
Plant Responses to Biotic Stress: Old Memories Matter




Abstract
:1. Introduction
2. Stress Memory in Plants
3. Plant Immunity and Immune Memory
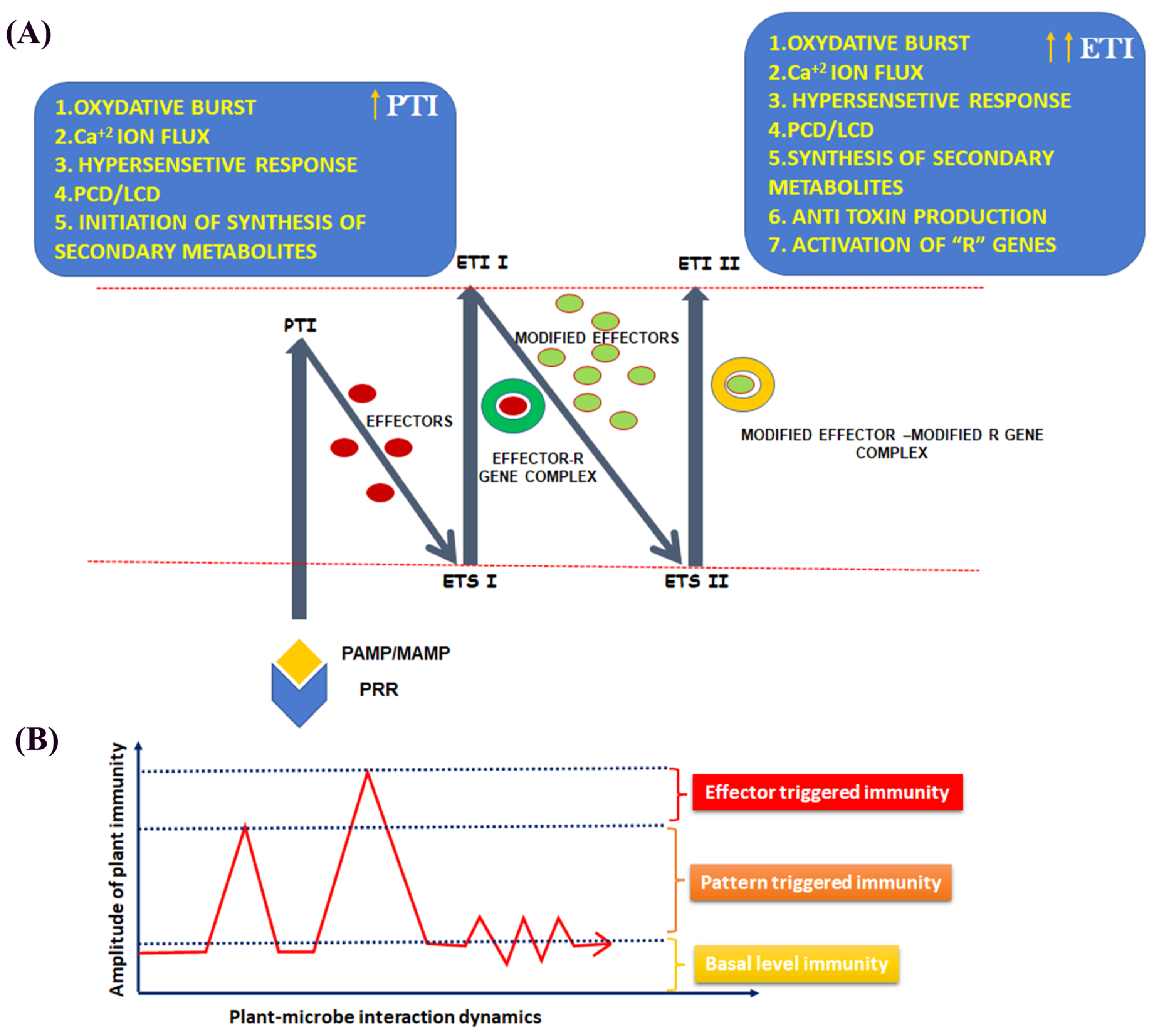
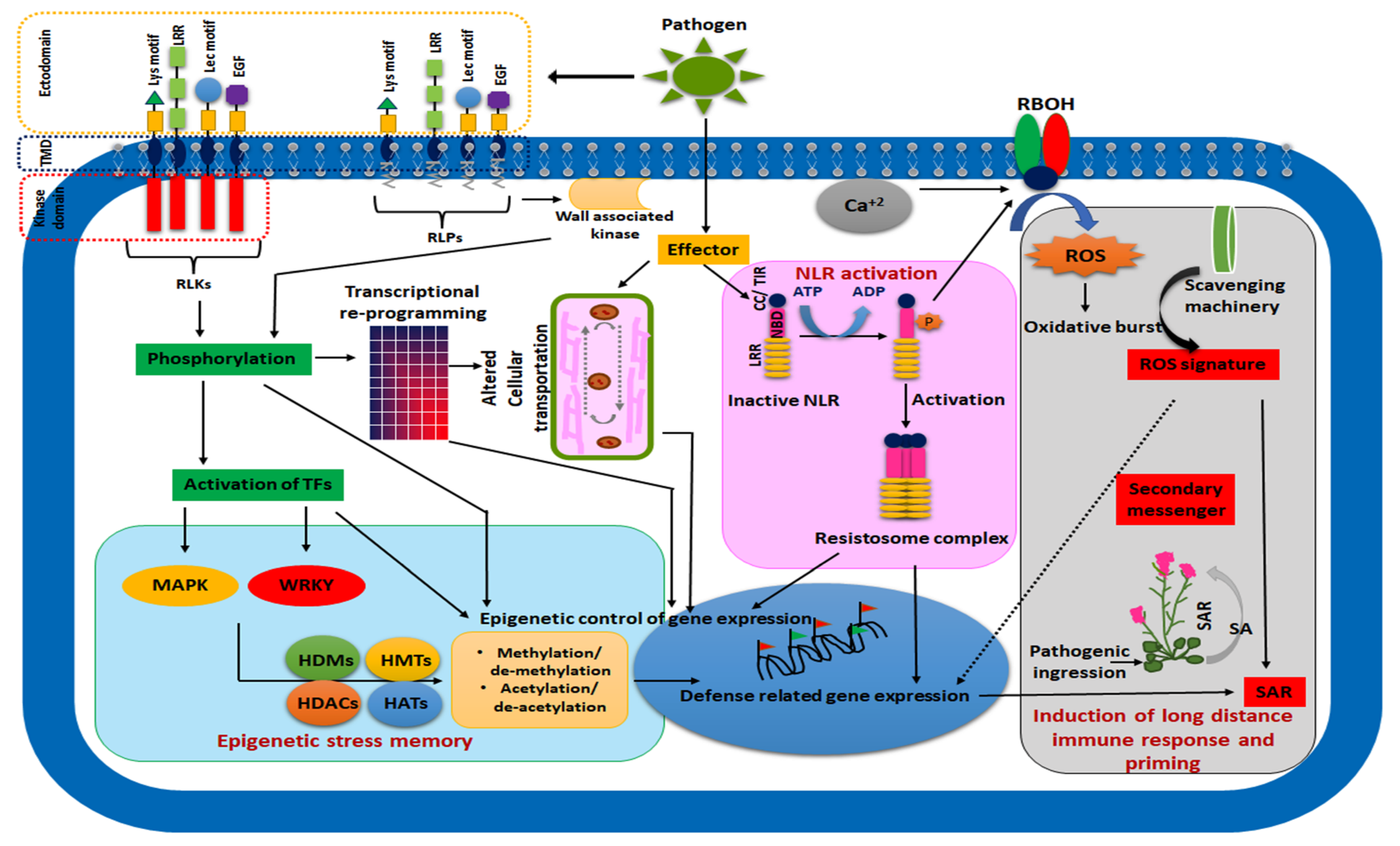
3.1. Architecture of the Plant Immune System
3.2. Immunogenic Memory in Plants: Facts and Reality
4. Epigenetic Regulation of Plant Memory
5. Priming vs. Memory and the Fine Line of the Difference
6. Conclusions and Future Questions
- Is the development of stress memory in plants species specific?
- Can different pathogens make similar modifications in histones?
- What is the actual mechanism of the transgenerational memory?
- How stable is this transgenerational memory with and without the selection of pressure?
- The difference between the genetics of priming and memory is still unclear.
