
The Omics Hunt for Novel Molecular Markers of Resistance to Phytophthora infestans

 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
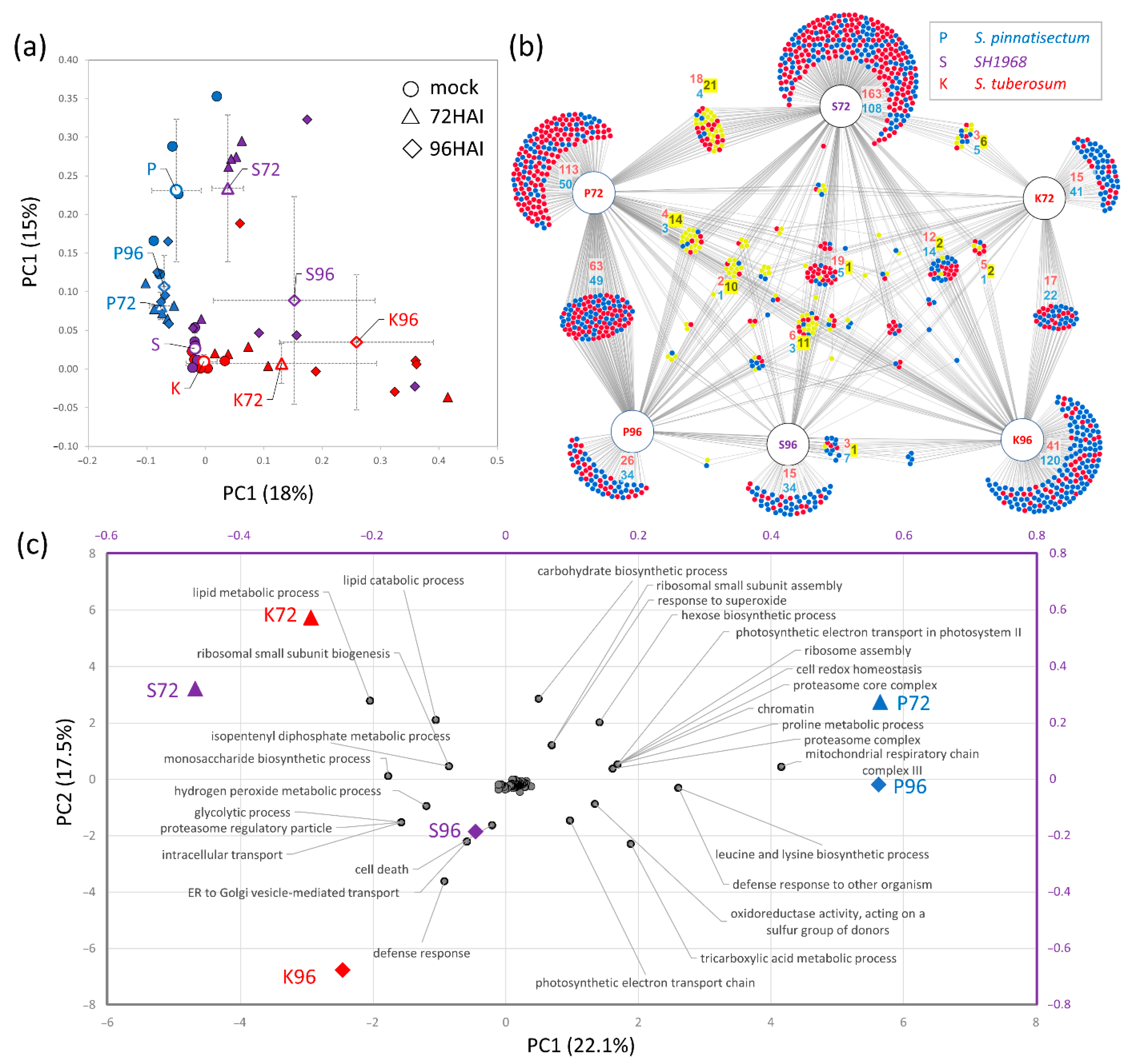
2.1. The Composition of Leaf Proteome in Response to Inoculation
2.2. Functional Analysis of Differentially Abundant Proteins
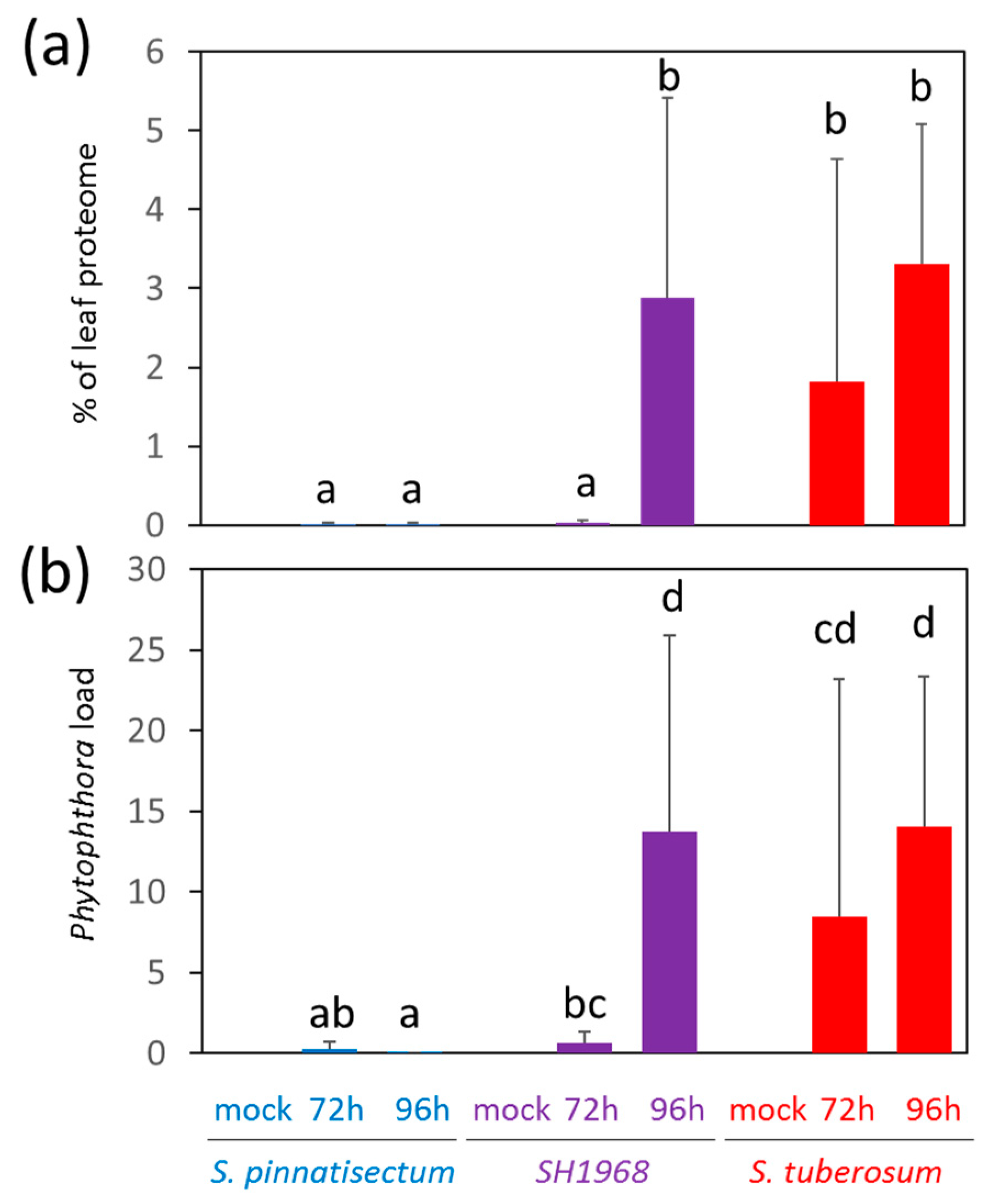
2.3. Peptide-Based Detection of P. infestans
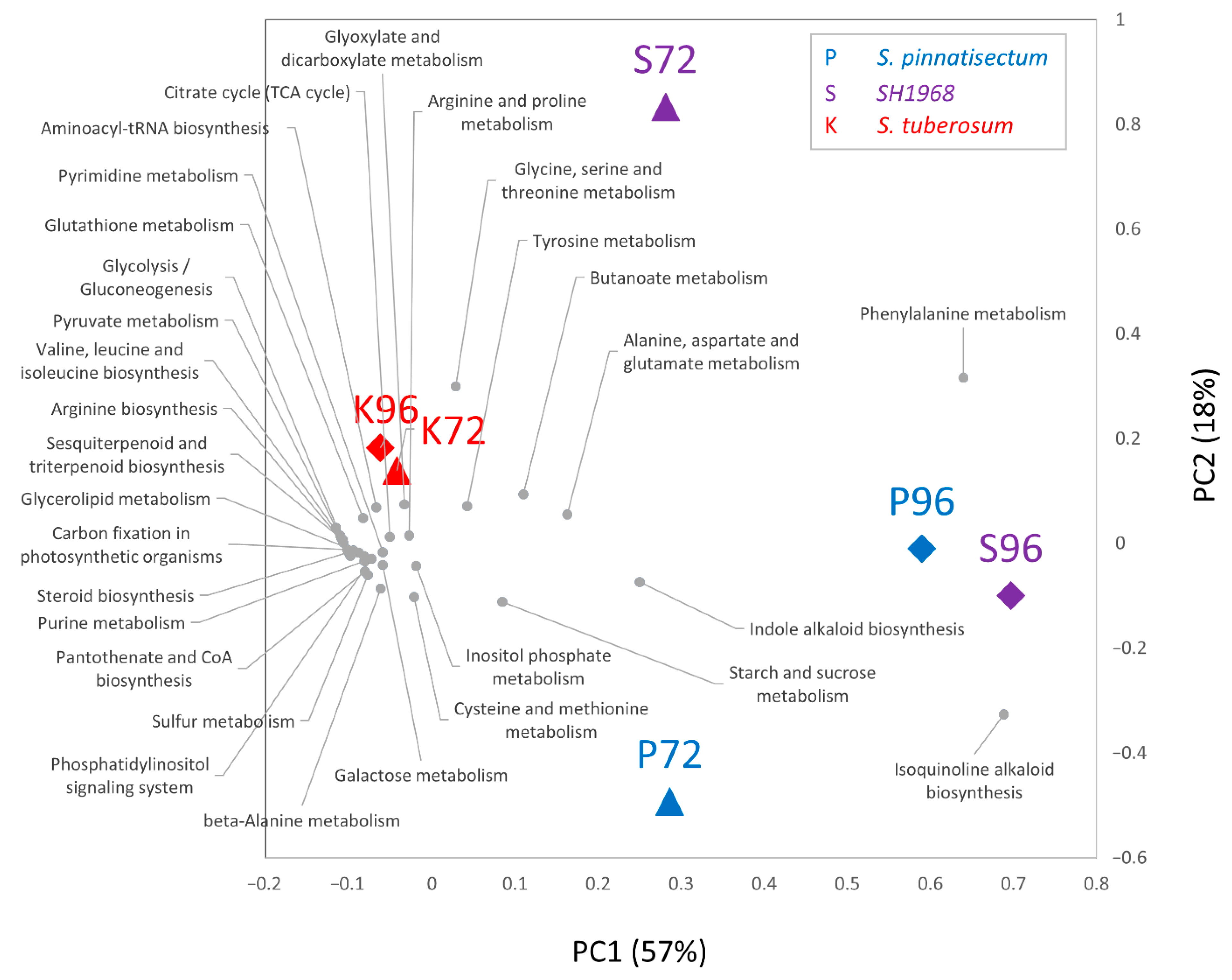
2.4. Metabolomics Revealed Modulation in Both Primary Metabolism and Production of Secondary Metabolites
2.5. Lipidomics Analysis Confirmed the Impact of P. infestans Inoculation on Leaf Lipidome
2.6. Identification of Compounds with a Putative Role in Resistance against P. infestans
2.7. Targeted Analysis of Putative Protein Resistance Markers in Genotypes with Contrasting Phytophthora Resistance
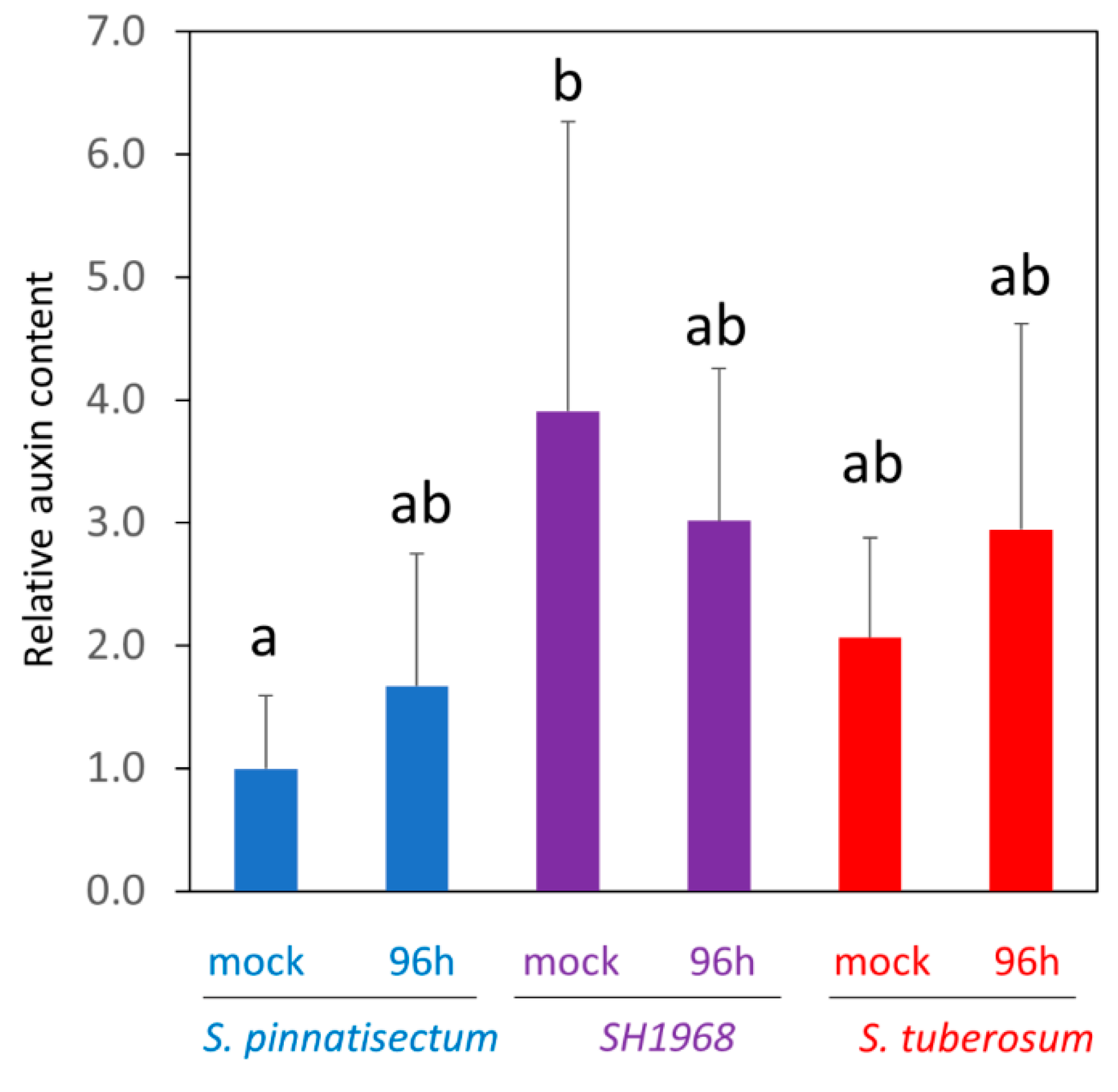
2.8. Determination of Auxin Content in Response to Inoculation
3. Discussion
3.1. The Genotype-Specific Resistance Gradient Was Attenuated but Preserved in the Detached Leaf Experiment
3.2. The Resistant Genotype Accumulated Proline, but the Abundance of the Corresponding Biosynthetic Enzyme Did Not Correlate with the Resistance
3.3. Uricase Activity in S. pinnatisectum May Promote the Resistance to Phytophthora
3.4. The Abundance of Auxin Metabolism Enzyme Was Positively Correlated with Resistance
3.5. Heat Shock Protein 70 Has a Putative Role in Phytophthora Resistance
3.6. The Resistant Genotype Accumulated Higher Amounts of Phenolics and Cysteamine
4. Materials and Methods
4.1. Plant Material, Cultivation, and Inoculation
4.2. Detection of P. infestans by qPCR
4.3. Proteome Analysis
4.4. Metabolomic Analysis
4.5. Lipidome Analysis
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Savary, S.; Willocquet, L.; Pethybridge, S.J.; Esker, P.; McRoberts, N.; Nelson, A. The global burden of pathogens and pests on major food crops. Nat. Ecol. Evol. 2019, 3, 430–439. [Google Scholar] [CrossRef] [PubMed]
