
Role of Spermidine in Photosynthesis and Polyamine Metabolism in Lettuce Seedlings under High-Temperature Stress


Abstract
:1. Introduction
2. Results
2.1. Effect of Exogenous Spermidine and D-arg on the Growth Indices of Lettuce Seedlings under High-Temperature Stress
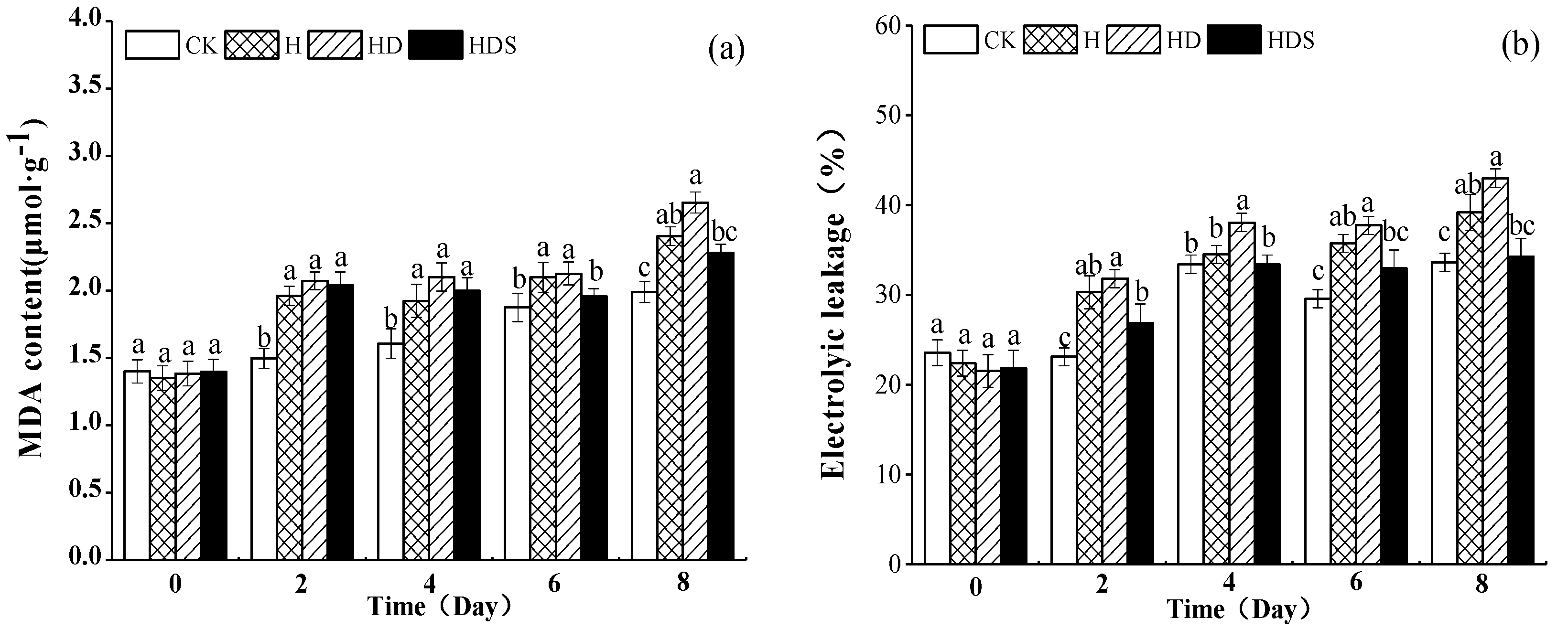
2.2. Effect of Exogenous Spermidine and D-arg on Malondialdehyde Content and Relative Electrolyte Leakage (REL) of Lettuce under High-Temperature Stress
2.3. Effect of Exogenous Spermidine and D-arg on the Chlorophyll Content of Lettuce under High-Temperature Stress
2.4. Effects of Exogenous Spermidine and D-arg on the Photosynthetic Parameters of Lettuce under High-Temperature Stress
2.5. Effect of Exogenous Spermidine and D-arg on Chlorophyll Fluorescence Parameters of Lettuce under High-Temperature Stress
2.6. Effect of Exogenous Spermidine and D-arg on the Endogenous Polyamine Content of Lettuce under High-Temperature Stress
2.7. Effect of Exogenous Spermidine and D-arg on the Polyamine Synthetic and Metabolic Enzyme Activity of Lettuce under High-Temperature Stress
2.8. Effect of Exogenous Spermidine and D-arg on the Gene Expression of Key Enzymes Involved in Endogenous Polyamine Synthesis and Metabolism in Lettuce
3. Discussion
3.1. Effect of Exogenous Spermidine and D-arg on the Growth of Lettuce under High-Temperature Stress
3.2. Effect of Exogenous Spermidine and D-arg on the Cell Membrane Permeability of Lettuce under High-Temperature Stress
3.3. Effect of Exogenous Spermidine and D-arg on Lettuce Photosynthesis under High-Temperature Stress
3.4. Effect of Exogenous Spermidine and D-arg on the Endogenous Polyamine Content of Lettuce under High-Temperature Stress
4. Conclusions
5. Materials and Methods
5.1. Plant Materials and Treatments
5.2. Biomass Production
5.3. Determination of Malondialdehyde and Electrolyte Leakage Rates
5.4. Determination of the Photosynthetic Pigments
- Chl a content (mg·g−1) = [13.95OD665 − 6.88OD649]V/1000W
- Chl b content (mg·g−1) = [24.96OD649 − 7.32OD665]V/1000W
- Chl content (mg·g−1) = [18.08OD649 + 6.63OD665]V/1000 W
- Carotenoid content (mg·g−1) = [1000OD470 − 2.05Chla − 114.8Chlb]/245
5.5. Determination of Photosynthetic and Chlorophyll Fluorescence Parameters
5.6. Determination of Endogenous Polyamine Content
5.7. Determination of Key Enzyme Activity for Polyamine Synthesis and Metabolism
5.8. Total RNA Extraction and SAMDC, ADC, ODC, PAO, and DAO Transcript Level Analysis
5.9. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Paim, B.T.; Crizel, R.L.; Tatiane, S.J.; Rodrigues, V.R.; Galli, V. Mild drought stress has potential to improve lettuce yield and quality. Sci. Hortic. 2020, 272, 109578. [Google Scholar] [CrossRef]
