
A Systematic Review on Opuntia (Cactaceae; Opuntioideae) Flower-Visiting Insects in the World with Emphasis on Mexico: Implications for Biodiversity Conservation

 ,
,  , , and
, , and
Abstract
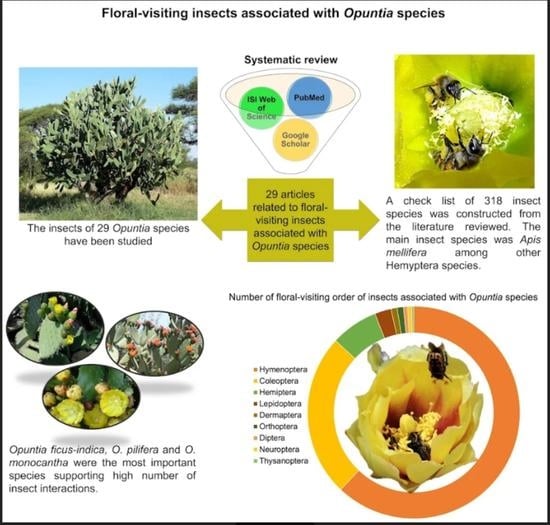
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
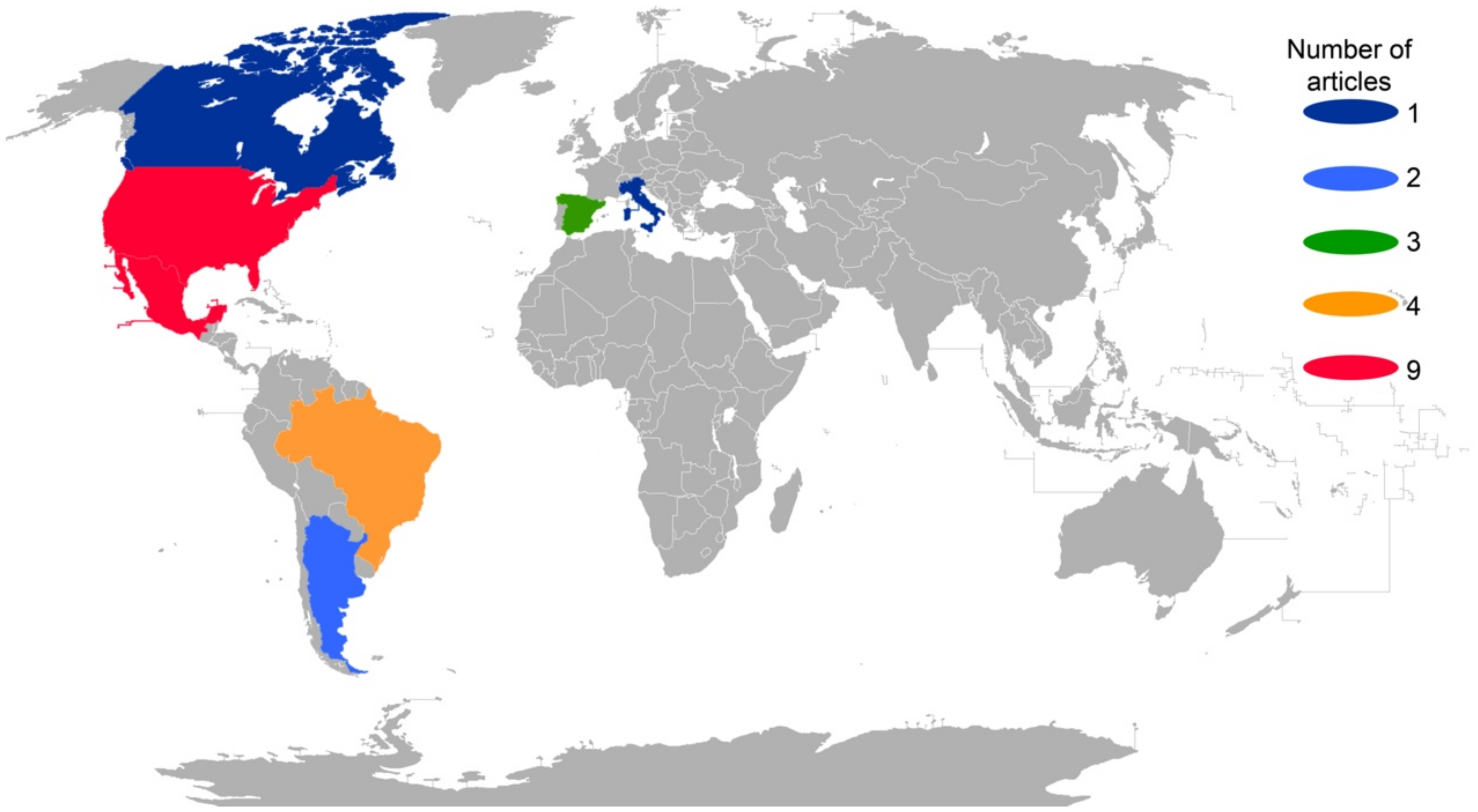
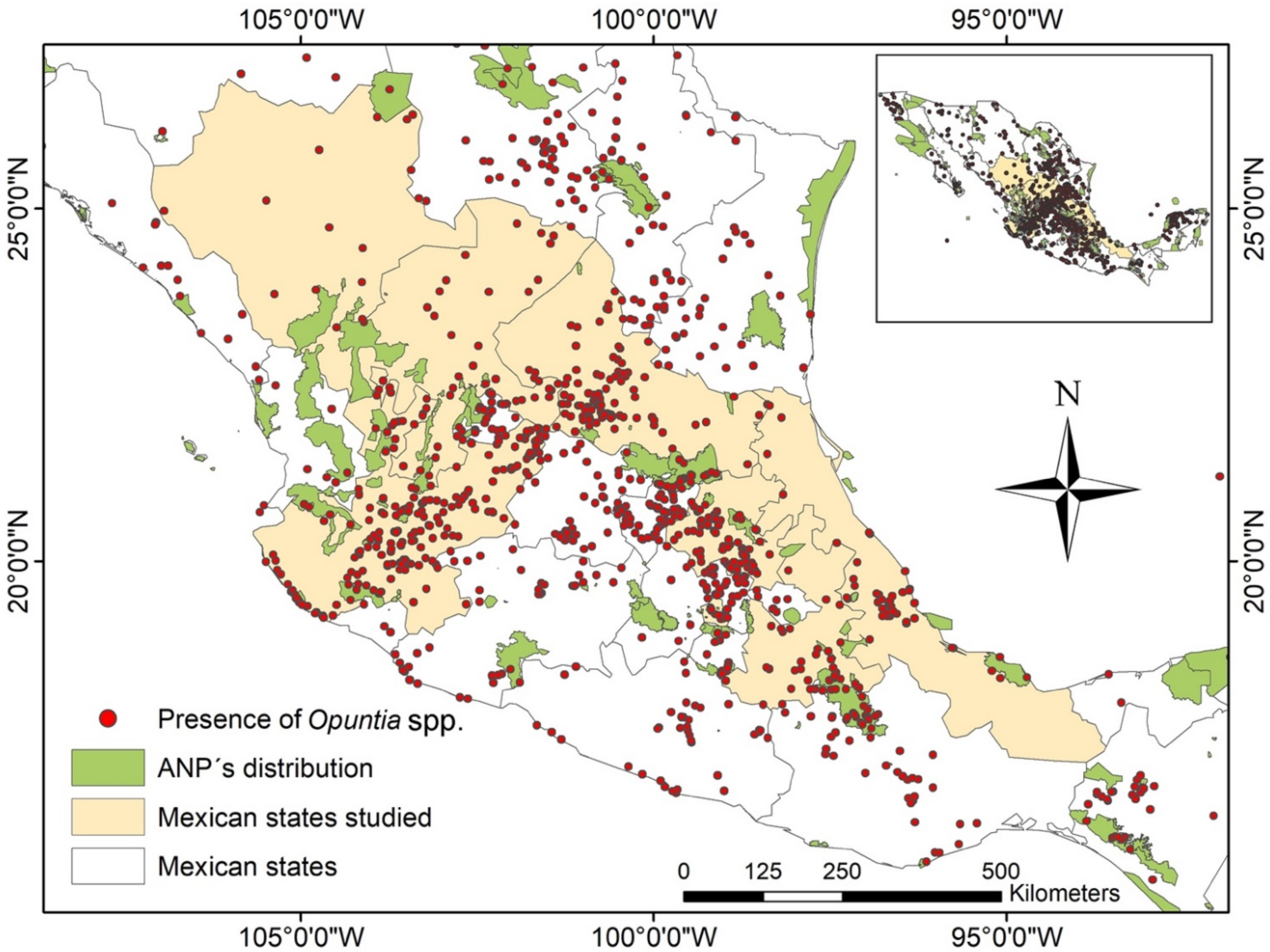
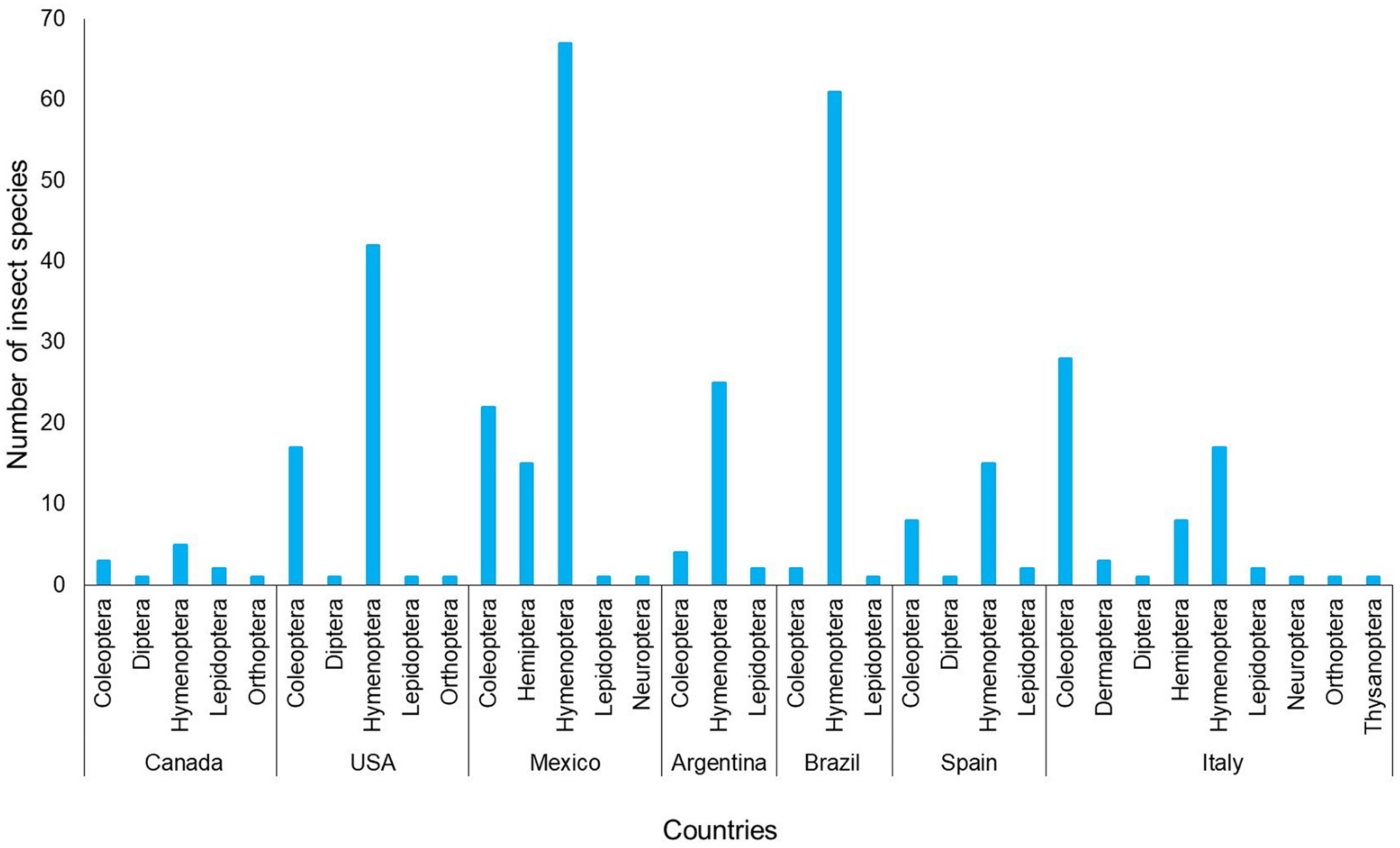
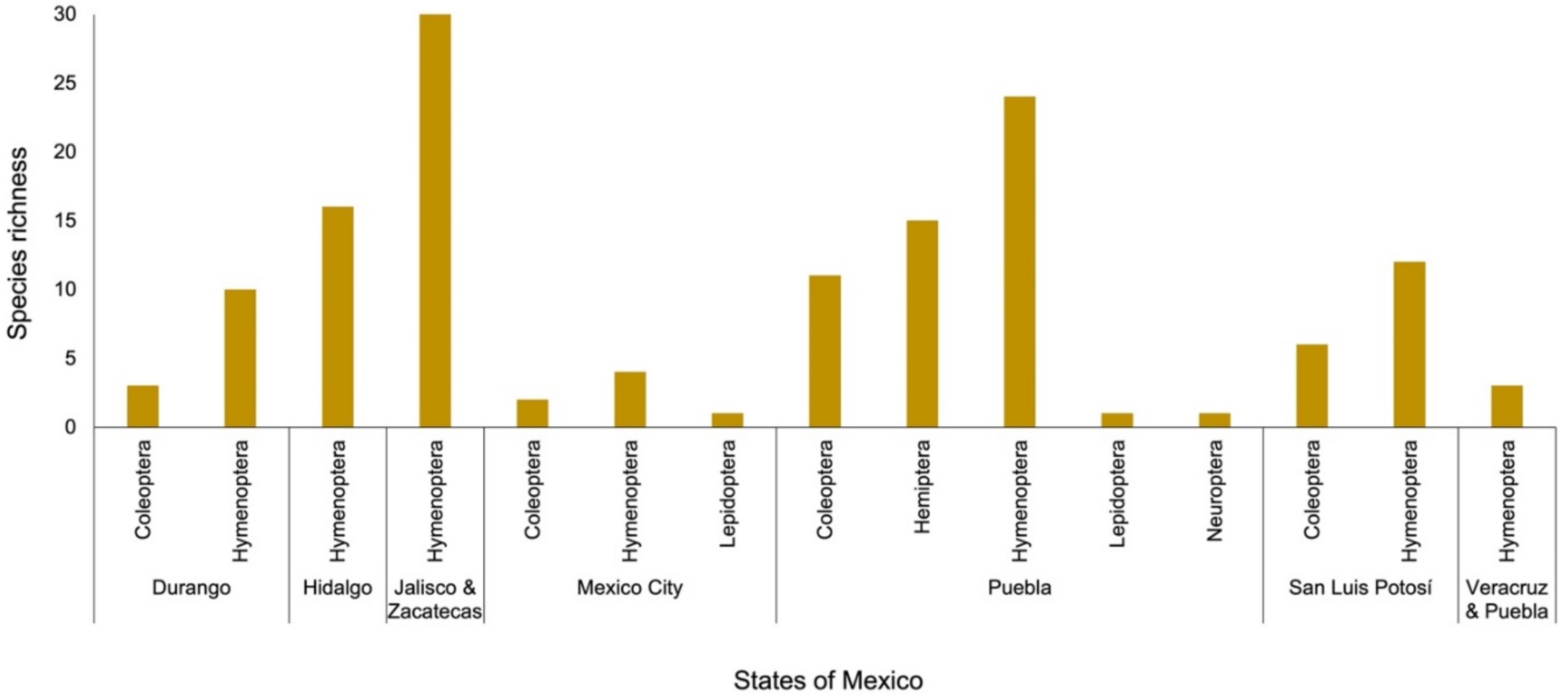
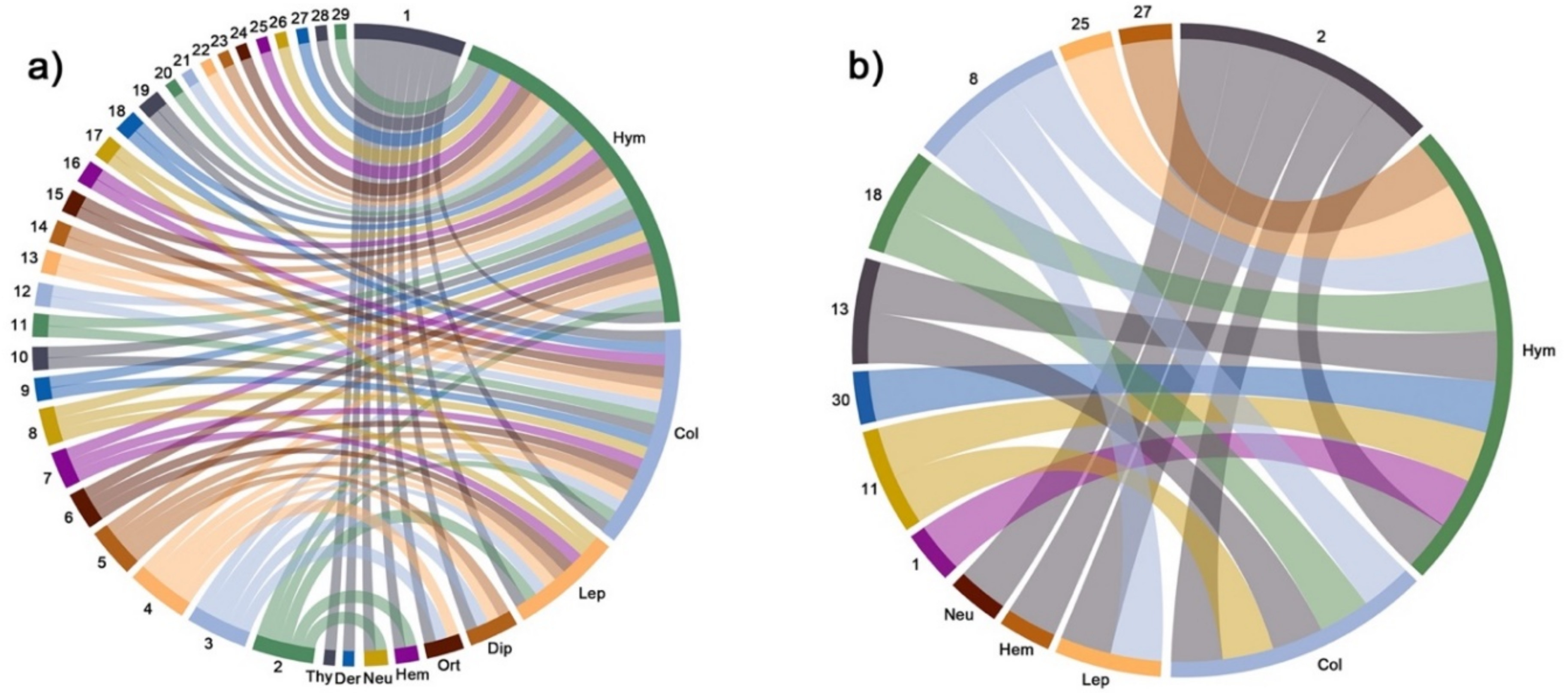
2. Results
3. Discussion
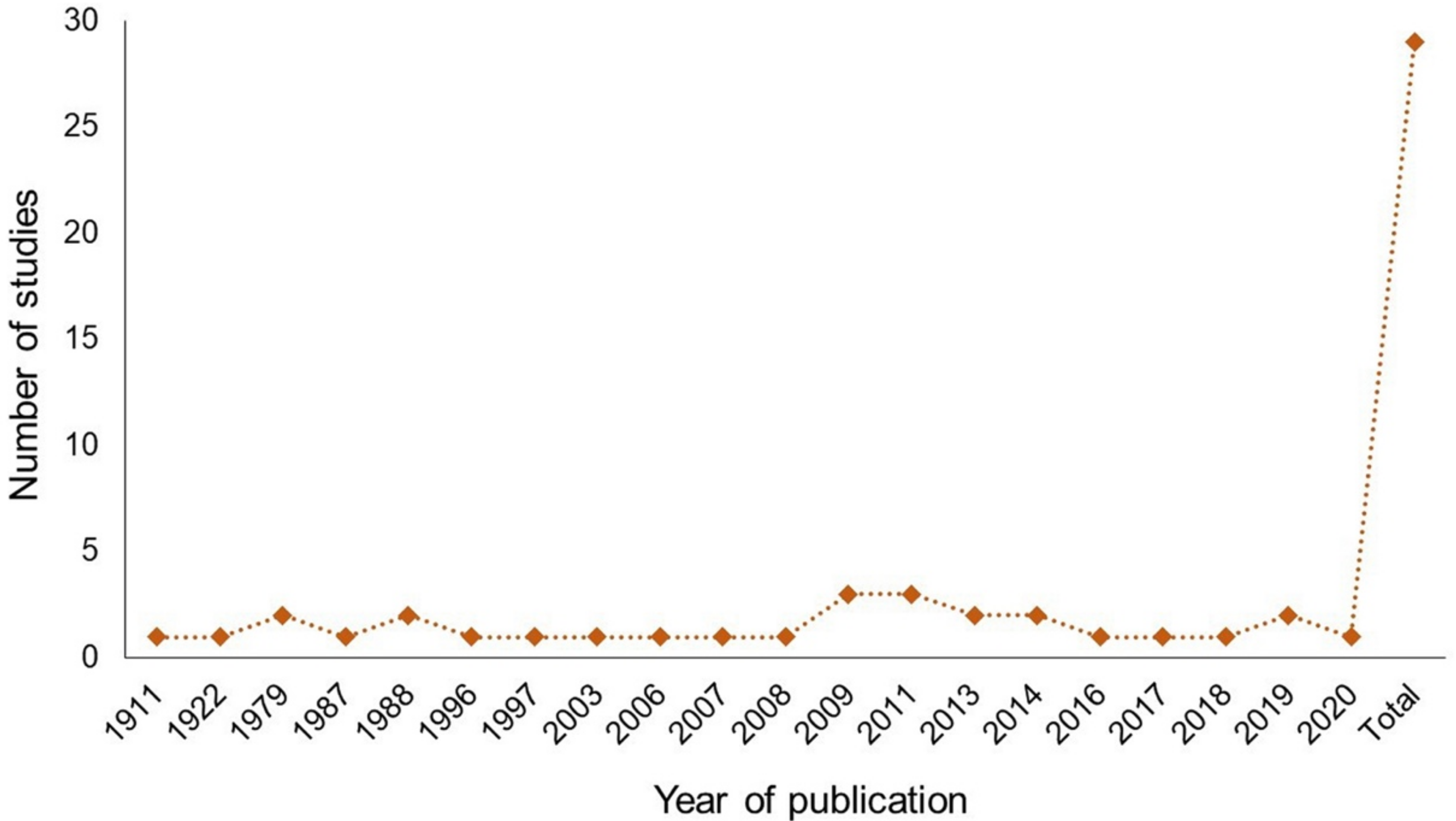
3.1. Timeline of Opuntia Studies
3.2. Insect Diversity: Are All the Species Efficient in Pollen Transport?
3.3. The Relationship between Opuntia and Insects in Mexico
3.4. The Role of Core Species in the Community Structure
3.5. Conservation Insights on Threatened Opuntia Species
4. Materials and Methods
5. Conclusions and Future Perspectives
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hernandez-Hernandez, T.; Brown, J.W.; Schlumpberger, B.O.; Eguiarte, L.E.; Magallón, S. Beyond aridification: Multiple explanations for the elevated diversification of cacti in the new world Succulent Biome. New Phytol. 2014, 202, 1382–1397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, E.F. The Cactus Family; Timber Press: Portland, OR, USA, 2001; p. 776. [Google Scholar]
- Hunt, D. The New Cactus Lexicon; DH Books: Milborne Port, UK, 2006; Volumes 1 and 2, p. 925. [Google Scholar]
- Guerrero, P.C.; Majure, L.C.; Cornejo-Romero, A.; Hernández-Hernández, T. Phylogenetic relationships and evolutionary trends in the cactus family. J. Hered. 2019, 110, 4–21. [Google Scholar] [CrossRef] [PubMed]
- González-Medrano, F. Las Zonas Áridas y Semiáridas de México y su Vegetación; Instituto Nacional de Ecología–Secretaria del Medio Ambiente y Recursos Naturales: Mexico City, Mexico, 2012; p. 194. [Google Scholar]
- Goettsch, B.; Hilton-Taylor, C.; Cruz-Piñón, G.; Duffy, J.P.; Frances, A.; Hernández, H.M.; Inger, R.; Pollock, C.; Schipper, J.; Superina, M.; et al. High proportion of cactus species threatened with extinction. Nat. Plants 2015, 1, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiménez-Sierra, C.L. Las cactáceas mexicanas y los riesgos que enfrentan. Revista Digital Universitaria 2011, 12, 5–13. [Google Scholar]
- Majure, L.C.; Puente, R. Phylogenetic relationships and morphological evolution in Opuntia s. str. and closely related members of tribe Opuntieae. Succ. Plant Res. 2014, 8, 9–30. [Google Scholar]
- Majure, L.C.; Puente, R.; Griffith, M.P.; Judd, W.S.; Soltis, P.S.; Soltis, D.E. Phylogeny of Opuntia s.s. (Cactaceae): Clade delineation, geographic origins, and reticulate evolution. Am. J. Bot. 2012, 99, 847–864. [Google Scholar] [CrossRef] [Green Version]
- Aliscioni, N.L.; Delbón, N.E.; Gurvich, D.E. Spine function in Cactaceae, a review. J. Prof. Assoc. Cactus 2021, 23, 1–11. [Google Scholar]
