3.9 | 34.5 | 7.4 | 124.2 b | 175.7 |
| Significance | ||||||||||||||||||
| RDI | * | ns | * | ns | ** | ** | * | ns | ** | ns | * | * | * | ns | ns | ns | ns | ns |
| CL | ns | ns | ns | ns | ns | ns | * | ns | * | ns | ns | ns | ns | ns | ns | ns | ns | ns |
| RDI*CL | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns |
| Treatment | ns | ns | * | ns | * | * | * | ns | * | ns | ns | * | ns | ns | ns | ns | * | ns |
| Dp | Cy | Pt | Pn | Mv | ∑G | DpA | CyA | PtA | PnA | MvA | ∑Ac | DpC | CyC | PtC | PnC | MvC | ∑Cm | |
| 2010 | ||||||||||||||||||
| T | 69.9 | 8.3 a | 70.8 | 27.0 a | 273.7 a | 460.5 a | 5.1 a | 1.1 | 7.6 a | 1.7 a | 38.8 | 55.1 a | 1.2 | 4.5 a | 49.2 | 9.5 a | 219.4 | 290.9 |
| EDI | 52.8 | 4.5 b | 61.1 | 19.0 ab | 233.9 b | 380.3 b | 4.3 b | 1.0 | 6.7 b | 1.0 b | 35.9 | 49.7 b | 1.2 | 3.7 ab | 45.1 | 8.2 ab | 203.5 | 268.2 |
| LDI | 58.6 | 6.8 ab | 65.7 | 17.0 b | 251.7 ab | 409.6 ab | 4.3 b | 1.0 | 6.5 b | 0.9 b | 35.7 | 49.2 b | 1.0 | 3.5 b | 45.6 | 7.7 b | 206.2 | 270.4 |
| C | 49.8 b | 5.0 b | 53.0 | 19.1 | 235.3 b | 371.0 b | 4.0 b | 0.9 | 6.6 b | 1.1 | 36.1 | 49.6 b | 1.1 | 3.4 b | 44.9 | 7.8 b | 211.9 | 275.7 |
| TH | 71.0 a | 8.1 a | 78.8 | 22.9 | 270.9 a | 462.6 a | 5.1 a | 1.2 | 7.2 a | 1.3 | 37.5 | 53.1 a | 1.2 | 4.4 a | 48.3 | 9.1 a | 207.5 | 277.3 |
| T-C | 57.6 | 6.0 bc | 59.4 ab | 25.1 | 259.2 ab | 417.1 abc | 4.5 ab | 0.9 | 7.3 a | 1.7 a | 38.7 | 54.0 ab | 1.1 | 4.3 ab | 48.7 | 9.4 a | 223.4 | 294.0 |
| T-TH | 82.2 | 10.5 a | 82.2 a | 28.9 | 288.2 a | 503.8 a | 5.7 a | 1.3 | 7.8 a | 1.7 a | 38.8 | 56.2 a | 1.3 | 4.8 a | 49.7 | 9.6 a | 215.5 | 287.7 |
| EDI-C | 43.7 | 3.9 c | 48.6 b | 16.3 | 207.2 b | 327.5 c | 3.7 b | 0.9 | 6.5 ab | 0.8 c | 35.1 | 47.8 ab | 1.1 | 3.0 b | 43.2 | 7.1 b | 205.9 | 266.6 |
| EDI-TH | 61.8 | 5.2 bc | 73.5 ab | 21.7 | 260.6 ab | 433.1 abc | 4.9 ab | 1.1 | 6.9 ab | 1.2 abc | 36.7 | 51.6 ab | 1.2 | 4.5 a | 47.1 | 9.4 a | 201.1 | 269.8 |
| LDI-C | 48.1 | 5.1 bc | 50.9 b | 16.0 | 239.4 ab | 368.3 bc | 3.9 b | 0.9 | 6.0 b | 0.8 bc | 34.4 | 46.9 b | 1.0 | 2.9 b | 42.9 | 6.9 b | 206.4 | 266.5 |