- Simko, I.; Hayes, R.J.; Mou, B.; Mccreight, J.D. Lettuce and spinach. In Proceedings of the EUCARPIA Leafy Vegetables 2003—EUCARPIA Meeting on Leafy Vegetables Genetics and Breeding, Noordwijkerhout, The Netherlands, 19–21 March 2003; Volume 33, pp. 53–86. [Google Scholar] [CrossRef]
- Kim, M.J.; Moon, Y.; Tou, J.C.; Mou, B.; Waterland, N.L. Nutritional value, bioactive compounds and health benefits of lettuce (Lactuca sativa L.). J. Food Compos. Anal. 2016, 49, 19–34. [Google Scholar] [CrossRef]
- Mou, B. Nutrient content of lettuce and its improvement. Curr. Nutr. Food Sci. 2009, 5, 242–248. [Google Scholar] [CrossRef]
- Gong, Z.; Xiong, L.; Shi, H.; Yang, S.; Zhu, J.K. Plant abiotic stress response and nutrient use efficiency. Sci. China Life Sci. 2020, 63, 635–674. [Google Scholar] [CrossRef] [PubMed]
- Gill, D.S.; Raj, T. Engineering temperature tolerance in agricultural crops. Agric. Rev. 2009, 30, 262–282. [Google Scholar]
- Dwivedi, S.K.; Basu, S.; Kumar, S.; Kumar, G.; Prakash, V.; Kumar, S.; Mishra, J.S.; Bhatt, B.P.; Malviya, N.; Singh, G.P. Heat stress induced impairment of starch mobilisation regulates pollen viability and grain yield in wheat: Study in eastern indo-gangetic plains. Field Crop. Res. 2017, 206, 106–114. [Google Scholar] [CrossRef]
- Guo, Y.; Zhang, L.; Li, S.; Cao, Y.; Qi, C.; Wang, J. Advances in molecular biology of vegetable crop response to abiotic stress. Sci. Agric. Sin. 2018, 51, 1167–1181. [Google Scholar] [CrossRef]
- Yang, X.; Han, Y.; Hao, J.; Qin, X.; Liu, C.; Fan, S. Exogenous spermidine enhances the photosynthesis and ultrastructure of lettuce seedlings under high-temperature stress. Sci. Hortic. 2021, 291, 110570. [Google Scholar] [CrossRef]
- Crafts-Brandner, S.J.; Salvucci, M.E. Sensitivity of photosynthesis in a c4 plant, maize, to heat stress. Plant Physiol. 2002, 129, 1773–1780. [Google Scholar] [CrossRef] [Green Version]
- Mathur, S.; Sharma, M.P.; Jajoo, A. Improved photosynthetic efficacy of maize (Zea mays) plants with arbuscular mycorrhizal fungi (amf) under high temperature stress. J. Photochem. Photobiol. B Biol. 2018, 180, 149–154. [Google Scholar] [CrossRef]
- Greer, D.H.; Weedon, M.M. Modelling photosynthetic responses to temperature of grapevine (Vitis vinifera cv. semillon) leaves on vines grown in a hot climate. Plant Cell Environ. 2012, 35, 1365–3040. [Google Scholar] [CrossRef]
- Yang, X.; Chen, X.; Ge, Q.; Li, B.; Tong, Y.; Zhang, A.; Li, Z.H.; Kuang, T.; Lu, C. Tolerance of photosynthesis to photoinhibition, high temperature and drought stress in flag leaves of wheat: A comparison between a hybridization line and its parents grown under field conditions. Plant Sci. 2006, 171, 389–397. [Google Scholar] [CrossRef] [PubMed]
- Cohen, E. Ethylene concentration and the duration of the degreening process in shamouti orange fruit. J. Hortic. Sci. 1978, 5, 139–142. [Google Scholar] [CrossRef]