- González-Elizondo, M.S.; González-Elizondo, M.; López-Enríquez, I.L.; Tena-Flores, J.A.; González-Gallegos, J.G.; Ruacho-González, L.; Melgoza-Castillo, A.; Villarreal-Quintanilla, J.A.; Estrada-Castillón, A.E. Diagnóstico del conocimiento taxonómico y florístico de las plantas vasculares del norte de México. Bot. Sci. 2017, 95, 760–779. [Google Scholar] [CrossRef] [Green Version]
- Flores Valdez, C.A.; Aguirre Rivera, J.R. El Nopal como Forraje; Universidad Autónoma Chapingo: Texcoco, México, 1979; p. 91. [Google Scholar]
- Anaya-Pérez, M.A.; Bautista-Zane, R. El nopal forrajero en México: Del siglo XVI al siglo XX. Agric. Soc. Desarro. 2008, 5, 167–183. [Google Scholar]
- Ortega-Baes, P.; Sühring, S.; Sajama, J.; Sotola, E.; Alonso-Pedano, M.; Bravo, S.; Godínez-Alvarez, H. Diversity and conservation in the Cactus family. In Desert Plants. Biology and Biotechnology, 1st ed.; Ramawat, K., Ed.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 157–173. [Google Scholar] [CrossRef]
- Dubeux, J.C.B., Jr.; Dos Santos, M.V.F.; Da Cunha, M.V.; Dos Santos, D.C.; De Almeida Souza, R.T.; De Mello, A.C.L.; De Souza, T.C. Cactus (Opuntia and Nopalea) nutritive value: A review. Anim. Feed Sci. and Tech. 2021, 275, 114890. [Google Scholar] [CrossRef]
- Ciriminna, R.; Chavarría-Hernández, N.; Rodríguez-Hernández, A.I.; Pagliaro, M. Toward unfolding the bioeconomy of nopal (Opuntia spp.). Biofuel Bioprod. Bior. 2019, 13, 1417–1427. [Google Scholar] [CrossRef] [Green Version]
- Le Houerou, H.N. Utilization of fodder trees and shrubs in the arid and semiarid zones of West Asia and North Africa. Arid Soil Res. Rehabil. 2000, 14, 101–135. [Google Scholar] [CrossRef]
- Silva, M.A.; Albuquerque, T.G.; Pereira, P.; Ramalho, R.; Vicente, F.; Oliveira, M.B.P.; Costa, H.S. Opuntia ficus-indica (L.) Mill.: A multi-benefit potential to be exploited. Molecules 2021, 26, 951. [Google Scholar] [CrossRef]
- Bartomeus, I.; Vilà, M. Breeding system and pollen limitation in two supergeneralist alien plants invading Mediterranean shrublands. Aust. J. Bot. 2009, 57, 109–115. [Google Scholar] [CrossRef]
- Padrón, B.; Traveset, A.; Biedenweg, T.; Díaz, D.; Nogales, M.; Olesen, J.M. Impact of alien plant invaders on pollination networks in two archipelagos. PLoS ONE 2009, 4, e6275. [Google Scholar] [CrossRef] [Green Version]
- Instituto Nacional de Estadística y Geografía (INEGI). Características Principales del Cultivo del Nopal en el Distrito Federal Caso Milpa Alta; Censo Agropecuario 2007; Instituto Nacional de Estadística y Geografía: Aguascalientes, Mexico, 2007; p. 68. [Google Scholar]
- Rebman, J.P.; Pinkava, D.J. Opuntia cacti of North America: An overview. Fla. Entomol. 2001, 84, 474–483. [Google Scholar] [CrossRef]
- Reyes-Agüero, J.A.; Valiente-Banuet, A. Reproductive biology of Opuntia: A review. J. Arid Environ. 2006, 64, 549–585. [Google Scholar] [CrossRef]
- Inglese, P.; Mondragon, C.; Nefzaoui, A.; Saenz, C.; Taguchi, M.; Makkar, H.; Louhaichi, M. Ecologia del Cultivo, Manejo y Usos del Nopal; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2018; p. 229. Available online: https://hdl.handle.net/20.500.11766/9380 (accessed on 22 October 2021).
