
Genetic and Morpho-Agronomic Characterization of Sicilian Tetraploid Wheat Germplasm

 , , and
, , and
Abstract
:1. Introduction
2. Results
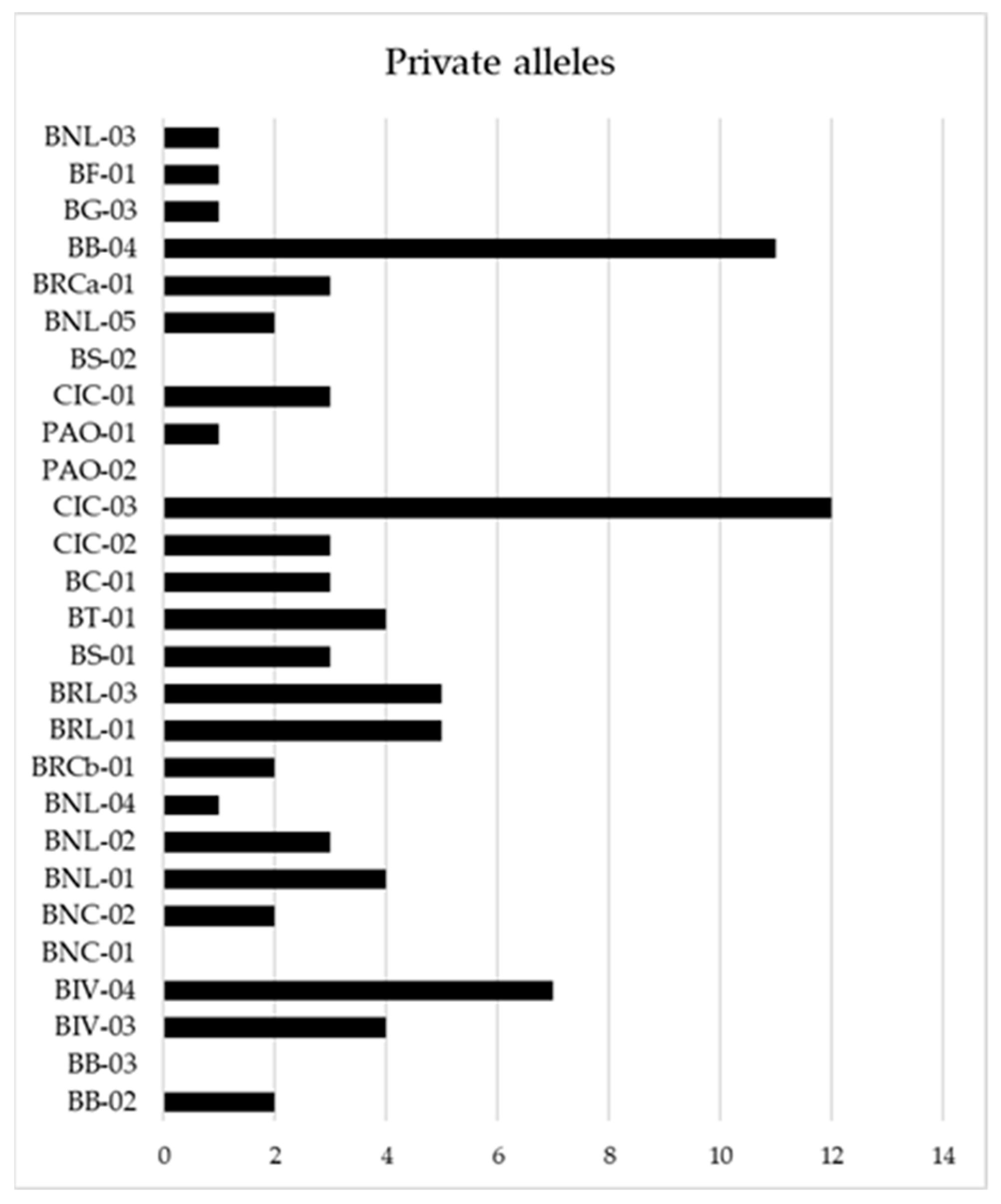
2.1. Genetic Profile
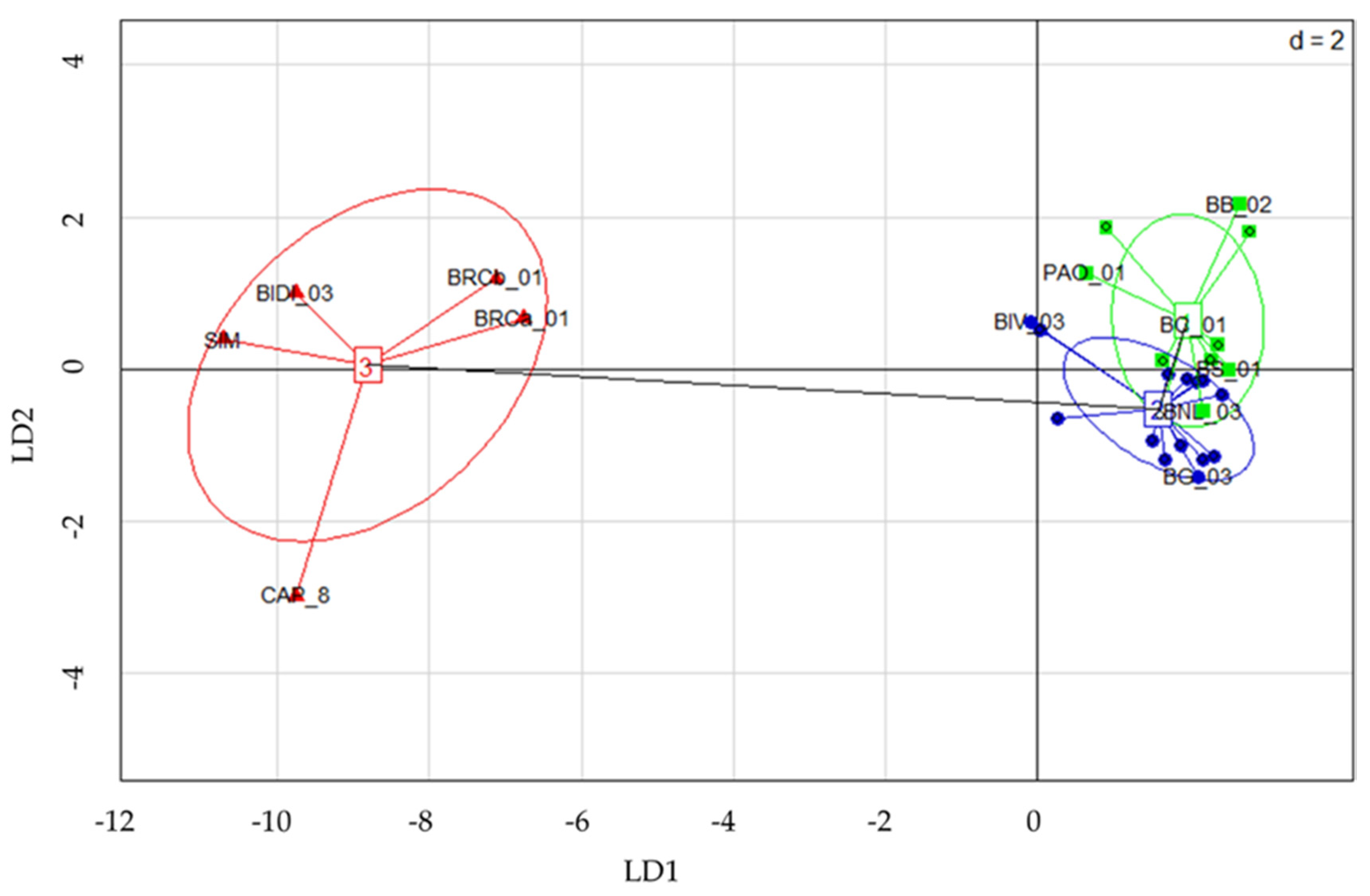
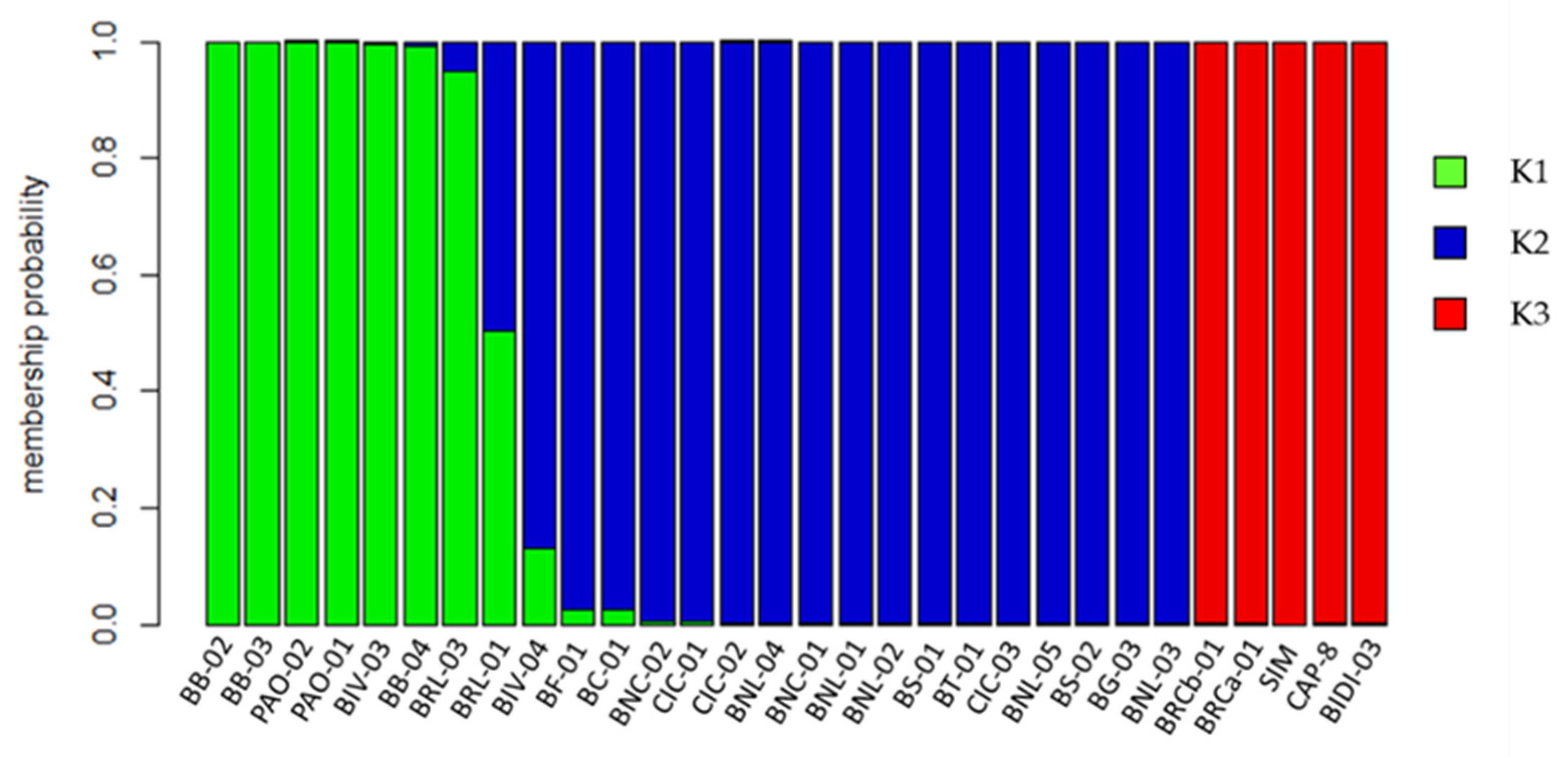
2.2. Genetic Structure
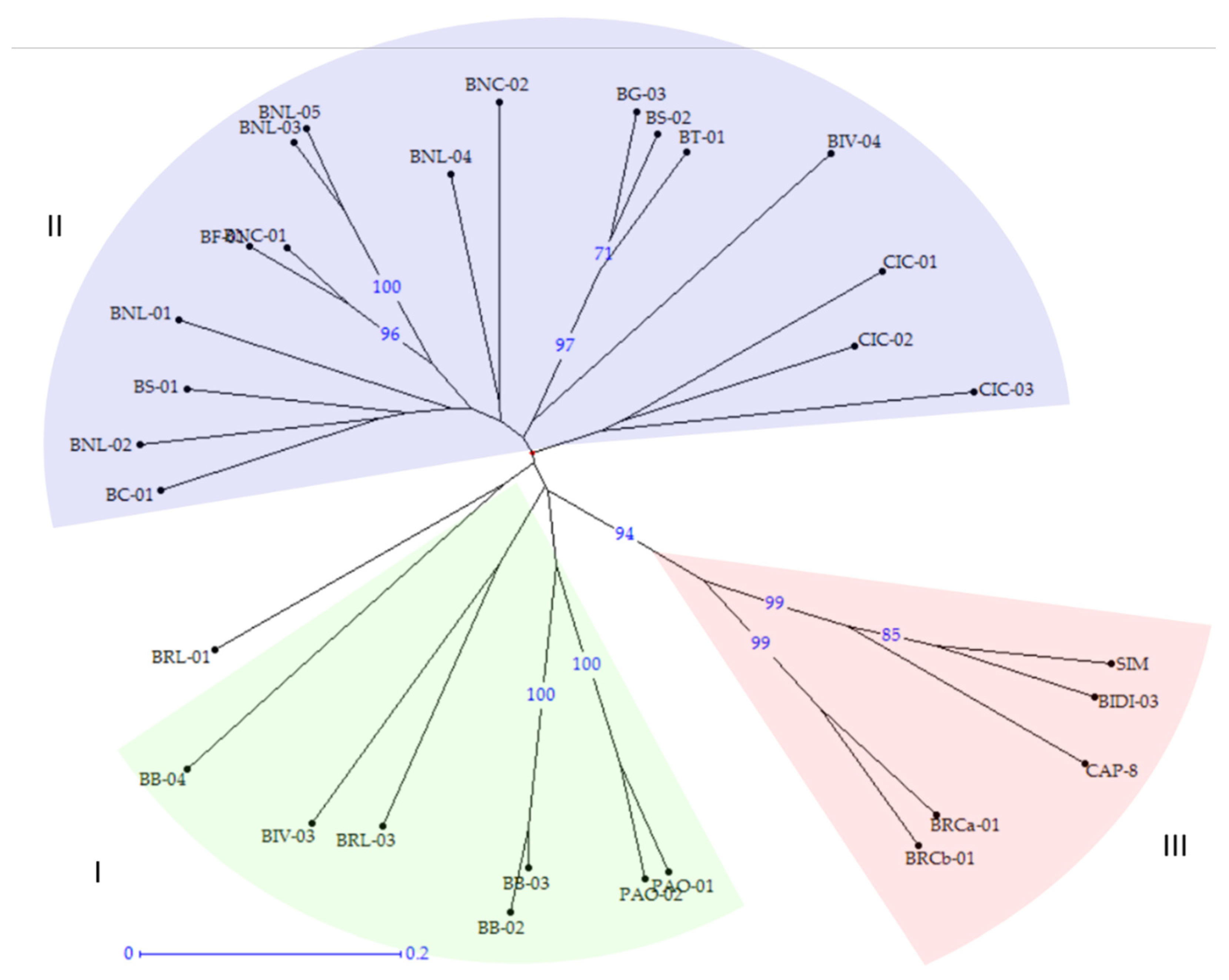
2.3. Phylogenetic Analysis
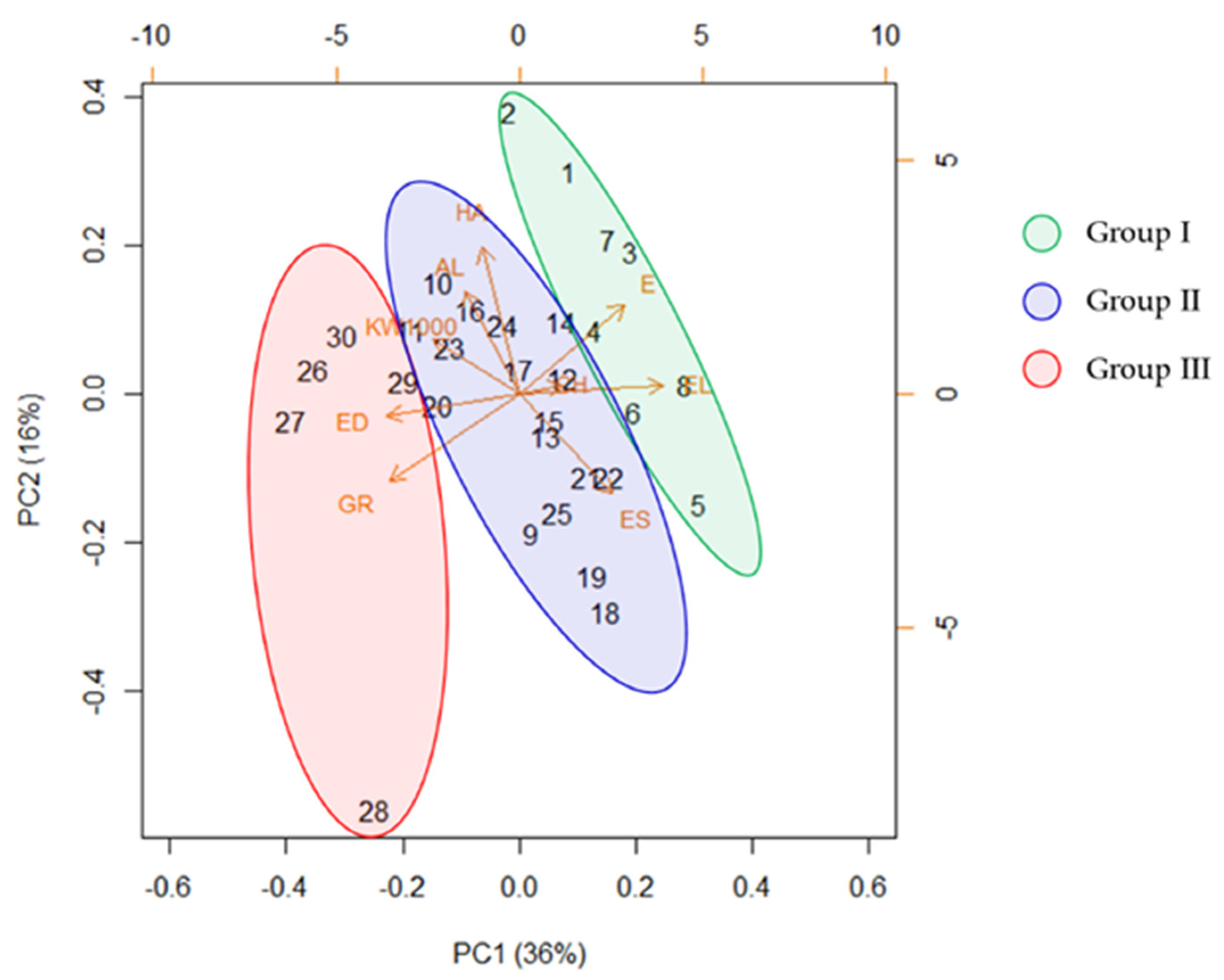
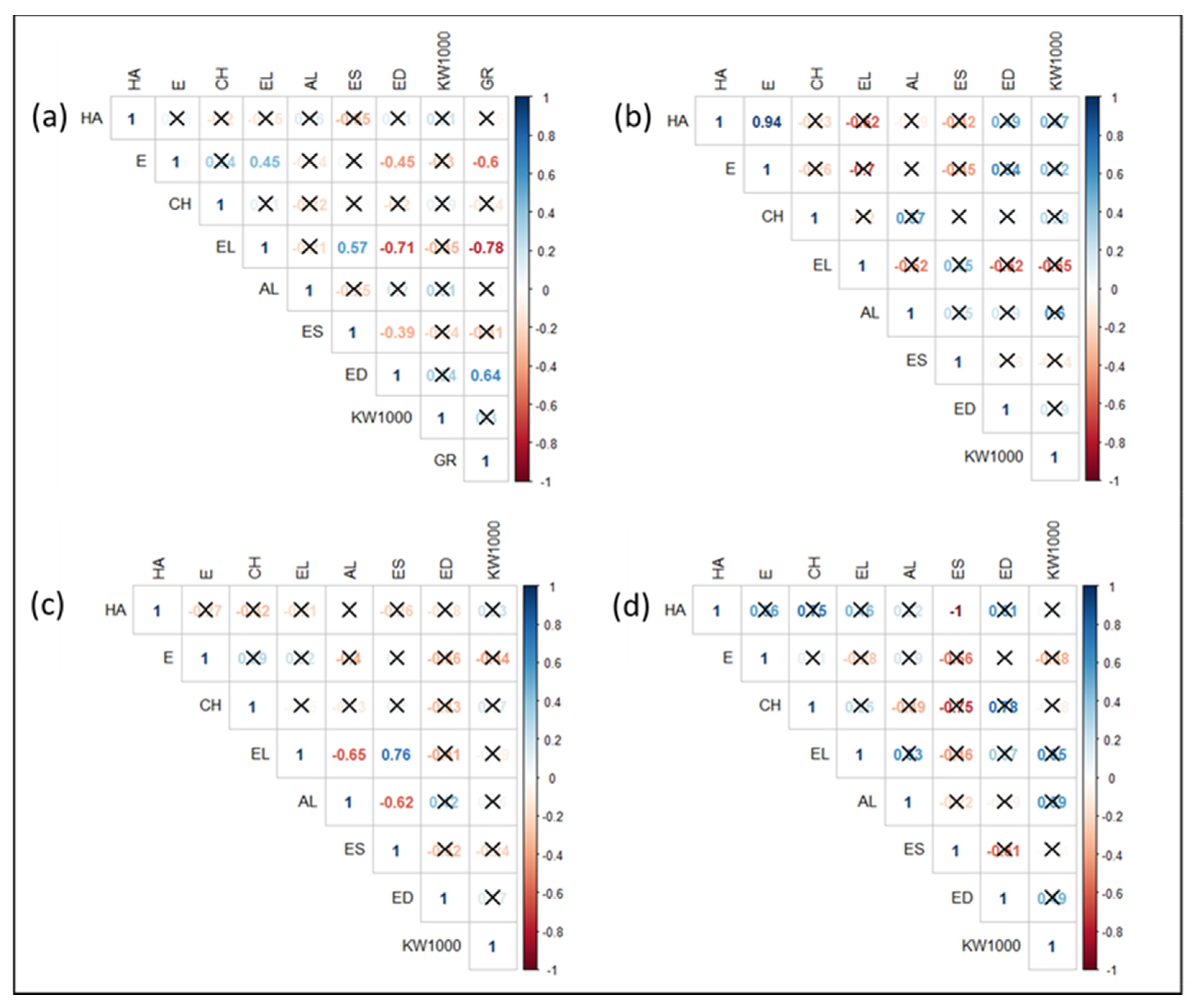
2.4. Morpho-Agronomic Traits
3. Discussion
4. Materials and Methods
4.1. Plant Material and DNA Extraction
4.2. Genotyping by Using SSR Markers
4.3. Statistical Analysis
4.4. Genetic Distance
4.5. Morpho-Agronomic Characterization
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Statement
References
- Villa, T.C.C.; Maxted, N.; Scholten, M.; Ford-Lloyd, B. Defining and identifying crop landraces. Plant Genet. Resour. 2005, 3, 373–384. [Google Scholar] [CrossRef] [Green Version]
- Harlan, J.R. Our vanishing genetic resources. Science 1975, 188, 618–621. [Google Scholar] [CrossRef]
- Dubcovsky, J.; Dvorak, J. Genome plasticity a key factor in the success of polyploid wheat under domestication. Science 2007, 316, 1862–1866. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, H.R.; Campana, M.G.; Jones, H.; Hunt, H.V.; Leigh, F.; Redhouse, D.I.; Lister, D.L.; Jones, M.K. Tetraploid wheat landraces in the Mediterranean basin: Taxonomy, evolution and genetic diversity. PLoS ONE 2012, 7, e37063. [Google Scholar] [CrossRef] [Green Version]