- Nahar, K.; Hasanuzzaman, M.; Alam, M.M.; Rahman, A.; Suzuki, T.; Fujita, M. Polyamine and nitric oxide crosstalk: Antagonistic effects on cadmium toxicity in mung bean plants through upregulating the metal detoxification, antioxidant defense and methylglyoxal detoxification systems. Ecotoxicol. Environ. Saf. 2016, 126, 245–255. [Google Scholar] [CrossRef]
- Singh, P.; Basu, S.; Kumar, G. Chapter 3—Polyamines metabolism: A way ahead for abiotic stress tolerance in crop plants. In Biochemical, Physiological and Molecular Avenues for Combating Abiotic Stress Tolerance in Plants; Academic Press: Cambridge, MA, USA, 2018; pp. 39–55. [Google Scholar] [CrossRef]
- Evans, P.T.; Malmberg, R.E. Do polyamines have roles in plant development. Annu. Rev. Plant Biol. 1989, 40, 235–242. [Google Scholar] [CrossRef]
- Antognoni, F.; Silvia, F.; Grimmer, C.; Komor, E.; Bagni, N. Long-distance translocation of polyamines in phloem and xylem of ricinus communis plants. Planta 1998, 204, 520–527. [Google Scholar] [CrossRef]
- Kasukabe, Y.; He, L.; Nada, K.; Misawa, S.; Ihara, I.; Tachibana, S. Overexpression of spermidine synthase enhances tolerance to multiple environmental stresses and up-regulates the expression of various stress-regulated genes in transgenic arabidopsis thaliana. Plant Cell Physiol. 2004, 45, 712. [Google Scholar] [CrossRef] [Green Version]
- Kasukabe, Y.; He, L.; Watakabe, Y.; Otani, M.; Shimada, T.; Tachibana, S. Improvement of environmental stress tolerance of sweet potato by introduction of genes for spermidine synthase. Plant Biotechnol. 2006, 23, 75–83. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.; Liu, R.; Han, Y.; Hao, J.; Liu, C.H.J.; Fan, S.H.X. Effects of exogenous spermidine on polyamine metabolism in lettuce (Lactuca sativa L.) under high-temperature stress. Pak. J. Bot. 2021, 53, 1571–1582. [Google Scholar] [CrossRef]
- Groppa, M.D.; Benavides, M.P.; Tomaro, M.L. Polyamine metabolism in sunflower and wheat leaf discs under cadmium or copper stress. Plant Sci. 2003, 164, 293–299. [Google Scholar] [CrossRef]
- Liu, H. Polyamines in Crops Under Water Stress; China Agricultural Press: Beijing, China, 2019; pp. 21–30. [Google Scholar]
- Spormann, S.; Soares, C.; Teixeira, J.; Fidalgo, F. Polyamines as key regulatory players in plants under metal stress—A way for an enhanced tolerance. Ann. Appl. Biol. 2021, 178, 209–226. [Google Scholar] [CrossRef]