- Mandujano, M.C.; Carrillo-Ángeles, I.; Martínez Peralta, C.; Golubov, J. Reproductive biology of Cactaceae. In Desert Plants. Biology and Biotechnology, 1st ed.; Ramawat, K., Ed.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 197–230. [Google Scholar] [CrossRef]
- Kearns, C.A.; Inouye, D.W.; Waser, N.M. Endangered mutualisms: The conservation of plant-pollinator interactions. Annu. Rev. Ecol. Syst. 1998, 29, 83–112. [Google Scholar] [CrossRef]
- Taki, H.; Kevan, P.G. Does habitat loss affect the communities of plants and insects equally in plant–pollinator interactions? Preliminary findings. Biodivers. Conserv. 2007, 16, 3147–3161. [Google Scholar] [CrossRef] [Green Version]
- Senapathi, D.; Biesmeijer, J.C.; Breeze, T.D.; Kleijn, D.; Potts, S.G.; Carvalheiro, L.G. Pollinator conservation—The difference between managing for pollination services and preserving pollinator diversity. Curr. Opin. Insect Sci. 2015, 12, 93–101. [Google Scholar] [CrossRef] [Green Version]
- Potts, S.G.; Biesmeijer, J.C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W.E. Global pollinator declines: Trends, impacts and drivers. Trends Ecol. Evol. 2010, 25, 345–353. [Google Scholar] [CrossRef]
- Van der Kooi, C.J.; Vallejo-Marín, M.; Leonhardt, S.D. Mutualisms and (A) symmetry in plant-pollinator interactions. Curr. Biol. 2021, 31, 91–99. [Google Scholar] [CrossRef]
- Morales-Trejo, J.J.; Sandoval-Ruiz, C.A.; Fascinetto-Zago, P.; Cruzado-Lima, A.L.; Vázquez-Hernández, C. Abundancia y diversidad de visitadores florales de Opuntia pilifera en Zapotitlán Salinas, Puebla. Entomol. Mex. 2014, 1, 1144–1148. [Google Scholar]
- Pimienta, B.E.; Del Castillo, R.F. Reproductive biology. In Cacti: Biology and Uses; Nobel, P.S., Ed.; University of California Press: Los Angeles, CA, USA, 2002; pp. 75–90. [Google Scholar] [CrossRef]
- Charbonneau, D.; Blonder, B.; Dornhaus, A. Social insects: A model system for network dynamics. In Temporal Networks, Understanding Complex Systems; Holme, P., Saramaki, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 217–244. [Google Scholar] [CrossRef]
- Dáttilo, W.; Fagundes, R.; Gurka, C.A.; Silva, M.S.; Vieira, M.C.; Izzo, T.J.; Díaz-Castelazo, C.; Del-Claro, K.; Rico-Gray, V. Individual-based ant-plant networks: Diurnal-nocturnal structure and species-area relationship. PLoS ONE 2014, 9, e99838. [Google Scholar] [CrossRef] [Green Version]
- Ávila-Gómez, E.S.; Meléndez-Ramírez, V.; Castellanos, I.; Zuria, I.; Moreno, C.E. Prickly pear crops as bee diversity reservoirs and the role of bees in Opuntia fruit production. Agric. Ecosyst. Environ. 2019, 279, 80–88. [Google Scholar] [CrossRef]
- González, A.B.D.J.; García, A.A.; Olguín, J.F.L.; Rivera, A.; Martínez, V.L. Entomofauna asociada al nopal verdura (Opuntia ficusindica Miller) en San Andrés Cholula, Puebla, México. Southwest Entomol. 2016, 41, 259–266. [Google Scholar] [CrossRef]