- Fry, W.E.; Birch, P.R.J.; Judelson, H.S.; Grünwald, N.J.; Danies, G.; Everts, K.L.; Gevens, A.J.; Gugino, B.K.; Johnson, D.A.; Johnson, S.B.; et al. Five reasons to consider Phytophthora infestans a reemerging pathogen. Phytopathology 2015, 105, 966–981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leesutthiphonchai, W.; Vu, A.L.; Ah-Fong, A.M.V.; Judelson, H.S. How does Phytophthora infestans evade control efforts? Modern insight into the late blight disease. Phytopathology 2018, 108, 916–924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nowicki, M.; Foolad, M.R.; Nowakowska, M.; Kozik, E.U. Potato and tomato late blight caused by Phytophthora infestans: An overview of pathology and resistance breeding. Plant Dis. 2012, 96, 4–17. [Google Scholar] [CrossRef] [Green Version]
- Machida-Hirano, R. Diversity of potato genetic resources. Breed. Sci. 2015, 65, 26–40. [Google Scholar] [CrossRef] [Green Version]
- Karki, H.S.; Jansky, S.H.; Halterman, D.A. Screening of wild potatoes identifies new sources of late blight resistance. Plant Dis. 2021, 105, 368–376. [Google Scholar] [CrossRef]
- Vetukuri, R.R.; Masini, L.; McDougal, R.; Panda, P.; de Zinger, L.; Brus-Szkalej, M.; Lankinen, Å.; Grenville-Briggs, L.J. The presence of Phytophthora infestans in the rhizosphere of a wild Solanum species may contribute to off-season survival and pathogenicity. Appl. Soil Ecol. 2020, 148, 103475. [Google Scholar] [CrossRef]
- Van Weymers, P.S.M.; Baker, K.; Chen, X.; Harrower, B.; Cooke, D.E.L.; Gilroy, E.M.; Birch, P.R.J.; Thilliez, G.J.A.; Lees, A.K.; Lynott, J.S.; et al. Utilizing “Omic” technologies to identify and prioritize novel sources of resistance to the oomycete pathogen Phytophthora infestans in potato germplasm collections. Front. Plant Sci. 2016, 7, 672. [Google Scholar] [CrossRef] [Green Version]
- Zheng, J.; Duan, S.; Armstrong, M.R.; Duan, Y.; Xu, J.; Chen, X.; Hein, I.; Jin, L.; Li, G. New findings on the resistance mechanism of an elite diploid wild potato species JAM1-4 in response to a super race strain of Phytophthora infestans. Phytopathology 2020, 110, 1375–1387. [Google Scholar] [CrossRef]