- Flores, H.E.; Galston, A.W. Osmotic stress-induced polyamine accumulation in cereal leaves 1. Plant Physiol. 1984, 75, 102–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, C.H.; Sheteiwy, M.S.; Han, J.J.; Dong, Z.R.; Pan, R.H.; Guan, Y.J.; Hamoud, H.Y.Y.; Hu, J. Polyamine biosynthetic pathways and their relation with the cold tolerance of maize (Zea mays L.) seedlings. Plant Signal. Behav. 2020, 15, 1807722. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Yuan, Z.; Wang, B.; Zheng, L.; Tan, J. Exogenous putrescine activates the arginine-polyamine pathway and inhibits the decomposition of endogenous polyamine in anthurium andraeanum under chilling stress. Sci. Hortic. 2021, 282, 110047. [Google Scholar] [CrossRef]
- Anjum, M.A. Response of cleopatra mandarin seedlings to a polyamine-biosynthesis inhibitor under salt stress. Acta Physiol. Plant. 2010, 32, 951–959. [Google Scholar] [CrossRef]
- Ma, X.; Peng, Y.; Huang, L.; Li, Z.H.; Zhang, Y.; Zhang, X.; Merewitz, E.; Yan, H. The alterations of endogenous polyamines and phytohormones induced by exogenous application of spermidine regulate antioxidant metabolism, metallothionein and relevant genes conferring drought tolerance in white clover. Environ. Exp. Bot. 2016, 124, 22–38. [Google Scholar] [CrossRef]
- Fu, Y.; Gu, Q.; Dong, Q.; Zhang, Z.; Lin, C.; Hu, W.; Pan, R.; Guan, Y.; Hu, J. Spermidine enhances heat tolerance of rice seeds by modulating endogenous starch and polyamine metabolism. Molecules 2019, 24, 1395. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Zhang, Z.H.; Wu, B.; Zhang, H. Effect of spermidine and D-arginine treatments on ethylene synthesis in Xinjiang ‘Saimaiti’apricot. Sci. Hortic. 2018, 245, 289–293. [Google Scholar] [CrossRef]
- Hussain, Z.; Singh, Z. Involvement of polyamines in creasing of sweet orange [Citrus sinensis (L.) Osbeck] fruit. Sci. Hortic. 2015, 190, 203–210. [Google Scholar] [CrossRef]
- Li, C.; Liu, R.; Han, Y.; Hao, J.; Liu, C.; Fan, S. Effects of exogenous spermidine on growth and antioxidant enzyme activity of lettuce seedlings under high temperature stress. J. Beijing Agric. Univ. 2019, 34, 56–61. [Google Scholar] [CrossRef]
- Kumar, S.; Kaur, R.; Kaur, N.; Bhandhari, K.; Kaushal, N.; Gupta, K.; Bains, T.; Nayyar, H. Heat-stress induced inhibition in growth and chlorosis in mungbean (Phaseolus aureus Roxb.) is partly mitigated by ascorbic acid application and is related to reduction in oxidative stress. Acta Physiol. Plant 2011, 33, 2091–2101. [Google Scholar] [CrossRef]
- Ahuja, I.; de Vos, R.; Bones, A.; Hall, R. Plant molecular stress responses face climate change. Trends Plant Sci. 2010, 15, 664–674. [Google Scholar] [CrossRef] [PubMed]
- Chang, N.; Zhou, Z.; Li, Y.; Zhang, X. Exogenously applied Spd and Spm enhance drought tolerance in tea plants by increasing fatty acid desaturation and plasma membrane H+-ATPase activity. Plant Physiol. Biochem. 2021, 170, 225–233. [Google Scholar] [CrossRef]
- Tian, J. Study on Physiological Regulation Mechanism and Proteomics of Exogenous Spermidine in Alleviating Injury of Cucumber Seedlings under High Temperature Stress; Nanjing Agricultural University: Nanjing, China, 2012. [Google Scholar]