- Asmamaw, M.; Keneni, G.; Tesfaye, K. Genetic Diversity of Ethiopian Durum Wheat (Triticum durum Desf.) Landrace Collections as Reveled by SSR Markers. Adv. Crop. Sci. Technol. 2019, 7, 413. [Google Scholar] [CrossRef]
- Tilman, D. The greening of the green revolution. Nature 1998, 396, 211–212. [Google Scholar] [CrossRef]
- Annicchiarico, P.; Pecetti, L. Yield vs. Morphophysiological trait-based criteria for selection of durum wheat in a semi-arid Mediterranean region (northern Syria). Field Crop. Res. 1998, 59, 163–173. [Google Scholar] [CrossRef]
- Anastasi, U.; Corinzia, S.A.; Cosentino, S.L.; Scordia, D. Performances of Durum Wheat Varieties Under Conventional and No-Chemical Input Management Systems in a Semiarid Mediterranean Environment. Agronomy 2019, 9, 788. [Google Scholar] [CrossRef] [Green Version]
- Newton, A.C.; Akar, T.; Baresel, J.P.; Bebeli, P.J.; Bettencourt, E.; Bladenopoulos, K.V.; Czembor, J.H.; Fasoula, D.A.; Katsiotis, A.; Koutis, K.; et al. Cereal landraces for sustainable agriculture. Sustain. Agric. 2009, 2, 147–186. [Google Scholar] [CrossRef]