- Gu, B.; Cao, X.; Zhou, X.; Chen, Z.; Wang, Q.; Liu, W.; Chen, Q.; Zhao, H. The histological, effectoromic, and transcriptomic analyses of Solanum pinnatisectum reveal an upregulation of multiple NBS-LRR genes suppressing Phytophthora infestans Infection. Int. J. Mol. Sci. 2020, 21, 3211. [Google Scholar] [CrossRef]
- Pérez, W.; Ñahui, M.; Ellis, D.; Forbes, G.A. Wide phenotypic diversity for resistance to Phytophthora infestans found in potato landraces from Peru. Plant Dis. 2014, 98, 1530–1533. [Google Scholar] [CrossRef] [Green Version]
- Bachmann-Pfabe, S.; Hammann, T.; Kruse, J.; Dehmer, K.J. Screening of wild potato genetic resources for combined resistance to late blight on tubers and pale potato cyst nematodes. Euphytica 2019, 215, 48. [Google Scholar] [CrossRef]
- Akino, S.; Takemoto, D.; Hosaka, K. Phytophthora infestans: A review of past and current studies on potato late blight. J. Gen. Plant Pathol. 2014, 80, 24–37. [Google Scholar] [CrossRef]
- Pathania, A.; Rialch, N.; Sharma, P.N. Marker-assisted selection in disease resistance breeding. In Current Developments in Biotechnology and Bioengineering; Elsevier: Amsterdam, The Netherlands, 2017; pp. 187–213. [Google Scholar]
- Ramakrishnan, A.P.; Ritland, C.E.; Blas Sevillano, R.H.; Riseman, A. Review of potato molecular markers to enhance trait selection. Am. J. Potato Res. 2015, 92, 455–472. [Google Scholar] [CrossRef]
- Slater, A.T.; Cogan, N.O.I.; Forster, J.W. Cost analysis of the application of marker-assisted selection in potato breeding. Mol. Breed. 2013, 32, 299–310. [Google Scholar] [CrossRef]
- Ortega, F.; Lopez-Vizcon, C. Application of molecular marker-assisted selection (MAS) for disease resistance in a practical potato breeding programme. Potato Res. 2012, 55, 1–13. [Google Scholar] [CrossRef]
- Lamichhane, J.R.; Osdaghi, E.; Behlau, F.; Köhl, J.; Jones, J.B.; Aubertot, J.-N. Thirteen decades of antimicrobial copper compounds applied in agriculture. A review. Agron. Sustain. Dev. 2018, 38, 28. [Google Scholar] [CrossRef] [Green Version]
- Najdabbasi, N.; Mirmajlessi, S.M.; Dewitte, K.; Ameye, M.; Mänd, M.; Audenaert, K.; Landschoot, S.; Haesaert, G. Green leaf volatile confers management of late blight disease: A green vaccination in potato. J. Fungi 2021, 7, 312. [Google Scholar] [CrossRef]