| LDI-TH | 69.2 | 8.5 ab | 80.6 ab | 18.0 | 263.9 ab | 450.8 ab | 4.7 ab | 1.1 | 6.9 ab | 1.0 abc | 37.1 | 51.6 ab | 1.0 | 4.0 ab | 48.3 | 8.4 ab | 206.0 | 274.3 |
| Significance | ||||||||||||||||||
| RDI | ns | *** | ns | * | * | * | * | ns | ** | ** | ns | * | ns | ** | ns | * | ns | ns |
| CL | * | *** | *** | ns | ** | *** | ** | ns | * | ns | ns | * | ns | ** | ns | * | ns | ns |
| RDI*CL | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns |
| Treatment | ns | *** | *** | ns | * | ** | ** | ns | ** | ** | ns | * | ns | ** | ns | * | ns | ns |
| My | Qc | Kp | Ih | |
|---|---|---|---|---|
| 2009 | ||||
| T | 30.3 | 10.3 a | 2.6 a | 4.6 a |
| EDI | 24.7 | 8.6 b | 1.6 b | 3.8 b |
| LDI | 26.3 | 8.3 b | 1.9 ab | 3.3 b |
| C | 29.2 a | 9.0 | 1.8 | 4.0 |
| TH | 25.0 b | 9.2 | 2.2 | 3.8 |
| T-C | 33.7 | 9.7 ab | 2.1 ab | 4.8 a |
| T-TH | 26.9 | 11.0 a | 3.0 ab | 4.3 ab |
| EDI-C | 27.7 | 9.2 ab | 1.5 b | 4.2 ab |
| EDI-TH | 21.8 | 8.0 b | 1.7 ab | 3.5 bc |
| LDI-C | 26.2 | 8.1 b | 1.7 ab | 3.1 c |
| LDI-TH | 26.4 | 8.6 ab | 2.0 ab | 3.6 bc |
| Significance | ||||
| RDI | ns | ** | * | *** |
| CL | * | ns | ns | ns |
| RDI*Cl | ns | ns | ns | * |
| Treatment | ns | * | * | ** |
| 2010 | ||||
| T | 24.4 a | 9.4 | 1.3 | 4.9 a |
| EDI | 19.7 b | 8.7 | 1.2 | 4.3 b |
| LDI | 22.8 ab | 9.3 | 1.6 | 4.8 ab |
| C | 17.6 b | 7.5 b | 1.2 b | 4.2 b |
| TH | 26.9 a | 10.8 a | 1.6 a | 5.1 a |
| T-C | 19.8 b | 9.2 | 1.2 ab | 4.6 ab |
| T-TH | 28.9 a | 9.6 | 1.4 ab | 5.2 a |
| EDI-C | 16.6 b | 7.9 | 1.2 ab | 4.0 b |
| EDI-TH | 22.8 ab | 9.4 | 1.2 ab | 4.6 ab |
| LDI-C | 16.5 b | 5.3 | 1.1 b | 4.1 b |
| LDI-TH | 29.1 a | 13.4 | 2.1 a | 5.5 a |
| Significance | ||||
| RDI | * | ns | ns | * |
| CL | *** | * | * | *** |
| RDI*CL | ns | ns | ns | ns |
| Treatment | *** | ns | * | *** |
| CAT | EPI | PB1 | PB2 | PB3 | |
|---|---|---|---|---|---|
| 2009 | |||||
| T | 1.43 b | 0.03 b | 1.37 c | 0.44 b | 2.96 b |
| EDI | 1.87 ab | 0.04 b | 2.11 b | 0.60 ab | 3.53 ab |
| LDI | 2.26 a | 0.26 a | 2.63 a | 0.80 a | 3.95 a |
| C | 1.74 | 0.06 b | 1.90 b | 0.67 | 3.66 |
| TH | 1.97 | 0.16 a | 2.18 a | 0.56 | 3.30 |
| T-C | 1.44 b | 0.04 b | 1.27 c | 0.52 ab | 3.26 ab |