- Tommasino, E.; Griffa, S.; Grunberg, K.; Ribotta, A.; Colomba, E.L.; Carloni, E.; Quiroga, M.; Luna, C.M. Malondialdehyde content as a potential biochemical indicator of tolerant Cenchrus ciliaris L. genotypes under heat stress treatment. Grass Forage Sci. 2012, 67, 456–459. [Google Scholar] [CrossRef]
- Liu, H.; Lee, Z.X.; Chuang, T.; Wu, H. Effect of heat stress on oxidative damage and antioxidant defense system in white clover (Trifolium repens L.). Planta 2021, 254, 103. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Liu, W.; Jahan, M.; Shu, S.; Sun, J.; Guo, S. Characterization of polyamine oxidase genes in cucumber and roles of CsPAO3 in response to salt stress. Environ. Exp. Bot. 2021, 194, 10469. [Google Scholar] [CrossRef]
- Zhang, X.; Wollenweber, B.; Jiang, D.; Liu, F.; Zhao, J. Water deficits and heat shock effects on photosynthesis of a transgenic Arabidopsis thaliana constitutively expressing ABP9, a bZIP transcription factor. J. Exp. Bot. 2008, 59, 839–848. [Google Scholar] [CrossRef] [Green Version]
- Chauhan, S.; Srivalli, S.; Nautiyal, A.R.; Khanna-Chopra, R. Wheat cultivars differing in heat tolerance show a differential response to monocarpic senescence under high-temperature stress and the involvement of serine proteases. Photosynthetica 2009, 47, 536–547. [Google Scholar] [CrossRef]
- Duan, J. Study on Physiological Regulation of Exogenous Spermidine to Improve Salt Tolerance of Cucumber Seedlings; Nanjing Agricultural University: Nanjing, China, 2008. [Google Scholar]
- Jie, Y.L.; Yang, H.Q.; Cui, M.G.; Luo, X. Relationship between soil water content and water use efficiency of apple leaves. Chin. J. Appl. Ecol. 2001, 12, 387–390. [Google Scholar]
- Zhu, Y.S.; Fan, J.J.; Feng, H. Effects of low light on photosynthetic characteris-tics of tomato at different growth stages. Chin. J. Appl. Ecol. 2010, 21, 3141–3146. [Google Scholar] [CrossRef]
- Behrooz, S.; Chantal, F.; Nasser, G.; Sahebali, B.; Peter, S.; Franz-Werner, B.; Sylvie, C.; Maëva, T.; Andoniaina, D.; Jaleh, G. Stomatal and non-stomatal limitations are responsible in down-regulation of photosynthesis in melon plants grown under the saline condition: Application of carbon isotope discrimination as a reliable proxy. Plant Physiol. Biochem. 2019, 141, 1–19. [Google Scholar] [CrossRef]
- Sun, Q.; Yamada, T.; Han, Y.; Takano, T. Influence of salt stress on C4 photosynthesis in Miscanthus sinensis Anderss. Plant Biol. 2020, 23, 44–56. [Google Scholar] [CrossRef] [PubMed]
- Shu, S.; Yuan, L.; Guo, S.; Sun, J.; Yuan, Y. Effects of exogenous spermine on chlorophyll fluorescence, antioxidant system and ultrastructure of chloroplasts in Cucumis sativus L. under salt stress. Plant Physiol. Biochem. 2013, 63, 209–216. [Google Scholar] [CrossRef]