- Autrique, E.; Nachit, M.M.; Monneveux, P.; Tanksley, S.D.; Sorrells, M.E. Genetic Diversity in Durum Wheat Based on RFLPs, Morphophysiological Traits, and Coefficient of Parentage. Crop Sci. 1996, 36, 735–742. [Google Scholar] [CrossRef]
- Maccaferri, M.; Sanguineti, M.C.; Donini, P.; Tuberosa, R. Microsatellite analysis reveals a progressive widening of the genetic basis in the elite durum wheat germplasm. Theor. Appl. Genet. 2003, 107, 783–797. [Google Scholar] [CrossRef]
- Fiore, M.C.; Mercati, F.; Spina, A.; Blangiforti, S.; Venora, G.; Dell’Acqua, M.; Lupini, A.; Preiti, G.; Monti, M.; Pè, M.E.; et al. High-Throughput Genotype, Morphology, and Quality Traits Evaluation for the Assessment of Genetic Diversity of Wheat Landraces from Sicily. Plants 2019, 8, 116. [Google Scholar] [CrossRef] [Green Version]
- Kabbaj, H.; Sall, A.T.; Al-Abdallat, A.; Geleta, M.; Amri, A.; Filali-Maltouf, A.; Belkadi, B.; Ortiz, R.; Bassi, F.M. Genetic Diversity within a Global Panel of Durum Wheat (Triticum durum) Landraces and Modern Germplasm Reveals the History of Alleles Exchange. Front. Plant. Sci. 2017, 8, 1277. [Google Scholar] [CrossRef] [Green Version]
- Moragues, M.; Moralejo, M.; Sorrells, M.E.; Royo, C. Dispersal of durum wheat [Triticum turgidum L. ssp. turgidum convar. durum (Desf.) MacKey] landraces across the Mediterranean basin assessed by AFLPs and microsatellites. Genet. Resour. Crop Evol. 2007, 54, 1133–1144. [Google Scholar] [CrossRef]