- Del Castillo, R.; González-Espinosa, M. Una interpretación evolutiva del polimorfismo sexual de Opuntia robusta (Cactaceae). Agrociencia 1988, 71, 185–196. [Google Scholar]
- Mandujano, M.D.C.; Montaña, C.; Eguiarte, L.E. Reproductive ecology and inbreeding depression in Opuntia rastrera (Cactaceae) in the Chihuahuan Desert: Why are sexually derived recruitments so rare? Am. J. Bot. 1996, 83, 63–70. [Google Scholar] [CrossRef]
- Mandujano, M.C.; Plasencia-López, L.; Aguilar-Morales, G.; Jiménez-Guzmán, G.; Galicia-Pérez, A.; Rojas-Aréchiga, M.; Martínez-Peralta, C. Sistema sexual de Opuntia tomentosa Salm-Dyck (Cactaceae, Opuntioideae) en un pedregal de origen volcánico. Cactáceas Suculentas Mex. 2014, 59, 100–120. [Google Scholar]
- Muñoz-Urías, A.; Palomino-Hasbach, G.; Huerta-Martínez, F.M.; Pimienta-Barrios, E.; Ramírez-Hernández, B.C. Reproductive isolation in fragmented wild populations of Opuntia streptacantha. J. Prof. Assoc. Cactus Dev. 2006, 8, 26–38. [Google Scholar]
- Piña, H.H.; Montaña, C.; Mandujano, M. Fruit abortion in the Chihuahuan-Desert endemic cactus Opuntia microdasys. Plant Ecol. 2007, 193, 305–313. [Google Scholar] [CrossRef]
- Sánchez-Echeverría, K.; Castellanos, I.; Mendoza-Cuenca, L.F. Abejas visitantes florales de Opuntia heliabravoana en un gradiente de urbanización. Biológicas 2016, 18, 27–34. [Google Scholar]
- Santa Anna-Aguayo, A.I.; Schaffner, C.M.; Golubov, J.; López-Portillo, J.; García-Franco, J.; Herrera-Meza, G.; Martínez, A.J. Behavioral repertoires and interactions between Apis mellifera (Hymenoptera: Apidae) and the native bee Lithurgus littoralis (Hymenoptera: Megachilidae) in flowers of Opuntia huajuapensis (Cactaceae) in the Tehuacan desert. Fla. Entomol. 2017, 100, 396–402. [Google Scholar] [CrossRef]
- Riojas-López, M.E.; Díaz-Herrera, I.A.; Fierros-López, H.E.; Mellink, E. The effect of adjacent habitat on native bee assemblages in a perennial low-input agroecosystem in a semiarid anthropized landscape. Agric. Ecosyst. Environ. 2019, 272, 199–205. [Google Scholar] [CrossRef]
- Gallegos-Vázquez, C.; Méndez-Gallegos, S.D.J.; Mondragón, J.C. Producción Sustentable de Tuna en San Luis Potosí; Colegio de Postgraduados–Fundación Produce San Luis Potosí: San Luis Potosí, Mexico, 2013; p. 203. [Google Scholar]
- Potgieter, J.; D’Aquino, S. Fruit production and post-harvest management. In Ecologia del Cultivo, Manejo y Usos del Nopal; Inglese, P., Mondragon, C., Nefzaoui, A., Saenz, C., Taguchi, M., Makkar, H., Louhaichi, M., Eds.; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2018; pp. 51–71. Available online: https://hdl.handle.net/20.500.11766/9380 (accessed on 13 December 2021).
- Riojas-López, M.; Mellink, E. Potential for biological conservation in man-modified semiarid habitats in northeastern Jalisco, Mexico. Biodivers. Conserv. 2005, 14, 2251–2263. [Google Scholar] [CrossRef]
- Bembower, W. Pollination notes from the Cedar Point region. Ohio Nat. 1911, 9, 378–383. [Google Scholar]
- Cockerell, T.D.A. Two new subgenera of north American bees. Am. Mus. Novit 1922, 47, 1–5. [Google Scholar]