- Elsherbiny, E.A.; Amin, B.H.; Aleem, B.; Kingsley, K.L.; Bennett, J.W. Trichoderma volatile organic compounds as a biofumigation tool against late blight pathogen Phytophthora infestans in postharvest potato tubers. J. Agric. Food Chem. 2020, 68, 8163–8171. [Google Scholar] [CrossRef]
- Yao, Y.; Li, Y.; Chen, Z.; Zheng, B.; Zhang, L.; Niu, B.; Meng, J.; Li, A.; Zhang, J.; Wang, Q. Biological control of potato late blight using isolates of Trichoderma. Am. J. Potato Res. 2016, 93, 33–42. [Google Scholar] [CrossRef]
- Hadwiger, L.A.; McDonel, H.; Glawe, D. Wild yeast strains as prospective candidates to induce resistance against potato late blight (Phytophthora infestans). Am. J. Potato Res. 2015, 92, 379–386. [Google Scholar] [CrossRef]
- De Vrieze, M.; Pandey, P.; Bucheli, T.D.; Varadarajan, A.R.; Ahrens, C.H.; Weisskopf, L.; Bailly, A. Volatile organic compounds from native potato-associated Pseudomonas as potential anti-oomycete agents. Front. Microbiol. 2015, 6, 1295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiwari, J.K.; Rawat, S.; Luthra, S.K.; Zinta, R.; Sahu, S.; Varshney, S.; Kumar, V.; Dalamu, D.; Mandadi, N.; Kumar, M.; et al. Genome sequence analysis provides insights on genomic variation and late blight resistance genes in potato somatic hybrid (parents and progeny). Mol. Biol. Rep. 2021, 48, 623–635. [Google Scholar] [CrossRef] [PubMed]
- Tian, T.; Liu, Y.; Yan, H.; You, Q.; Yi, X.; Du, Z.; Xu, W.; Su, Z. agriGO v2.0: A GO analysis toolkit for the agricultural community, 2017 update. Nucleic Acids Res. 2017, 45, W122–W129. [Google Scholar] [CrossRef]
- Sun, L.; Dong, S.; Ge, Y.; Fonseca, J.P.; Robinson, Z.T.; Mysore, K.S.; Mehta, P. DiVenn: An interactive and integrated web-based visualization tool for comparing gene lists. Front. Genet. 2019, 10, 421. [Google Scholar] [CrossRef]
- Berka, M.; Greplová, M.; Saiz-Fernández, I.; Novák, J.; Luklová, M.; Zelená, P.; Tomšovský, M.; Brzobohatý, B.; Černý, M. Peptide-based identification of Phytophthora isolates and Phytophthora detection in planta. Int. J. Mol. Sci. 2020, 21, 9463. [Google Scholar] [CrossRef]