- The basis of the recall of memory has to be adequately resolved.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Walkup, L.K. Junk DNA: Evolutionary discards or God’s tools. Tech. J. 2000, 14, 18–30. [Google Scholar]
- Kornberg, R.D. Eukaryotic transcriptional control. Trends Biochem. Sci. 1999, 24, M46–M49. [Google Scholar] [CrossRef]
- Shanker, A.K.; Bhanu, D.; Maheswari, M. Epigenetics and transgenerational memory in plants under heat stress. Plant Physiol. Rep. 2020, 25, 583–593. [Google Scholar] [CrossRef]
- Espinas, N.A.; Saze, H.; Saijo, Y. Epigenetic control of defense signaling and priming in plants. Front. Plant Sci. 2016, 7, 1201. [Google Scholar] [CrossRef]
- Hilker, M.; Schmülling, T. Stress priming, memory, and signalling in plants. Plant Cell Environ. 2019, 42, 753–761. [Google Scholar] [CrossRef] [PubMed]
- Chester, K.S. The problem of acquired physiological immunity in plants. Q. Rev. Biol. 1933, 8, 275–324. [Google Scholar] [CrossRef]
- Reimer-Michalski, E.-M.; Conrath, U. Innate immune memory in plants. In Seminars in Immunology; Academic Press: Cambridge, MA, USA, 2016; pp. 319–327. [Google Scholar]
- Galviz, Y.C.; Ribeiro, R.V.; Souza, G.M. Yes, plants do have memory. Theor. Exp. Plant Physiol. 2020, 32, 195–202. [Google Scholar] [CrossRef]
- Mladenov, V.; Fotopoulos, V.; Kaiserli, E.; Karalija, E.; Maury, S.; Baranek, M.; Segal, N.; Testillano, P.S.; Vassileva, V.; Pinto, G. Deciphering the epigenetic alphabet involved in transgenerational stress memory in crops. Int. J. Mol. Sci. 2021, 22, 7118. [Google Scholar] [CrossRef]
- Bhar, A.; Chatterjee, M.; Gupta, S.; Das, S. Salicylic acid regulates systemic defense signaling in chickpea during Fusarium oxysporum f. sp. ciceri race 1 infection. Plant Mol. Biol. Rep. 2018, 36, 162–175. [Google Scholar] [CrossRef]
- Gupta, S.; Bhar, A.; Chatterjee, M.; Das, S. Fusarium oxysporum f. sp. ciceri race 1 induced redox state alterations are coupled to downstream defense signaling in root tissues of chickpea (Cicer arietinum L.). PLoS ONE 2013, 8, e73163. [Google Scholar] [CrossRef] [Green Version]
- Zander, M.; Lewsey, M.G.; Clark, N.M.; Yin, L.; Bartlett, A.; Guzmán, J.P.S.; Hann, E.; Langford, A.E.; Jow, B.; Wise, A. Integrated multi-omics framework of the plant response to jasmonic acid. Nat. Plants 2020, 6, 290–302. [Google Scholar] [CrossRef]
- Aerts, N.; Pereira Mendes, M.; Van Wees, S.C. Multiple levels of crosstalk in hormone networks regulating plant defense. Plant J. 2021, 105, 489–504. [Google Scholar] [CrossRef] [PubMed]
- Tuteja, N.; Mahajan, S. Calcium signaling network in plants: An overview. Plant Signal. Behav. 2007, 2, 79–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, J.P.; Sweatt, J.D. Molecular psychology: Roles for the ERK MAP kinase cascade in memory. Annu. Rev. Pharmacol. Toxicol. 2002, 42, 135–163. [Google Scholar] [CrossRef]
- Rasmussen, M.W.; Roux, M.; Petersen, M.; Mundy, J. MAP kinase cascades in Arabidopsis innate immunity. Front. Plant Sci. 2012, 3, 169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, J.D.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [Green Version]
- Bigeard, J.; Colcombet, J.; Hirt, H. Signaling mechanisms in pattern-triggered immunity (PTI). Mol. Plant 2015, 8, 521–539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhar, A.; Gupta, S.; Chatterjee, M.; Das, S. Redox Regulatory Networks in Response to Biotic Stress in Plants: A New Insight Through Chickpea-Fusarium Interplay. In Mechanism of Plant Hormone Signaling under Stress; Pandey, G.K., Ed.; John Wiley & Sons: Hoboken, NJ, USA, 2017; Volume 2, pp. 23–43. [Google Scholar]