| T-TH | 1.41 b | 0.03 b | 1.47 c | 0.37 b | 2.66 b |
| EDI-C | 1.75 ab | 0.00 b | 1.82 bc | 0.66 ab | 3.66 ab |
| EDI-TH | 1.99 ab | 0.08 b | 2.40 ab | 0.53 ab | 3.40 ab |
| LDI-C | 2.03 ab | 0.13 b | 2.60 a | 0.82 a | 4.06 a |
| LDI-TH | 2.50 a | 0.38 a | 2.65 a | 0.79 a | 3.84 a |
| Significance | |||||
| RDI | ** | *** | *** | ** | * |
| CL | ns | * | * | ns | ns |
| RDI*CL | ns | * | ns | ns | ns |
| Treatment | ** | *** | *** | ** | ns |
| 2010 | |||||
| T | 1.54 | 0.41 | 3.25 | 0.41 | 3.46 ab |
| EDI | 1.61 | 0.49 | 3.19 | 0.44 | 3.26 b |
| LDI | 1.86 | 0.57 | 3.46 | 0.37 | 4.14 a |
| C | 1.49 | 0.49 | 2.57 b | 0.38 | 3.11 b |
| TH | 1.85 | 0.49 | 4.03 a | 0.43 | 4.14 a |
| T-C | 1.34 | 0.40 | 2.47 b | 0.39 | 3.46 b |
| T-TH | 1.74 | 0.42 | 4.03 a | 0.42 | 3.47 ab |
| EDI-C | 1.36 | 0.53 | 2.53 b | 0.36 | 2.53 b |
| EDI-TH | 1.86 | 0.45 | 3.84 a | 0.52 | 4.00 ab |
| LDI-C | 1.77 | 0.55 | 2.71 b | 0.40 | 3.34 b |
| LDI-TH | 1.96 | 0.59 | 4.22 a | 0.34 | 4.95 a |
| Significance | |||||
| RDI | ns | ns | ns | ns | * |
| CL | ns | ns | *** | ns | ** |
| RDI*CL | ns | ns | ns | ns | * |
| Treatment | ns | ns | * | ns | ** |
| CF | COU | FE | CF-T | COU-T | FE-T | Trans-R | |
|---|---|---|---|---|---|---|---|
| 2009 | |||||||
| T | 1.2 ab | 3.0 a | 0.7 a | 2.8 a | 7.9 b | 0.7 ab | 0.08 b |
| EDI | 1.3 a | 2.6 ab | 0.4 b | 3.1 a | 12.7 a | 0.9 a | 0.18 a |
| LDI | 0.9 b | 2.2 b | 0.5 ab | 1.9 b | 7.2 b | 0.6 b | 0.10 b |
| C | 1.1 | 2.6 | 0.5 | 2.3 b | 10.7 a | 0.8 | 0.11 |
| TH | 1.2 | 2.6 | 0.6 | 2.9 a | 7.8 b | 0.6 | 0.13 |
| T-C | 1.1 | 3.0 | 0.8 a | 2.6 a | 8.1 b | 0.7 ab | 0.10 b |
| T-TH | 1.2 | 3.0 | 0.7 ab | 3.0 a | 7.8 b | 0.6 ab | 0.07 b |
| EDI-C | 1.2 | 2.8 | 0.3 b | 3.0 a | 16.6 a | 1.0 a | 0.13 b |
| EDI-TH | 1.3 | 2.5 | 0.5 ab | 3.2 a | 8.8 b | 0.7 ab | 0.22 a |
| LDI-C | 0.9 | 1.9 | 0.4 ab | 1.4 b | 7.5 b | 0.6 b | 0.10 b |
| LDI-TH | 1.0 | 2.5 | 0.5 ab | 2.5 a | 7.0 b | 0.6 b | 0.10 b |
| Significance | |||||||
| RDI | * | * | * | *** | *** | ** | *** |
| CL | ns | ns | ns | ** | *** | ns | ns |
| RDI*CL | ns | ns | ns | ns | *** | ns | ** |
| Treatment | ns | ns | * | ** | *** | * | *** |