- Pang, Z.; Chong, J.; Zhou, G.; De Lima Morais, D.A.; Chang, L.; Barrette, M.; Gauthier, C.; Jacques, P.É.; Li, S.; Xia, J. MetaboAnalyst 5.0: Narrowing the gap between raw spectra and functional insights. Nucleic Acids Res. 2021, 49, W388–W396. [Google Scholar] [CrossRef]
- Von Saint Paul, V.; Zhang, W.; Kanawati, B.; Geist, B.; Faus-Keßler, T.; Schmitt-Kopplin, P.; Schäffner, A.R. The Arabidopsis glucosyltransferase UGT76B1 conjugates isoleucic acid and modulates plant defense and senescence. Plant Cell 2011, 23, 4124–4145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blum, M.; Chang, H.-Y.; Chuguransky, S.; Grego, T.; Kandasaamy, S.; Mitchell, A.; Nuka, G.; Paysan-Lafosse, T.; Qureshi, M.; Raj, S.; et al. The InterPro protein families and domains database: 20 years on. Nucleic Acids Res. 2021, 49, D344–D354. [Google Scholar] [CrossRef] [PubMed]
- Thomma, B.P.H.J.; Eggermont, K.; Penninckx, I.A.M.A.; Mauch-Mani, B.; Vogelsang, R.; Cammue, B.P.A.; Broekaert, W.F. Separate jasmonate-dependent and salicylate-dependent defense-response pathways in Arabidopsis are essential for resistance to distinct microbial pathogens. Proc. Natl. Acad. Sci. USA 1998, 95, 15107–15111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayat, S.; Hayat, Q.; Alyemeni, M.N.; Wani, A.S.; Pichtel, J.; Ahmad, A. Role of proline under changing environments. Plant Signal. Behav. 2012, 7, 1456–1466. [Google Scholar] [CrossRef] [Green Version]
- Daumann, M.; Fischer, M.; Niopek-Witz, S.; Girke, C.; Möhlmann, T. Apoplastic nucleoside accumulation in Arabidopsis leads to reduced photosynthetic performance and increased susceptibility against Botrytis cinerea. Front. Plant Sci. 2015, 6, 1158. [Google Scholar] [CrossRef]
- Zhang, J.; Jiang, L.; Sun, C.; Jin, L.; Lin, M.; Huang, Y.; Zheng, X.; Yu, T. Indole-3-acetic acid inhibits blue mold rot by inducing resistance in pear fruit wounds. Sci. Hortic. 2018, 231, 227–232. [Google Scholar] [CrossRef]
- Martínez Noël, G.M.A.; Madrid, E.A.; Bottini, R.; Lamattina, L. Indole acetic acid attenuates disease severity in potato-Phytophthora infestans interaction and inhibits the pathogen growth in vitro. Plant Physiol. Biochem. 2001, 39, 815–823. [Google Scholar] [CrossRef]