- Huang, H.; Ullah, F.; Zhou, D.-X.; Yi, M.; Zhao, Y. Mechanisms of ROS regulation of plant development and stress responses. Front. Plant Sci. 2019, 10, 800. [Google Scholar] [CrossRef]
- Tsuge, T.; Harimoto, Y.; Akimitsu, K.; Ohtani, K.; Kodama, M.; Akagi, Y.; Egusa, M.; Yamamoto, M.; Otani, H. Host-selective toxins produced by the plant pathogenic fungus Alternaria alternata. FEMS Microbiol. Rev. 2013, 37, 44–66. [Google Scholar] [CrossRef]
- Bacon, C.; Porter, J.; Norred, W.; Leslie, J. Production of fusaric acid by Fusarium species. Appl. Environ. Microbiol. 1996, 62, 4039–4043. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Zuo, C.; Deng, G.; Kuang, R.; Yang, Q.; Hu, C.; Sheng, O.; Zhang, S.; Ma, L.; Wei, Y. Contamination of bananas with beauvericin and fusaric acid produced by Fusarium oxysporum f. sp. cubense. PLoS ONE 2013, 8, e70226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomma, B.P.; Nürnberger, T.; Joosten, M.H. Of PAMPs and effectors: The blurred PTI-ETI dichotomy. Plant Cell 2011, 23, 4–15. [Google Scholar] [CrossRef] [Green Version]
- Kaur, B.; Bhatia, D.; Mavi, G. Eighty years of gene-for-gene relationship and its applications in identification and utilization of R genes. J. Genet. 2021, 100, 50. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Sun, P.; Yuan, J.; Gong, K.; Li, N.; Meng, F.; Zhang, Z.; Li, X.; Hu, J.; Wang, J. The celery genome sequence reveals sequential paleo-polyploidizations, karyotype evolution and resistance gene reduction in apiales. Plant Biotechnol. J. 2021, 19, 731–744. [Google Scholar] [CrossRef]
- Kahlon, P.S.; Stam, R. Polymorphisms in plants to restrict losses to pathogens: From gene family expansions to complex network evolution. Curr. Opin. Plant Biol. 2021, 62, 102040. [Google Scholar] [CrossRef] [PubMed]
- Kenrick, P.; Crane, P.R. The origin and early evolution of plants on land. Nature 1997, 389, 33–39. [Google Scholar] [CrossRef]
- Fürst-Jansen, J.M.; de Vries, S.; de Vries, J. Evo-physio: On stress responses and the earliest land plants. J. Exp. Bot. 2020, 71, 3254–3269. [Google Scholar] [CrossRef] [Green Version]
- Lyu, D.; Msimbira, L.A.; Nazari, M.; Antar, M.; Pagé, A.; Shah, A.; Monjezi, N.; Zajonc, J.; Tanney, C.A.; Backer, R. The Coevolution of Plants and Microbes Underpins Sustainable Agriculture. Microorganisms 2021, 9, 1036. [Google Scholar] [CrossRef]
- Song, W.-Y.; Wang, G.-L.; Chen, L.-L.; Kim, H.-S.; Pi, L.-Y.; Holsten, T.; Gardner, J.; Wang, B.; Zhai, W.-X.; Zhu, L.-H. A receptor kinase-like protein encoded by the rice disease resistance gene, Xa21. Science 1995, 270, 1804–1806. [Google Scholar] [CrossRef] [Green Version]
- Mubassir, M.; Naser, M.A.; Abdul-Wahab, M.F.; Jawad, T.; Alvy, R.I.; Hamdan, S. Comprehensive in silico modeling of the rice plant PRR Xa21 and its interaction with RaxX21-sY and OsSERK2. RSC Adv. 2020, 10, 15800–15814. [Google Scholar] [CrossRef]
- Peng, H.; Chen, Z.; Fang, Z.; Zhou, J.; Xia, Z.; Gao, L.; Chen, L.; Li, L.; Li, T.; Zhai, W. Rice Xa21 primed genes and pathways that are critical for combating bacterial blight infection. Sci. Rep. 2015, 5, 12165. [Google Scholar] [CrossRef] [Green Version]
- Chinchilla, D.; Zipfel, C.; Robatzek, S.; Kemmerling, B.; Nürnberger, T.; Jones, J.D.; Felix, G.; Boller, T. A flagellin-induced complex of the receptor FLS2 and BAK1 initiates plant defence. Nature 2007, 448, 497–500. [Google Scholar] [CrossRef]
- Koc, A.; Markovic, D.; Ninkovic, V.; Martinez, G. Molecular mechanisms regulating priming and stress memory. In Priming-Mediated Stress and Cross-Stress Tolerance in Crop Plants; Elsevier: Amsterdam, The Netherlands, 2020; pp. 247–265. [Google Scholar]