- Mareckova, M.; Barta, M.; Hajek, J. Temperature effects on photosynthetic performance of Antarctic lichen Dermatocarpon polyphyllizum: A chlorophyll fluorescence study. Polar Biol. 2019, 42, 685–701. [Google Scholar] [CrossRef]
- Thomas, D.S.; Turner, D.W. Banana (Musa sp.) leaf gas exchange and chlorophyll fluorescence in response to soil drought, shading and lamina folding. Sci. Hortic. 2001, 90, 93–108. [Google Scholar] [CrossRef]
- Baker, N.R.; Rosenqvist, E. Applications of chlorophyll fluorescence can improve crop production strategies: An examination of future possibilities. J. Exp. Bot. 2004, 55, 1607–1621. [Google Scholar] [CrossRef] [Green Version]
- Krause, G.H.; Santarius, K.A. Relative thermostability of chloroplast envelope. Planta 1975, 127, 285–299. [Google Scholar] [CrossRef] [PubMed]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence—A practical guide. J. Exp. Bot. 2000, 985, 59–668. [Google Scholar] [CrossRef]
- Pinnola, A.; Dall’Osto, L.; Gerotto, C.; Morosinotto, T.; Bassi, R.; Alboresi, A. Zeaxanthin binds to light-harvesting complex stress-related protein to enhance nonphotochemical quenching in physcomitrella patens. Plant Cell. 2013, 25, 3519–3534. [Google Scholar] [CrossRef] [Green Version]
- Gururani, M.A.; Venkatesh, J.; Tran, L.S.P. Regulation of photosynthesis during abiotic stress-induced photoinhibition. Mol. Plant 2015, 8, 1304–1320. [Google Scholar] [CrossRef] [Green Version]
- Tikkanen, M.; Gollan, P.J.; Mekala, N.R.; Isojarvi, J.; Aro, E.M. Light-harvesting mutants show differential gene expression upon shift to high light as a consequence of photosynthetic redox and reactive oxygen species metabolism. Philos Trans. R. Soc. B Biol. Sci. 2014, 369, 1640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, A.; Taylor, M.; Altabella, T.; Tiburciob, A.F. Recent advances in polyamine research. Trends Plant Sci. 1997, 2, 124–130. [Google Scholar] [CrossRef]
- Chen, D.D.; Shao, Q.S.; Yin, L.H.; Younis, A.; Zheng, B.S. Polyamine function in plants: Metabolism, regulation on development and roles in abiotic stress responses. Front. Plant Sci. 2019, 9, 1945. [Google Scholar] [CrossRef] [PubMed]
- Farooq, M.; Wahid, A.; Lee, D.J. Exogenously applied polyamines increase drought tolerance of rice by improving leaf water status, photosynthesis and membrane properties. Acta Physiol. Plant 2009, 31, 937–945. [Google Scholar] [CrossRef]
- Ebeed, H.T.; Hassan, N.M.; Aljarani, A.M. Exogenous applications of polyamines modulate drought responses in wheat through osmolytes accumulation, increasing free polyamine levels and regulation of polyamine biosynthetic genes. Plant Physiol. Biochem. 2017, 118, 438–448. [Google Scholar] [CrossRef]