- Figliuolo, G.; Mazzeo, M.; Greco, I. Temporal variation of diversity in Italian durum wheat germplasm. Genet. Resour. Crop Evol. 2007, 54, 615–626. [Google Scholar] [CrossRef]
- Ruiz, M.; Giraldo, P.; Royo, C.; Villegas, D.; Aranzana, M.J.; Carrillo, J.M. Diversity and Genetic Structure of a Collection of Spanish Durum Wheat Landraces. Crop Sci. 2012, 52, 2262–2275. [Google Scholar] [CrossRef] [Green Version]
- Laidò, G.; Mangini, G.; Taranto, F.; Gadaleta, A.; Blanco, A.; Cattivelli, L.; Marone, D.; Mastrangelo, A.M.; Papa, R.; De Vita, P. Genetic Diversity and Population Structure of Tetraploid Wheats (Triticum turgidum L.) Estimated by SSR, DArT and Pedigree Data. PLoS ONE 2013, 8, e67280. [Google Scholar] [CrossRef] [Green Version]
- Sahri, A.; Chentoufi, L.; Arbaoui, M.; Ardisson, M.; Belqadi, L.; Birouk, A.; Roumet, P.; Muller, M.H. Towards a comprehensive characterization of durum wheat landraces in Moroccan traditional agrosystems: Analysing genetic diversity in the light of geography, farmers’ taxonomy and tetraploid wheat domestication history. BMC Evol. Biol. 2014, 14, 264. [Google Scholar] [CrossRef] [Green Version]
- Röder, M.S.; Korzun, V.; Wendehake, K.; Plaschke, J.; Tixier, M.H.; Leroy, P.; Ganal, M.W. A microsatellite map of wheat. Genetics 1998, 149, 2007–2023. [Google Scholar] [CrossRef]