- Grant, V.; Grant, K.A. Pollination of Opunta basilaris and O. littoralis. Plant Syst. Evol. 1979, 132, 321–325. [Google Scholar] [CrossRef]
- Grant, V.; Grant, K.A.; Hurd, P.D. Pollination of Opuntia lindheimeri and related species. Plant Syst. Evol. 1979, 132, 313–320. [Google Scholar] [CrossRef]
- Fachardo, A.L.S.; Sigrist, M.R. Pre-zygotic reproductive isolation between two synchronopatric Opuntia (Cactaceae) species in the Brazilian Chaco. Plant Biol. 2020, 22, 487–493. [Google Scholar] [CrossRef]
- Pretto, F.; Celesti-Grapow, L.; Carli, E.; Blasi, C. Influence of past land use and current human disturbance on non-native plant species on small Italian islands. Plant Ecol. 2010, 210, 225–239. [Google Scholar] [CrossRef]
- Lo Verde, G.; La Mantia, T. The role of native flower visitors in pollinating Opuntia ficus-indica (L.) Mill., naturalized in Sicily. Acta Oecologica 2011, 37, 413–417. [Google Scholar] [CrossRef]
- Bartomeus, I.; Vilà, M.; Santamaría, L. Contrasting effects of invasive plants in plant–pollinator networks. Oecologia 2008, 155, 761–770. [Google Scholar] [CrossRef]
- Jauker, F.; Speckmann, M.; Wolters, V. Intra-specific body size determines pollination effectiveness. Basic Appl. Ecol. 2016, 17, 714–719. [Google Scholar] [CrossRef]
- Osborn, M.M.; Kevan, P.G.; Lane, M.A. Pollination biology of Opuntia polyacantha and Opuntia phaeacantha (Cactaceae) in southern Colorado. Plant Syst. Evol. 1988, 159, 85–94. [Google Scholar] [CrossRef]
- Cota-Sánchez, J.H.; Almeida, O.J.G.; Falconer, D.J.; Choi, H.J.; Bevan, J. Intriguing thigmonastic (sensitive) stamens in the plains prickly pear Opuntia polyacantha (Cactaceae). Flora 2013, 208, 381–389. [Google Scholar] [CrossRef]
- Schlindwein, C.; Wittmann, D. Stamen movements in flowers of Opuntia (Cactaceae) favour oligolectic pollinators. Plant Syst. Evol. 1997, 204, 179–193. [Google Scholar] [CrossRef]
- Lenzi, M.; Orth, A.I. Floral visitors of the Opuntia monacantha (Cactaceae) in sandbank of the Florianópolis, SC, Brazil. Acta Biológica Paranaense 2011, 40, 19–32. [Google Scholar] [CrossRef] [Green Version]
- Gómez, J.M. Effectiveness of ants as pollinators of Lobularia maritima: Effects on main sequential fitness components of the host plant. Oecologia 2000, 122, 90–97. [Google Scholar] [CrossRef]
- Maubecin, C.C.; Boero, L.; Sérsic, A.N. Specialisation in pollen collection, pollination interactions and phenotypic variation of the oil-collecting bee Chalepogenus cocuccii. Apidologie 2020, 51, 710–723. [Google Scholar] [CrossRef]
- Arroyo-Pérez, E.; Jiménez-Sierra, C.L.; Zavala Hurtado, J.A.; Flores, J. Shared pollinators and sequential flowering phenologies in two sympatric cactus species. Plant Ecol. Evo. 2021, 154, 28–38. [Google Scholar] [CrossRef]
- Samra, S.; Samocha, Y.; Eisikowitch, D.; Vaknin, Y. Can ants equal honeybees as effective pollinators of the energy crop Jatropha curcas L. under Mediterranean conditions? GCB Bioenergy 2014, 6, 756–767. [Google Scholar] [CrossRef] [Green Version]