- Beckers, G.J.; Jaskiewicz, M.; Liu, Y.; Underwood, W.R.; He, S.Y.; Zhang, S.; Conrath, U. Mitogen-activated protein kinases 3 and 6 are required for full priming of stress responses in Arabidopsis thaliana. Plant Cell 2009, 21, 944–953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tateda, C.; Zhang, Z.; Shrestha, J.; Jelenska, J.; Chinchilla, D.; Greenberg, J.T. Salicylic acid regulates Arabidopsis microbial pattern receptor kinase levels and signaling. Plant Cell 2014, 26, 4171–4187. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Liu, N.; Gao, C.; Cai, H.; Romeis, T.; Tang, D. The Arabidopsis exocyst subunits EXO70B1 and EXO70B2 regulate FLS2 homeostasis at the plasma membrane. New Phytol. 2020, 227, 529–544. [Google Scholar] [CrossRef]
- Huang, P.Y.; Yeh, Y.H.; Liu, A.C.; Cheng, C.P.; Zimmerli, L. The Arabidopsis LecRK-VI. 2 associates with the pattern-recognition receptor FLS 2 and primes Nicotiana benthamiana pattern-triggered immunity. Plant J. 2014, 79, 243–255. [Google Scholar] [CrossRef] [PubMed]
- Giovannoni, M.; Marti, L.; Ferrari, S.; Tanaka-Takada, N.; Maeshima, M.; Ott, T.; De Lorenzo, G.; Mattei, B. The plasma membrane-associated Ca2+-binding protein PCaP1 is required for oligogalacturonide and flagellin-induced priming and immunity. Plant Cell Environ. 2021, 44, 3775–3792. [Google Scholar] [CrossRef]
- Lolle, S.; Stevens, D.; Coaker, G. Plant NLR-triggered immunity: From receptor activation to downstream signaling. Curr. Opin. Immunol. 2020, 62, 99–105. [Google Scholar] [CrossRef]
- Adachi, H.; Kamoun, S.; Maqbool, A. A resistosome-activated ‘death switch’. Nat. Plants 2019, 5, 457–458. [Google Scholar] [CrossRef] [PubMed]
- Banani, S.F.; Lee, H.O.; Hyman, A.A.; Rosen, M.K. Biomolecular condensates: Organizers of cellular biochemistry. Nat. Rev. Mol. Cell Biol. 2017, 18, 285–298. [Google Scholar] [CrossRef]
- Wang, W.; Gu, Y. The emerging role of biomolecular condensates in plant immunity. Plant Cell 2021, koab240. [Google Scholar] [CrossRef]
- Cutter, A.R.; Hayes, J.J. A brief review of nucleosome structure. FEBS Lett. 2015, 589, 2914–2922. [Google Scholar] [CrossRef] [Green Version]
- Bräutigam, K.; Cronk, Q. DNA methylation and the evolution of developmental complexity in plants. Front. Plant Sci. 2018, 9, 1447. [Google Scholar] [CrossRef] [Green Version]
- Baulcombe, D.C.; Dean, C. Epigenetic regulation in plant responses to the environment. Cold Spring Harb. Perspect. Biol. 2014, 6, a019471. [Google Scholar] [CrossRef]
- Duempelmann, L.; Skribbe, M.; Bühler, M. Small RNAs in the transgenerational inheritance of epigenetic information. Trends Genet. 2020, 36, 203–214. [Google Scholar] [CrossRef] [Green Version]
- Stassen, J.H.; López, A.; Jain, R.; Pascual-Pardo, D.; Luna, E.; Smith, L.M.; Ton, J. The relationship between transgenerational acquired resistance and global DNA methylation in Arabidopsis. Sci. Rep. 2018, 8, 14761. [Google Scholar] [CrossRef] [Green Version]
- Vivas, M.; Hernández, J.; Corcobado, T.; Cubera, E.; Solla, A. Transgenerational Induction of Resistance to Phytophthora cinnamomi in Holm Oak. Forests 2021, 12, 100. [Google Scholar] [CrossRef]
- Lämke, J.; Brzezinka, K.; Bäurle, I. HSFA2 orchestrates transcriptional dynamics after heat stress in Arabidopsis thaliana. Transcription 2016, 7, 111–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabrizio, P.; Garvis, S.; Palladino, F. Histone methylation and memory of environmental stress. Cells 2019, 8, 339. [Google Scholar] [CrossRef] [Green Version]