- Somers, D.J.; Isaac, P.; Edwards, K. A high-density microsatellite consensus map for bread wheat (Triticum aestivum L.). Theor. Appl. Genet. 2004, 109, 1105–1114. [Google Scholar] [CrossRef]
- Paux, E.; Sourdille, P.; Mackay, I.; Feuillet, C. Sequence-based marker development in wheat: Advances and applications to breeding. Biotechnol. Adv. 2012, 30, 1071–1088. [Google Scholar] [CrossRef]
- Motzo, R.; Giunta, F. The effect of breeding on the phenology of Italian durum wheats: From landraces to modern cultivars. Eur. J. Agron. 2007, 26, 462–470. [Google Scholar] [CrossRef]
- De Vita, P.; Nicosia, O.L.D.; Nigro, F.; Platani, C.; Riefolo, C.; Di Fonzo, N.; Cattivelli, L. Breeding progress in morpho-physiological, agronomical and qualitative traits of durum wheat cultivars released in Italy during the 20th century. Eur. J. Agron. 2007, 26, 39–53. [Google Scholar] [CrossRef]
- Hoisington, D.; Khairallah, M.; Reeves, T.; Ribaut, J.M.; Skovmand, B.; Taba, S.; Warburton, M. Plant genetic resources: What can they contribute toward increased crop productivity? Proc. Natl. Acad. Sci. USA 1999, 96, 5937–5943. [Google Scholar] [CrossRef] [Green Version]