- Pickett, C.H.; Clark, W.D. The function of extrafloral nectaries in Opuntia acanthocarpa (Cactaceae). Am. J. Bot. 1979, 66, 618–625. [Google Scholar] [CrossRef]
- LeVan, K.E.; Hung, K.L.J.; McCann, K.R.; Ludka, J.T.; Holway, D.A. Floral visitation by the Argentine ant reduces pollinator visitation and seed set in the coast barrel cactus, Ferocactus viridescens. Oecologia 2014, 174, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Mauseth, J.D.; Rebmann, J.P.; Machado, S.R. Extrafloral nectaries in cacti. Cactus Succul. J. 2016, 88, 156–171. [Google Scholar] [CrossRef]
- Wagner, D.; Kay, A. Do extrafloral nectaries distract ants from visiting flowers? An experimental test of an overlooked hypothesis. Evol. Ecol. Res. 2002, 4, 293–305. [Google Scholar]
- Komamura, R.; Koyama, K.; Yamauchi, T.; Konno, Y.; Gu, L. Pollination contribution differs among insects visiting Cardiocrinum cordatum flowers. Forests 2021, 12, 452. [Google Scholar] [CrossRef]
- Beattie, A.J.; Turnbull, C.; Knox, R.B.; Williams, E.G. Ant inhibition of pollen function—A possible reason why ant pollination is rare. Am. J. Bot. 1984, 71, 421–426. [Google Scholar] [CrossRef]
- Rostás, M.; Bollmann, F.; Saville, D.; Riedel, M. Ants contribute to pollination but not to reproduction in a rare calcareous grassland forb. PeerJ 2018, 6, e4369. [Google Scholar] [CrossRef] [Green Version]
- Rostás, M.; Tautz, J. Ants as pollinators of plants and the role of floral scents. In All Flesh is Grass. Cellular Origin, Life in Extreme Habitats and Astrobiology, 1st ed.; Dubinsky, Z., Seckbach, J., Eds.; Springer: Dordrecht, The Netherlands, 2010; Volume 16, pp. 149–161. [Google Scholar] [CrossRef]
- Gallai, N.; Salles, J.M.; Settele, J.; Vaissière, B.E. Economic valuation of the vulnerability of world agriculture confronted with pollinator decline. Ecol. Econ. 2009, 68, 810–821. [Google Scholar] [CrossRef]
- Rzedowski, J. Las principales zonas áridas de México y su vegetación. Bios Revista del Seminario de Estudios Biológicos 1968, 1, 4–24. [Google Scholar]
- Soberon, J.; Golubov, J.; Sarukhán, J. The importance of Opuntia in Mexico and routes of invasion and impact of Cactoblastis cactorum (Lepidoptera: Pyralidae). Fla. Entomol. 2001, 84, 486–492. [Google Scholar] [CrossRef]
- Vanegas-Rico, J.M.; Lomeli–Flores, J.R.; Rodríguez–Leyva, E.; Mora–Aguilera, G.; Valdez, J.M. Natural enemies of Dactylopius opuntiae (Cockerell) on Opuntia ficus-indica (L.) Miller in Central México. Acta Zoológica Mex. 2010, 26, 415–433. [Google Scholar]
- McCravy, K.W. A review of sampling and monitoring methods for beneficial arthropods in agroecosystems. Insects 2018, 9, 170. [Google Scholar] [CrossRef] [Green Version]
- Montgomery, G.A.; Belitz, M.W.; Guralnick, R.P.; Tingley, M.W. Standards and best practices for monitoring and benchmarking insects. Front. Ecol. Evol. 2021, 8, 579193. [Google Scholar] [CrossRef]
- Vigueras, A.L.; Portillo, L. Uses of Opuntia species and the potential impact of Cactoblastis cactorum (Lepidoptera: Pyralidae) in Mexico. Fla. Entomol. 2001, 84, 493–498. [Google Scholar] [CrossRef]