- Khan, A.R.; Enjalbert, J.; Marsollier, A.-C.; Rousselet, A.; Goldringer, I.; Vitte, C. Vernalization treatment induces site-specific DNA hypermethylation at the VERNALIZATION-A1 (VRN-A1) locus in hexaploid winter wheat. BMC Plant Biol. 2013, 13, 209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noh, S.W.; Seo, R.-R.; Park, H.J.; Jung, H.W. Two Arabidopsis homologs of human lysine-specific demethylase function in epigenetic regulation of plant defense responses. Front. Plant Sci. 2021, 12, 688003. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-H. Multifaceted Chromatin Structure and Transcription Changes in Plant Stress Response. Int. J. Mol. Sci. 2021, 22, 2013. [Google Scholar] [CrossRef]
- Amaral, J.; Lamelas, L.; Valledor, L.; Castillejo, M.Á.; Alves, A.; Pinto, G. Comparative proteomics of Pinus–Fusarium circinatum interactions reveal metabolic clues to biotic stress resistance. Physiol. Plant. 2021, 173, 2142–2154. [Google Scholar] [CrossRef] [PubMed]
- Ando, S.; Jaskiewicz, M.; Mochizuki, S.; Koseki, S.; Miyashita, S.; Takahashi, H.; Conrath, U. Priming for enhanced ARGONAUTE2 activation accompanies induced resistance to cucumber mosaic virus in Arabidopsis thaliana. Mol. Plant Pathol. 2021, 22, 19–30. [Google Scholar] [CrossRef]
- Jaskiewicz, M.; Conrath, U.; Peterhänsel, C. Chromatin modification acts as a memory for systemic acquired resistance in the plant stress response. EMBO Rep. 2011, 12, 50–55. [Google Scholar] [CrossRef] [Green Version]
- Lusser, A.; Kölle, D.; Loidl, P. Histone acetylation: Lessons from the plant kingdom. Trends Plant Sci. 2001, 6, 59–65. [Google Scholar] [CrossRef]
- Hartl, M.; Füßl, M.; Boersema, P.J.; Jost, J.O.; Kramer, K.; Bakirbas, A.; Sindlinger, J.; Plöchinger, M.; Leister, D.; Uhrig, G. Lysine acetylome profiling uncovers novel histone deacetylase substrate proteins in Arabidopsis. Mol. Syst. Biol. 2017, 13, 949. [Google Scholar] [CrossRef]
- Zhou, C.; Zhang, L.; Duan, J.; Miki, B.; Wu, K. HISTONE DEACETYLASE19 is involved in jasmonic acid and ethylene signaling of pathogen response in Arabidopsis. Plant Cell 2005, 17, 1196–1204. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.M.; Song, H.R.; Han, S.K.; Han, M.; Kim, C.Y.; Park, J.; Lee, Y.H.; Jeon, J.S.; Noh, Y.S.; Noh, B. HDA19 is required for the repression of salicylic acid biosynthesis and salicylic acid-mediated defense responses in Arabidopsis. Plant J. 2012, 71, 135–146. [Google Scholar] [CrossRef]
- Backer, R.; Naidoo, S.; Van den Berg, N. The NONEXPRESSOR OF PATHOGENESIS-RELATED GENES 1 (NPR1) and related family: Mechanistic insights in plant disease resistance. Front. Plant Sci. 2019, 10, 102. [Google Scholar] [CrossRef] [Green Version]
- Yildiz, I.; Mantz, M.; Hartmann, M.; Zeier, T.; Kessel, J.; Thurow, C.; Gatz, C.; Petzsch, P.; Köhrer, K.; Zeier, J. The mobile SAR signal N-hydroxypipecolic acid induces NPR1-dependent transcriptional reprogramming and immune priming. Plant Physiol. 2021, 186, 1679–1705. [Google Scholar] [CrossRef]
- Wu, K.; Zhang, L.; Zhou, C.; Yu, C.-W.; Chaikam, V. HDA6 is required for jasmonate response, senescence and flowering in Arabidopsis. J. Exp. Bot. 2008, 59, 225–234. [Google Scholar] [CrossRef]
- Ramirez-Prado, J.S.; Piquerez, S.J.; Bendahmane, A.; Hirt, H.; Raynaud, C.; Benhamed, M. Modify the histone to win the battle: Chromatin dynamics in plant–pathogen interactions. Front. Plant Sci. 2018, 9, 355. [Google Scholar] [CrossRef]
- Singh, V.; Banday, Z.Z.; Nandi, A.K. Exogenous application of histone demethylase inhibitor trans-2-phenylcyclopropylamine mimics FLD loss-of-function phenotype in terms of systemic acquired resistance in Arabidopsis thaliana. Plant Signal. Behav. 2014, 9, e29658. [Google Scholar] [CrossRef] [Green Version]
- Gully, K.; Celton, J.-M.; Degrave, A.; Pelletier, S.; Brisset, M.-N.; Bucher, E. Biotic stress-induced priming and de-priming of transcriptional memory in Arabidopsis and apple. Epigenomes 2019, 3, 3. [Google Scholar] [CrossRef] [Green Version]