- Giunta, F.; Motzo, R.; Pruneddu, G. Trends since 1900 in the yield potential of Italian-bred durum wheat cultivars. Eur. J. Agron. 2007, 27, 12–24. [Google Scholar] [CrossRef]
- Martos, V.; Royo, C.; Rharrabti, Y.; Garcia Del Moral, L.F. Using AFLPs to determine phylogenetic relationships and genetic erosion in durum wheat cultivars released in Italy and Spain throughout the 20th century. F. Crop. Res. 2005, 91, 107–116. [Google Scholar] [CrossRef]
- Marzario, S.; Logozzo, G.; David, J.L.; Zeuli, P.S.; Gioia, T. Molecular genotyping (Ssr) and agronomic phenotyping for utilization of durum wheat (Triticum durum desf.) ex situ collection from southern italy: A combined approach including pedigreed varieties. Genes 2018, 9, 465. [Google Scholar] [CrossRef] [Green Version]
- Maccaferri, M.; Harris, N.S.; Twardziok, S.O.; Pasam, R.K.; Gundlach, H.; Spannagl, M.; Ormanbekova, D.; Lux, T.; Prade, V.M.; Milner, S.G.; et al. Durum wheat genome highlights past domestication signatures and future improvement targets. Nat. Genet. 2019, 51, 885–895. [Google Scholar] [CrossRef] [Green Version]
- Hildebrand, C.E.; Torney, D.C.; Wagner, R.P. Informativeness of Polymorphic DNA Markers. Los Alamos Science 1992, 20, 100–102. [Google Scholar]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software structure: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [Green Version]