- Park, C.-J.; Seo, Y.-S. Heat shock proteins: A review of the molecular chaperones for plant immunity. Plant Pathol. J. 2015, 31, 323–333. [Google Scholar] [CrossRef] [Green Version]
- ul Haq, S.; Khan, A.; Ali, M.; Khattak, A.M.; Gai, W.X.; Zhang, H.X.; Wei, A.M.; Gong, Z.H. Heat shock proteins: Dynamic biomolecules to counter plant biotic and abiotic stresses. Int. J. Mol. Sci. 2019, 20, 5321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hýsková, V.; Bělonožníková, K.; Čeřovská, N.; Ryšlavá, H. HSP70 plays an ambiguous role during viral infections in plants. Biol. Plant. 2021, 65, 68–79. [Google Scholar] [CrossRef]
- Noël, L.D.; Cagna, G.; Stuttmann, J.; Wirthmüller, L.; Betsuyaku, S.; Witte, C.-P.P.; Bhat, R.; Pochon, N.; Colby, T.; Parker, J.E. Interaction between SGT1 and cytosolic/nuclear HSC70 chaperones regulates Arabidopsis immune responses. Plant Cell 2008, 19, 4061–4076. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Yan, J.; Wu, Y.; Zhang, H.; Mo, S.; Xu, X.; Zhou, F.; Ding, H. Proteomic analysis by iTRAQ-PRM provides integrated insight into mechanisms of resistance in pepper to Bemisia tabaci (Gennadius). BMC Plant Biol. 2019, 19, 270. [Google Scholar] [CrossRef]
- Klink, V.P.; Overall, C.C.; Alkharouf, N.W.; MacDonald, M.H.; Matthews, B.F. Laser capture microdissection (LCM) and comparative microarray expression analysis of syncytial cells isolated from incompatible and compatible soybean (Glycine max) roots infected by the soybean cyst nematode (Heterodera glycines). Planta 2007, 226, 1389–1409. [Google Scholar] [CrossRef]
- Szajko, K.; Plich, J.; Przetakiewicz, J.; Sołtys-Kalina, D.; Marczewski, W. Comparative proteomic analysis of resistant and susceptible potato cultivars during Synchytrium endobioticum infestation. Planta 2020, 251, 4. [Google Scholar] [CrossRef]
- Guo, H.; Zhang, H.; Wang, G.; Wang, C.; Wang, Y.; Liu, X.; Ji, W. Identification and expression analysis of heat-shock proteins in wheat infected with powdery mildew and stripe rust. Plant Genome 2021, 14, e20092. [Google Scholar] [CrossRef] [PubMed]