- Yuan, J.; Wang, S.; Liu, Y.-H.; Li, J.-X.; Zhou, L.-Y.; Li, T.; Zhou, L.; Zhang, W.-J.; Guo, L.-P.; Huang, L.-Q. Preliminary study on memory function of Sorbus aucuparia suspension cell to biotic stress. Zhongguo Zhong Yao Za Zhi=Zhongguo Zhongyao Zazhi=China J. Chin. Mater. Med. 2021, 46, 2467–2473. [Google Scholar]
- Abulfaraj, A.A. Investigating the Role of the Arabidopsis Homologue of the Human G3BP in RNA Metabolism, Cellular Stress Responses and Innate Immunity. Ph.D. Thesis, King Abdullah University of Science and Technology, Thuwal, Saudi Arabia, 2018. [Google Scholar]
- Zhao, K.; Kong, D.; Jin, B.; Smolke, C.D.; Rhee, S.Y. A novel bivalent chromatin associates with rapid induction of camalexin biosynthesis genes in response to a pathogen signal in Arabidopsis. Elife 2021, 10, e69508. [Google Scholar] [CrossRef] [PubMed]
- Werghi, S.; Herrero, F.A.; Fakhfakh, H.; Gorsane, F. Auxin drives tomato spotted wilt virus (TSWV) resistance through epigenetic regulation of auxin response factor ARF8 expression in tomato. Gene 2021, 804, 145905. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Ménard, R.; Li, Y.; Coruzzi, G.M.; Heitz, T.; Shen, W.-H.; Berr, A. Arabidopsis SDG8 potentiates the sustainable transcriptional induction of the pathogenesis-related genes PR1 and PR2 during plant defense response. Front. Plant Sci. 2020, 11, 277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramírez-Tejero, J.A.; Cabanás, C.G.-L.; Valverde-Corredor, A.; Mercado-Blanco, J.; Luque, F. Epigenetic Regulation of Verticillium dahliae Virulence: Does DNA Methylation Level Play A Role? Int. J. Mol. Sci. 2020, 21, 5197. [Google Scholar] [CrossRef]
- Chen, J.; Clinton, M.; Qi, G.; Wang, D.; Liu, F.; Fu, Z.Q. Reprogramming and remodeling: Transcriptional and epigenetic regulation of salicylic acid-mediated plant defense. J. Exp. Bot. 2020, 71, 5256–5268. [Google Scholar] [CrossRef] [PubMed]
- Crespo-Salvador, Ó.; Sánchez-Giménez, L.; López-Galiano, M.; Fernández-Crespo, E.; Scalschi, L.; García-Robles, I.; Rausell, C.; Real, M.D.; González-Bosch, C. The histone marks signature in exonic and intronic regions is relevant in early response of tomato genes to Botrytis cinerea and in miRNA regulation. Plants 2020, 9, 300. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Chen, D.; Sun, F.; Guo, W.; Wang, W.; Li, X.; Lan, Y.; Du, L.; Li, S.; Fan, Y. ARGONAUTE 2 increases rice susceptibility to rice black-streaked dwarf virus infection by epigenetically regulating HEXOKINASE 1 expression. Mol. Plant Pathol. 2021, 22, 1029–1040. [Google Scholar] [CrossRef]
- Lodhi, N.; Singh, M.; Srivastava, R.; Sawant, S.V.; Tuli, R. Epigenetic Malleability at Core Promoter Regulates Tobacco PR-1a Expression after Salicylic Acid Treatment. bioRxiv 2021. [Google Scholar] [CrossRef]
- Martínez-Aguilar, K.; Hernández-Chávez, J.L.; Alvarez-Venegas, R. Priming of seeds with INA and its transgenerational effect in common bean (Phaseolus vulgaris L.) plants. Plant Sci. 2021, 305, 110834. [Google Scholar] [CrossRef]
- Da Silva, A.R.; da Costa Silva, D.; dos Santos Pinto, K.N.; Santos Filho, H.P.; Coelho Filho, M.A.; dos Santos Soares Filho, W.; Ferreira, C.F.; da Silva Gesteira, A. Epigenetic responses to Phytophthora citrophthora gummosis in citrus. Plant Sci. 2021, 313, 111082. [Google Scholar] [CrossRef]
- Chacón-Cerdas, R.; Barboza-Barquero, L.; Albertazzi, F.J.; Rivera-Mendez, W. Transcription factors controlling biotic stress response in potato plants. Physiol. Mol. Plant Pathol. 2020, 112, 101527. [Google Scholar] [CrossRef]
- Burns, A.M.; Gräff, J. Cognitive epigenetic priming: Leveraging histone acetylation for memory amelioration. Curr. Opin. Neurobiol. 2021, 67, 75–84. [Google Scholar] [CrossRef]