- Griffith, M.P. The origins of an important cactus crop, Opuntia ficus-indica (Cactaceae): New molecular evidence. Am. J. Bot. 2004, 91, 1915–1921. [Google Scholar] [CrossRef] [Green Version]
- Barbera, G.; Carimi, F.; Inglese, P. Past and present role of the indian-fig prickly-pear (Opuntia ficus-indica (L) Miller, Cactaceae) in the agriculture of Sicily. Econ. Bot. 1992, 46, 10–20. [Google Scholar] [CrossRef]
- Stang, M.; Klinkhamer, P.G.; Van Der Meijden, E. Size constraints and flower abundance determine the number of interactions in a plant–flower visitor web. Oikos 2006, 112, 111–121. [Google Scholar] [CrossRef] [Green Version]
- Lopezaraiza-Mikel, M.E.; Hayes, R.B.; Whalley, M.R.; Memmott, J. The impact of an alien plant on a native plant–pollinator network: An experimental approach. Ecol. Lett. 2007, 10, 539–550. [Google Scholar] [CrossRef]
- Ness, J.H. Hot spots and hot moments for on-plant foraging by ants within the flora of warm North American Deserts. Am. Midl. Nat. 2020, 183, 145–163. [Google Scholar] [CrossRef]
- Vithanage, V. The role of the European honeybee (Apis mellifera L.) in avocado pollination. J. Hortic. Sci. 1990, 65, 81–86. [Google Scholar] [CrossRef]
- Rizzardo, R.A.; Milfont, M.O.; Silva, E.; Freitas, B.M. Apis mellifera pollination improves agronomic productivity of anemophilous castor bean (Ricinus communis). Anais da Academia Brasileira de Ciências 2012, 84, 1137–1145. [Google Scholar] [CrossRef] [Green Version]
- Moritz, R.F.; Härtel, S.; Neumann, P. Global invasions of the western honeybee (Apis mellifera) and the consequences for biodiversity. Ecoscience 2005, 12, 289–301. [Google Scholar] [CrossRef]
- De la Rúa, P.; Paxton, R.J.; Moritz, R.F.A.; Roberts, S.; Allen, D.J.; Pinto, M.A.; Cauia, E.; Fontana, P.; Kryger, P.; Bouga, M.; et al. Apis mellifera. The IUCN Red List of Threatened Species 2014, e.T42463639A42463665. Available online: https://ec.europa.eu/environment/nature/conservation/species/redlist/downloads/European_bees.pdf (accessed on 13 March 2021).
- Paudel, Y.P.; Mackereth, R.; Hanley, R.; Qin, W. Honeybees (Apis mellifera L.) and pollination issues: Current status, impacts, and potential drivers of decline. J. Agric. Sci. 2015, 7, 93–109. [Google Scholar]
- Grixti, J.C.; Wong, L.T.; Cameron, S.A.; Favret, C. Decline of bumble bees (Bombus) in the North American Midwest. Biol. Conserv. 2009, 142, 75–84. [Google Scholar] [CrossRef]
- Jacobson, M.M.; Tucker, E.M.; Mathiasson, M.E.; Rehan, S.M. Decline of bumble bees in northeastern North America, with special focus on Bombus terricola. Biol. Conserv. 2018, 217, 437–445. [Google Scholar] [CrossRef]
- Hatfield, R.; Jepsen, S.; Thorp, R.; Richardson, L.; Colla, S.; Foltz Jordan, S. Bombus pensylvanicus. The IUCN Red List of Threatened Species 2015, e.T21215172A21215281. Available online: https://dx.doi.org/10.2305/IUCN.UK.2015-4.RLTS.T21215172A21215281.en (accessed on 25 February 2021).