- Li, Z.; Zhou, H.; Peng, Y.; Zhang, X.; Ma, X.; Huang, L.; Yan, Y. Exogenously applied spermidine improves drought tolerance in creeping bentgrass associated with changes in antioxidant defense, endogenous polyamines and phytohormones. Plant Growth Regul. 2015, 76, 71–82. [Google Scholar] [CrossRef]
- Collado-González, J.; Piñero, M.C.; Otálora, G.; López-Marín, J.; Amor, F.M. Exogenous spermidine modifies nutritional and bioactive constituents of cauliflower (Brassica oleracea var. botrytis L.) florets under heat stress. Sci. Hortic. 2021, 277, 109818. [Google Scholar] [CrossRef]
- Botella, M.N.; Amor, F.D.; Asunción, A.; María, S.; Antonio, C. Polyamine, ethylene and other physico-chemical parameters in tomato (Lycopersicon esculentum) fruits as affected by salinity. Physiol. Plant. 2000, 109, 428–434. [Google Scholar] [CrossRef]
- Kaur-Sawhney, R.; Dai, Y.R.; Galston, A.W. Effect of inhibitors of polyamine biosynthesis on gibberellin-induced internode growth in light-grown dwarf peas. Plant Cell Physiol. 1986, 27, 253–260. [Google Scholar]
- Hao, Y.; Kitashiba, H.; Honda, C.; Nada, K.; Moriguchi, T. Expression of arginine decarboxylase and ornithine decarboxylase genes in apple cells and stressed shoots. J. Exp. Bot. 2005, 56, 11051115. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Zhu, J.; Sun, L.; Cheng, Y.; Hou, J.; Fan, Y.; Ge, Y. Exogenous γ-aminobutyric acid maintains fruit quality of apples through regulation of ethylene anabolism and polyamine metabolism. Plant Physiol. Biochem. 2021, 169, 92–101. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Cao, D.; Hu, Q.; Guan, Y.; Hu, W.; Nawaz, A.; Hu, J. Soluble protein, sugar, GA3 and sHSPs genes related with seed germination during maturation of hybrid rice seed. J. Sci. Food Agric. 2015, 96, 1764–1771. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X. Crop Physiology Research Method; Agricultural Press: Beijing, China, 1992. [Google Scholar]
- Salah, M.S.; Guan, Y.; Cao, D.; Li, J.; Nawaz, A.; Hu, Q.; Hu, W.; Ning, M.; Hu, J. Seed priming with polyethylene glycol regulating the physio-logical and molecular mechanism in rice (Oryza sativa L.) undernano-ZnO stress. Sci. Rep. 2015, I, 14278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, S.; Zhang, F.; Xu, Y. Establishment of an ultra-high performance liquid chromatographic method for the determination of polyamines in plants. J. Nanjing Agric. Univ. 2016, 39, 8. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Plant Height (cm) | Shoot | Root | Plant Weight (g) | Dry Weight (g) | Water Content (%) | ||
|---|---|---|---|---|---|---|---|---|
| Fresh Weight (g) | Dry Weight (g) | Fresh Weight (g) | Dry Weight (g) | |||||
| CK | 17.93 ± 0.47 b | 14.24 ± 1.02 a | 0.73 ± 0.06 a | 1.78 ± 0.18 a | 0.08 ± 0.01 a | 16.01 ± 1.19 a | 0.81 ± 0.06 a | 95.45 ± 0.001 a |