- Mambula, S.S.; Stevenson, M.A.; Ogawa, K.; Calderwood, S.K. Mechanisms for Hsp70 secretion: Crossing membranes without a leader. Methods 2007, 43, 168–175. [Google Scholar] [CrossRef] [Green Version]
- Yao, K.; De Luca, V.; Brisson, N. Creation of a metabolic sink for tryptophan alters the phenylpropanoid pathway and the susceptibility of potato to Phytophthora infestans. Plant Cell 1995, 7, 1787–1799. [Google Scholar] [CrossRef]
- Henriquez, M.A.; Adam, L.R.; Daayf, F. Alteration of secondary metabolites’ profiles in potato leaves in response to weakly and highly aggressive isolates of Phytophthora infestans. Plant Physiol. Biochem. 2012, 57, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Kröner, A.; Marnet, N.; Andrivon, D.; Val, F. Nicotiflorin, rutin and chlorogenic acid: Phenylpropanoids involved differently in quantitative resistance of potato tubers to biotrophic and necrotrophic pathogens. Plant Physiol. Biochem. 2012, 57, 23–31. [Google Scholar] [CrossRef]
- Yogendra, K.N.; Pushpa, D.; Mosa, K.A.; Kushalappa, A.C.; Murphy, A.; Mosquera, T. Quantitative resistance in potato leaves to late blight associated with induced hydroxycinnamic acid amides. Funct. Integr. Genom. 2014, 14, 285–298. [Google Scholar] [CrossRef] [PubMed]
- Fraser-Pitt, D.J.; Mercer, D.K.; Smith, D.; Kowalczuk, A.; Robertson, J.; Lovie, E.; Perenyi, P.; Cole, M.; Doumith, M.; Hill, R.L.R.; et al. Cysteamine, an endogenous aminothiol, and cystamine, the disulfide product of oxidation, increase Pseudomonas aeruginosa sensitivity to reactive oxygen and nitrogen species and potentiate therapeutic antibiotics against bacterial infection. Infect. Immun. 2018, 86, e00947-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brus-Szkalej, M.; Andersen, C.B.; Vetukuri, R.R.; Grenville-Briggs, L.J. A family of cell wall transglutaminases is essential for appressorium development and pathogenicity in Phytophthora infestans. Biorxiv 2021. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Tucker Hallmark, H.; Černý, M.; Brzobohatý, B.; Rashotte, A.M.A.M.; Hallmark, H.T.; Černý, M.; Brzobohatý, B.; Rashotte, A.M.A.M. trans-Zeatin-N-glucosides have biological activity in Arabidopsis thaliana. PLoS ONE 2020, 15, e0232762. [Google Scholar] [CrossRef] [PubMed]