- Bhatt, D.; Saxena, S.C.; Arora, S. ROS Signaling Under Oxidative Stress in Plants. In Microbes and Signaling Biomolecules against Plant Stress; Springer: Berlin/Heidelberg, Germany, 2021; pp. 269–286. [Google Scholar]
- Park, S.; Waterland, N.L. Evaluation of Calcium Application Methods on Delaying Plant Wilting under Water Deficit in Bedding Plants. Agronomy 2021, 11, 1383. [Google Scholar] [CrossRef]
- Singhal, R.K.; Jatav, H.S.; Aftab, T.; Pandey, S.; Mishra, U.N.; Chauhan, J.; Chand, S.; Saha, D.; Dadarwal, B.K.; Chandra, K. Roles of nitric oxide in conferring multiple abiotic stress tolerance in plants and crosstalk with other plant growth regulators. J. Plant Growth Regul. 2021, 40, 2303–2328. [Google Scholar] [CrossRef]
- Singh, P.; Arif, Y.; Siddiqui, H.; Hayat, S. Jasmonate: A Versatile Messenger in Plants. In Jasmonates and Salicylates Signaling in Plants; Springer: Berlin/Heidelberg, Germany, 2021; pp. 129–158. [Google Scholar]
- Bhadouriya, S.L.; Mehrotra, S.; Basantani, M.K.; Loake, G.J.; Mehrotra, R. Role of chromatin architecture in plant stress responses: An update. Front. Plant Sci. 2021, 11, 2131. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Plants | Pathogen | Effector | Epigenetic Control | Pathway Induced | Reference |
|---|---|---|---|---|---|
| Arabidopsis and Apple | - | SA analogue | DNA methylation | - | Gully et al., 2019 [70] |
| Sorbus aucuparia suspension cell (SASC) | - | Yeast extract | - | Secondary metabolite induction | Yuan et al., 2021 [71] |
| Arabidopsis | - | MAMP | - | MAP kinase cascade, G3BP | Abulfaraj, 2018 [72] |
| Arabidopsis | - | - | H3K27me3 (repression) and H3K18Ac (activation) | Induction of the camalexin pathway | Zhao et al., 2021 [73] |
| Arabidopsis | Pseudomonas syringae | - | H3K4 | LDL1 and LDL2-mediated pathway | Noh et al., 2021 [54] |
| Tomato (Solanum lycopersicum) | TSWV | - | Methylation in cytosine residue of ARF8 and miRNA167a | Auxin-mediated pathway | Werghi et al., 2021 [74] |
| Arabidopsis | Pseudomonas syringae | - | SDG8 mediated methylation at H3K36me3 | PR1 and PR2-mediated parhway | Zhang et al., 2020 [75] |
| Olive (Olea sp.) | Verticillium dahliae | - | Methylation of 831 gene | - | Ramírez-Tejero et al., 2020 [76] |
| Tomato (Solanum lycopersicum) | Pseudomonas syringae pv. tomato DC3000 | - | SDG8 and SDG25-induced methylation in receptor CER3 locus (H3K4 and H3K36) and in the promoter region of PR1 (H3Ac, H4Ac and H3K4me3) | SAR pathway | Chen et al., 2020 [77] |
| Tomato (Solanum lycopersicum) | Botrytis cinerea | - | H3K9Ac of SlyDES, SlyDOX1 and SlyLoxD | Oxylipin pathway | Crespo-Salvador et al., 2020 [78] |
| Rice (Oryza sativa) | Black Streaked Dwarf Virus (RBSDV) | - | Argonoute (OsAGO2) methylates and suppresses hexokinase (OsHXK1) | ROS-mediated pathway | Wang et al., 2021 [79] |
| Tobacco (Nicotiana tabacum) | - | SA | Epigenetic modification at H3K9, H4K20 and H4K16 of PR1a gene | SAR pathway | Lodhi et al., 2021 [80] |
| Common bean (Phaseolus vulgaris) | Pseudomonas syringae pv. phaseolicola | INA (2,6 dichloro isonicotinic acid) | Epigenetic modification of H3K4me3 and H3K36me3 of PvPR1 gene promoter | SAR pathway | Martínez-Aguilar et al., 2021 [81] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bhar, A.; Chakraborty, A.; Roy, A. Plant Responses to Biotic Stress: Old Memories Matter. Plants 2022, 11, 84. https://doi.org/10.3390/plants11010084
Bhar A, Chakraborty A, Roy A. Plant Responses to Biotic Stress: Old Memories Matter. Plants. 2022; 11(1):84. https://doi.org/10.3390/plants11010084
Chicago/Turabian StyleBhar, Anirban, Amrita Chakraborty, and Amit Roy. 2022. "Plant Responses to Biotic Stress: Old Memories Matter" Plants 11, no. 1: 84. https://doi.org/10.3390/plants11010084