- Nei, M. Estimation of average heterozygosity and genetic distance from a small number of individuals. Genetics 1978, 89, 583–590. [Google Scholar] [CrossRef]
- Marone, D.; Laidò, G.; Gadaleta, A.; Colasuonno, P.; Ficco, D.B.M.; Giancaspro, A.; Giove, S.; Panio, G.; Russo, M.A.; De Vita, P.; et al. A high-density consensus map of A and B wheat genomes. Theor. Appl. Genet. 2012, 125, 1619–1638. [Google Scholar] [CrossRef] [Green Version]
- Abbasov, M.; Akparov, Z.; Gross, T.; Babayeva, S.; Izzatullayeva, V.; Hajiyev, E.; Rustamov, K.; Gross, P.; Tekin, M.; Akar, T.; et al. Genetic relationship of diploid wheat (Triticum spp.) species assessed by SSR markers. Genet. Resour. Crop. Evol. 2018, 65, 1441–1453. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research—An update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [Green Version]
- Jombart, T.; Devillard, S.; Balloux, F. Discriminant analysis of principal components: A new method for the analysis of genetically structured populations. BMC Genet. 2010, 11, 94. [Google Scholar] [CrossRef] [Green Version]
- Jombart, T. adegenet: A R package for the multivariate analysis of genetic markers. Bioinformatics 2008, 24, 1403–1405. [Google Scholar] [CrossRef] [Green Version]
- RStudio Team–RStudio. Available online: https://www.rstudio.com/products/team/ (accessed on 13 April 2021).