- Perez-Riverol, Y.; Csordas, A.; Bai, J.; Bernal-Llinares, M.; Hewapathirana, S.; Kundu, D.J.; Inuganti, A.; Griss, J.; Mayer, G.; Eisenacher, M.; et al. The PRIDE database and related tools and resources in 2019: Improving support for quantification data. Nucleic Acids Res. 2019, 47, D442–D450. [Google Scholar] [CrossRef]
- Berka, M.; Luklová, M.; Dufková, H.; Malých, V.; Novák, J.; Saiz-Fernández, I.; Rashotte, A.M.; Brzobohaty, B.; Cerny, M. Barley root proteome and metabolome in response to cytokinin and abiotic stimuli. Front. Plant Sci. 2020, 11, 1647. [Google Scholar] [CrossRef] [PubMed]
- Saiz-Fernández, I.; Milenković, I.; Berka, M.; Černý, M.; Tomšovský, M.; Brzobohatý, B.; Kerchev, P. Integrated proteomic and metabolomic profiling of Phytophthora cinnamomi attack on sweet chestnut (Castanea sativa) reveals distinct molecular reprogramming proximal to the infection site and away from it. Int. J. Mol. Sci. 2020, 21, 8525. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein–protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dufková, H.; Berka, M.; Greplová, M.; Shejbalová, Š.; Hampejsová, R.; Luklová, M.; Domkářová, J.; Novák, J.; Kopačka, V.; Brzobohatý, B.; et al. The Omics Hunt for Novel Molecular Markers of Resistance to Phytophthora infestans. Plants 2022, 11, 61. https://doi.org/10.3390/plants11010061
Dufková H, Berka M, Greplová M, Shejbalová Š, Hampejsová R, Luklová M, Domkářová J, Novák J, Kopačka V, Brzobohatý B, et al. The Omics Hunt for Novel Molecular Markers of Resistance to Phytophthora infestans. Plants. 2022; 11(1):61. https://doi.org/10.3390/plants11010061
Chicago/Turabian StyleDufková, Hana, Miroslav Berka, Marie Greplová, Šarlota Shejbalová, Romana Hampejsová, Markéta Luklová, Jaroslava Domkářová, Jan Novák, Viktor Kopačka, Břetislav Brzobohatý, and et al. 2022. "The Omics Hunt for Novel Molecular Markers of Resistance to Phytophthora infestans" Plants 11, no. 1: 61. https://doi.org/10.3390/plants11010061