- SEMARNAT. Norma Oficial Mexicana NOM-059-SEMARNAT-2010, Protección Ambiental-Especies Nativas de México de Flora y Fauna Silvestres-Categorías de Riesgo y Especificaciones para su Inclusión, Exclusión o Cambio-Lista de Especies en Riesgo; Diario Oficial de la Federación: Mexico City, Mexico, 2010; p. 78. [Google Scholar]
- Hernández, H.M.; Gómez-Hinostrosa, C.; Goettsch, B.K. Opuntia chaffeyi. The IUCN Red List of Threatened Species 2013, e.T41222A2952609. Available online: https://dx.doi.org/10.2305/IUCN.UK.2013-1.RLTS.T41222A2952609.en (accessed on 22 April 2021).
- Hernández, H.M.; Gómez-Hinostrosa, C.; Goettsch, B.K.; Sotomayor, M. Opuntia megarrhiza. The IUCN Red List of Threatened Species 2013, e.T41219A2952324. Available online: https://dx.doi.org/10.2305/IUCN.UK.2013-1.RLTS.T41219A2952324.en (accessed on 22 April 2021).
- Majure, L.; Griffith, P. Opuntia abjecta. The IUCN Red List of Threatened Species 2013, e.T199640A2608155. Available online: https://dx.doi.org/10.2305/IUCN.UK.2013-1.RLTS.T199640A2608155.en (accessed on 22 April 2021).
- Nassar, J.; Majure, L.; Griffith, P. Opuntia schumannii. The IUCN Red List of Threatened Species 2017, e.T152123A121580174. Available online: https://dx.doi.org/10.2305/IUCN.UK.2017-3.RLTS.T152123A121580174.en (accessed on 22 April 2021).
- Majure, L.; Griffith, P.; Gann, G.D. Opuntia triacantha. The IUCN Red List of Threatened Species 2017, e.T152237A121584692. Available online: https://dx.doi.org/10.2305/IUCN.UK.2017-3.RLTS.T152237A121584692.en (accessed on 22 April 2021).
- Nassar, J.; Majure, L. Opuntia curassavica (Amended Version of 2013 Assessment). The IUCN Red List of Threatened Species 2017, e.T152801A121610952. Available online: https://dx.doi.org/10.2305/IUCN.UK.2017-3.RLTS.T152801A121610952.en (accessed on 22 April 2021).
- Negrón-Ortiz, V. Reproductive biology of a rare cactus, Opuntia spinosissima (Cactaceae), in the Florida Keys: Why is seed set very low? Sex. Plant Reprod. 1998, 11, 208–212. [Google Scholar] [CrossRef]
- Areces-Mallea, A.E. A new opuntioid cactus from the Cayman Islands, B.W.I., with a discussion and key to the genus Consolea Lemaire. Brittonia 2001, 53, 96–107. [Google Scholar] [CrossRef]
- Locatelli, E.; Machado, I.C.S. Comparative study of the floral biology in two ornithophilous species of Cactaceae: Melocactus zehntneri and Opuntia palmadora. Bradleya 1999, 17, 75–85. [Google Scholar] [CrossRef]
- Akcelrad Lerner, L. Comisión Nacional Para el Conocimiento y Uso de la Biodiversidad C. Especies Silvestres de Nopales Mexicanos. Version 1.9. Comisión Nacional Para el Conocimiento y Uso de la Biodiversidad. Occurrence Dataset. 2021. Available online: GBIF.org (accessed on 17 December 2021). [CrossRef]
- Jordano, P.; Vázquez, D.; Bascompte, J. Redes complejas de interacciones mutualistas planta-animal. In Ecología y Evolución de Interacciones Planta-Animal, 1st ed.; Medel, R., Aizen, M.A., Zamora, R., Eds.; Editorial Universitaria: Santiago, Chile, 2009; pp. 17–41. [Google Scholar]
- Dáttilo, W.; Guimarães, P.R., Jr.; Izzo, T.J. Spatial structure of ant–plant mutualistic networks. Oikos 2013, 122, 1643–1648. [Google Scholar] [CrossRef]
- Agüero, J.I.; Galati, B.G.; Torretta, J.P. Structure and ultrastructure of floral nectaries of two Opuntia species (Cactaceae) in relation to their floral visitors. Plant Syst. Evol. 2018, 304, 1057–1067. [Google Scholar] [CrossRef]
- Díaz, L.; Cocucci, A.A. Functional gynodioecy in Opuntia quimilo (Cactaceae), a tree cactus pollinated by bees and hummingbirds. Plant Biol. 2003, 5, 531–539. [Google Scholar] [CrossRef]
- Janeba, Z. Insect flower visitors and pollinators of cacti from the southwest USA. Bradleya 2009, 27, 59–68. [Google Scholar] [CrossRef]
- Ribbens, E.; Anderson, B.A.; Fant, J.B. Opuntia fragilis (Nuttall) Haworth in Illinois: Pad dynamics and sexual reproduction. Haseltonia 2011, 16, 67–78. [Google Scholar] [CrossRef]
- Mandujano, M.C. Reproductive ecology of Opuntia macrocentra (Cactaceae) in the northern Chihuahuan Desert. Am. Midl. Nat. 2013, 169, 274–285. [Google Scholar] [CrossRef]
- De Castro Verçoza, F. Comportamento de Xylocopa ordinaria Smith (Hymenoptera, apidae) na polinização de espécies em duas comunidades vegetais de restinga do município do Rio de Janeiro, Brasil. Revista Dissertar 2019, 1, 27–35. [Google Scholar] [CrossRef]
- Spears, E.E., Jr. Island and mainland pollination ecology of Centrosema virginianum and Opuntia stricta. J. Ecol. 1987, 75, 351–362. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tenorio-Escandón, P.; Ramírez-Hernández, A.; Flores, J.; Juan-Vicedo, J.; Martínez-Falcón, A.P. A Systematic Review on Opuntia (Cactaceae; Opuntioideae) Flower-Visiting Insects in the World with Emphasis on Mexico: Implications for Biodiversity Conservation. Plants 2022, 11, 131. https://doi.org/10.3390/plants11010131
Tenorio-Escandón P, Ramírez-Hernández A, Flores J, Juan-Vicedo J, Martínez-Falcón AP. A Systematic Review on Opuntia (Cactaceae; Opuntioideae) Flower-Visiting Insects in the World with Emphasis on Mexico: Implications for Biodiversity Conservation. Plants. 2022; 11(1):131. https://doi.org/10.3390/plants11010131
Chicago/Turabian StyleTenorio-Escandón, Perla, Alfredo Ramírez-Hernández, Joel Flores, Jorge Juan-Vicedo, and Ana Paola Martínez-Falcón. 2022. "A Systematic Review on Opuntia (Cactaceae; Opuntioideae) Flower-Visiting Insects in the World with Emphasis on Mexico: Implications for Biodiversity Conservation" Plants 11, no. 1: 131. https://doi.org/10.3390/plants11010131