| H | 18.10 ± 0.67 ab | 8.55 ± 0.59 b | 0.51 ± 0.01 b | 0.50 ± 0.02 b | 0.02 ± 0.00 c | 9.04 ± 0.59 b | 0.54 ± 0.01 b | 94.25 ± 0.001 b |
| HD | 16.20 ± 0.50 b | 7.85 ± 0.18 c | 0.52 ± 0.00 b | 0.48 ± 0.04 b | 0.03 ± 0.00 b | 8.33 ± 0.16 c | 0.55 ± 0.01 b | 93.99 ± 0.001 b |
| HDS | 18.17 ± 0.49 a | 10.56 ± 0.49 ab | 0.66 ± 0.05 a | 0.83 ± 0.09 b | 0.04 ± 0.00 ab | 11.39 ± 0.54 ab | 0.70 ± 0.05 a | 94.88 ± 0.002 b |
| Treatment | Net Photosynthetic Rate (μmol m−2s−1) | Stomatal Conductance (mmol m−2s−1) | Transpiration Rate (mmol m−2s−1) | Intercellular CO2 Concentration (μmol m−2s−1) | Limiting Value of Stomata | Water Use Efficiency |
|---|---|---|---|---|---|---|
| CK | 7.87 ± 0.20 a | 117.00 ± 9.61 ab | 3.14 ± 0.12 a | 313.67 ± 23.62 a | 0.30 ± 0.04 a | 2.21 ± 0.09 b |
| H | 5.60 ± 0.17 b | 105.00 ± 26.51 b | 3.07 ± 0.06 a | 343.33 ± 10.65 a | 0.18 ± 0.02 b | 2.53 ± 0.54 a |
| HD | 3.70 ± 0.26 c | 90.33 ± 19.92 b | 2.22 ± 0.08 a | 350.00 ± 10.02 a | 0.17 ± 0.03 b | 2.57 ± 0.60 a |
| HDS | 4.83 ± 0.19 bc | 119.67 ± 27.72 a | 3.20 ± 0.07 a | 303.33 ± 12.81 a | 0.30 ± 0.01 a | 2.75 ± 0.57 a |
| Treatment | qP | NPQ | ETR | ΦPSⅡ | Fv/Fm |
|---|---|---|---|---|---|
| CK | 0.63 ± 0.03 a | 0.31 ± 0.02 d | 71.12 ± 1.18 a | 0.45 ± 0.09 a | 0.79 ± 0.02 a |
| H | 0.34 ± 0.03 b | 0.69 ± 0.03 b | 64.23 ± 0.91 b | 0.18 ± 0.01 b | 0.70 ± 0.01 b |
| HD | 0.18 ± 0.01 d | 1.11 ± 0.02 a | 40.43 ± 0.95 c | 0.04 ± 0.01 c | 0.66 ± 0.02 c |
| HDS | 0.25 ± 0.01 c | 0.41 ± 0.01 c | 61.29 ± 0.94 b | 0.09 ± 0.00 bc | 0.69 ± 0.02 b |
| Gene Name | Forwards Primer (5′-3′) | Reverse Primer (5′-3′) | Product Size |
|---|---|---|---|
| 18S | GTGAGTGAAGAAGGGCAATG | CACTTTCAACCCGATTCACC | / |
| SAMDC | TACAATGACGACATGGCGGATAT | CTGGTGGTGGCAACGGAAACTG | 80 bp |
| ADC | AATTAGTCCGCCTTGGTGCTTCC | TACTGCCTGAACAACCGCCATTG | 146 bp |
| SPDS | CCTGATGTAGCGGTTGGATACGAAG | CAGTTCTTGTGCGGGACCTATTGG | 144 bp |
| SPS | ACGGAAAGAGACGAGTTTGCCTATC | CACCATCACCACCACCCACAAC | 103 bp |
| PAO | GCTGCTGGGAGATTCGCTTACG | TATTGAACAGGCTCGGTTGCTTCC | 113 bp |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, X.; Hao, J.; Fan, S.; Liu, C.; Han, Y. Role of Spermidine in Photosynthesis and Polyamine Metabolism in Lettuce Seedlings under High-Temperature Stress. Plants 2022, 11, 1385. https://doi.org/10.3390/plants11101385
He X, Hao J, Fan S, Liu C, Han Y. Role of Spermidine in Photosynthesis and Polyamine Metabolism in Lettuce Seedlings under High-Temperature Stress. Plants. 2022; 11(10):1385. https://doi.org/10.3390/plants11101385
Chicago/Turabian StyleHe, Xin, Jinghong Hao, Shuangxi Fan, Chaojie Liu, and Yingyan Han. 2022. "Role of Spermidine in Photosynthesis and Polyamine Metabolism in Lettuce Seedlings under High-Temperature Stress" Plants 11, no. 10: 1385. https://doi.org/10.3390/plants11101385