| 2010 | |||||||
| T | 0.8 b | 1.9 b | 0.5 b | 2.2 b | 4.7 b | 0.2 | 0.06 b |
| EDI | 1.6 a | 6.0 a | 1.5 a | 3.0 a | 9.2 a | 0.2 | 0.11 a |
| LDI | 0.7 b | 6.0 a | 1.2 ab | 1.7 b | 2.8 c | 0.2 | 0.07 b |
| C | 0.6 b | 4.4 | 1.1 | 1.9 b | 3.4 b | 0.2 | 0.09 |
| TH | 1.4 a | 4.9 | 1.0 | 2.9 a | 7.7 a | 0.2 | 0.07 |
| T-C | 0.5 d | 1.4 b | 0.4 b | 1.6 c | 3.4 bc | 0.2 | 0.05 |
| T-TH | 1.2 b | 2.5 b | 0.6 b | 2.9 ab | 5.9 b | 0.3 | 0.07 |
| EDI-C | 1.0 bc | 3.3 b | 1.1 b | 2.4 bc | 4.9 b | 0.3 | 0.11 |
| EDI-TH | 2.3 a | 8.7 a | 1.9 a | 3.7 a | 13.5 a | 0.2 | 0.10 |
| LDI-C | 0.5 cd | 8.5 a | 1.8 a | 1.7 c | 2.0 c | 0.2 | 0.09 |
| LDI-TH | 0.8 bcd | 3.5 b | 0.7 b | 2.1 bc | 3.5 bc | 0.2 | 0.05 |
| Significance | |||||||
| RDI | *** | *** | * | ** | *** | ns | * |
| CL | *** | ns | ns | *** | *** | ns | ns |
| RDI*CL | ** | *** | ns | ns | *** | ns | ns |
| Treatment | *** | *** | * | *** | *** | ns | * |
| Meteorological Data | 2009 | 2010 | |
|---|---|---|---|
| Mean Temperature (°C) | Pre-veraison 1 | 19 | 19 |
| Post-veraison 2 | 25 | 26 | |
| Growing season 3 | 22 | 23 | |
| Rainfall (mm) | Pre-veraison | 85 | 137 |
| Post-veraison | 6 | 5 | |
| Growing season | 91 | 142 | |
| Annual 4 | 402 | 734 | |
| Irrigation (mm) | |||
| EDI | Total | 332 | 207 |
| LDI | Total | 274 | 129 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valdés, M.E.; Talaverano, M.I.; Moreno, D.; Uriarte, D.; Mancha, L.; Vilanova, M. Improving the Phenolic Content of Tempranillo Grapes by Sustainable Strategies in the Vineyard. Plants 2022, 11, 1393. https://doi.org/10.3390/plants11111393
Valdés ME, Talaverano MI, Moreno D, Uriarte D, Mancha L, Vilanova M. Improving the Phenolic Content of Tempranillo Grapes by Sustainable Strategies in the Vineyard. Plants. 2022; 11(11):1393. https://doi.org/10.3390/plants11111393
Chicago/Turabian StyleValdés, M. Esperanza, M. Inmaculada Talaverano, Daniel Moreno, David Uriarte, Luis Mancha, and Mar Vilanova. 2022. "Improving the Phenolic Content of Tempranillo Grapes by Sustainable Strategies in the Vineyard" Plants 11, no. 11: 1393. https://doi.org/10.3390/plants11111393