- Falush, D.; Stephens, M.; Pritchard, J.K. Inference of population structure using multilocus genotype data: Dominant markers and null alleles. Mol. Ecol. Notes 2007, 7, 574–578. [Google Scholar] [CrossRef]
- Earl, D.A.; vonHoldt, B.M. Structure harvester: A website and program for visualizing structure output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef]
- Roussel, V.; Leisova, L.; Exbrayat, F.; Stehno, Z.; Balfourier, F. SSR allelic diversity changes in 480 European bread wheat varieties released from 1840 to 2000. Theor. Appl. Genet. 2005, 111, 162–170. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession | Abbreviation | Origin (Farm) |
|---|---|---|
| Rivet wheat landraces (Bufala and Bufala-related group) | ||
| Bufala Bianca 02 | BB-02 | Randazzo (Catania)—C.da Flascio 1 |
| Bufala Bianca 03 | BB-03 | Gangi (Palermo)—C.da Cavaliere 1 |
| Bufala Bianca 04 | BB-04 | USDA 157984 2 |
| Bufale Cerami 01 | BC-01 | Cerami (Enna) 1 |
| Bufale Flascio 01 | BF-01 | Randazzo (Catania)—C.da Flascio 1 |
| Bufale Gangi 03 | BG-03 | Gangi (Palermo) 3 |
| Bufala Nera Corta 01 | BNC-01 | Maletto (Catania)—C.da Piana 1 |
| Bufala Nera Corta 02 | BNC-02 | IPK 3517 2 |
| Bufala Nera Lunga 01 | BNL-01 | Randazzo (Catania)—C.da Flascio 1 |
| Bufala Nera Lunga 02 | BNL-02 | IPK 4291 2 |
| Bufala Nera Lunga 03 | BNL-03 | Randazzo (Catania)—C.da Flascio 1 |
| Bufala Nera Lunga 04 | BNL-04 | Gangi (Palermo)—C.da Mengarda 1 |
| Bufala Nera Lunga 05 | BNL-05 | Maletto (Catania)—C.da Cimitero 1 |
| Bufala Rossa Corta a 01 | BRCa-01 | Randazzo (Catania)—C.da Flascio 1 |
| Bufala Rossa Corta b 01 | BRCb-01 | Randazzo (Catania)—C.da Flascio 1 |
| Bufala Rossa Lunga 01 | BRL-01 | Randazzo (Catania)—C.da Flascio 1 |
| Bufala Rossa Lunga 03 | BRL-03 | Maletto (Catania)—C.da Piana 1 |
| Bufale Salice 01 | BS-01 | Salice village (Messina) 4 |
| Bufale Salice 02 | BS-02 | Salice village (Messina) 4 |
| Bufale Troina 01 | BT-01 | Troina (Enna) 1 |
| Bivona 03 | BIV-03 | Gangi (Palermo)—C.da Mengarda 1 |
| Bivona 04 | BIV-04 | Santo Stefano Quisquina (Palermo) 1 |
| Ciciredda 01 | CIC-01 | Maletto (Catania)—C.da Piana 1 |
| Ciciredda 02 | CIC-02 | Randazzo (Catania)—C.da Flascio1 |
| Ciciredda 03 | CIC-03 | IPK TRI 28458 2 |
| Paola 01 | PAO-01 | Maletto (Catania)—C.da S. Venera 1 |
| Paola 02 | PAO-02 | Randazzo (Catania)—C.da Flascio 1 |
| Durum wheat improved varieties (Outgroup) | ||
| Bidì 03 | BIDI-03 | IPK TRI 26213 5 |
| Capeiti | CAP-8 | Patended as Capeiti 8 (Eiti 6 × Cappelli) in 1969 at the Stazione Sperimentale di Granicoltura |
| Simeto | SIM | Variety (Capeiti 8 × Valnova) patended in 1988 at Stazione Sperimentale di Granicoltura |
| Genotype Group | N° of Samples | Na | Ho | He | PIC |
|---|---|---|---|---|---|
| Bufala | 20 | 5.56 | 0.35 | 0.55 | 0.50 |
| Bufala-related | 7 | 4.16 | 0.34 | 0.55 | 0.48 |
| Outgroup | 3 | 2.25 | 0.27 | 0.42 | 0.32 |
| Total | 30 | 6.98 | 0.34 | 0.60 | 0.56 |
| Genotype Group | DAPC | ||
|---|---|---|---|
| C1 | C2 | C3 | |
| Bufala | 5 | 13 | 2 |
| Bufala-related | 3 | 4 | 0 |
| Outgroup | 0 | 0 | 3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sicilia, A.; Anastasi, U.; Bizzini, M.; Montemagno, S.; Nicotra, C.; Blangiforti, S.; Spina, A.; Cosentino, S.L.; Lo Piero, A.R. Genetic and Morpho-Agronomic Characterization of Sicilian Tetraploid Wheat Germplasm. Plants 2022, 11, 130. https://doi.org/10.3390/plants11010130
Sicilia A, Anastasi U, Bizzini M, Montemagno S, Nicotra C, Blangiforti S, Spina A, Cosentino SL, Lo Piero AR. Genetic and Morpho-Agronomic Characterization of Sicilian Tetraploid Wheat Germplasm. Plants. 2022; 11(1):130. https://doi.org/10.3390/plants11010130
Chicago/Turabian StyleSicilia, Angelo, Umberto Anastasi, Michele Bizzini, Stefania Montemagno, Carmelo Nicotra, Sebastiano Blangiforti, Alfio Spina, Salvatore Luciano Cosentino, and Angela Roberta Lo Piero. 2022. "Genetic and Morpho-Agronomic Characterization of Sicilian Tetraploid Wheat Germplasm" Plants 11, no. 1: 130. https://doi.org/10.3390/plants11010130