
Mineral Fertilization Influences the Growth, Cryptolepine Yield, and Bioefficacy of Cryptolepis sanguinolenta (Lindl.) Schlt.

 , , ,
, , ,
Abstract
:1. Introduction
2. Results and Discussion
2.1. Effect of Fertilizer Application on Vegetative Growth and Biomass Yield of C. sanguinolenta
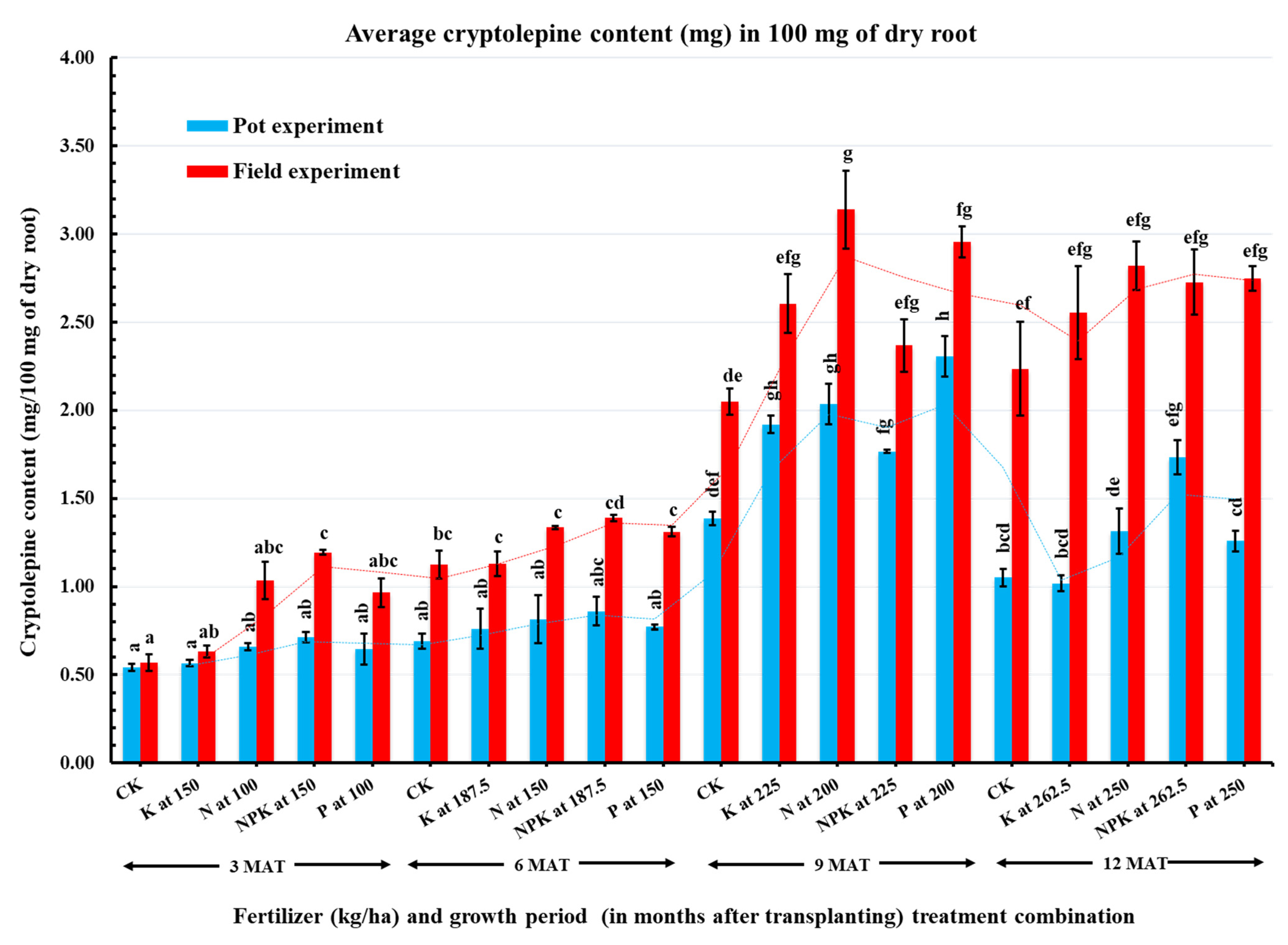
2.2. Effect of Inorganic Fertilizers on Cryptolepine Content
2.3. Effect of Growth Periods on Cryptolepine Content
2.4. Effect of Treatment Combination (Fertilizers and Growth Periods) on Cryptolepine Yield
2.5. Comparison of Variations in Cryptolepine Yield Due to Fertilizer Application and Growth Period
2.6. Effects of Inorganic Fertilizer Application on Soil Fertility
2.7. Effect of Growth Periods on the Bioactivity of C. sanguinolenta Extracts
3. Materials and Methods
3.1. Study Sites
3.2. Transplanting of Seedlings
3.3. Field Experiment
3.4. Pot Experiment
3.5. Plant Growth Measurements, Crop Harvest, and Biomass Analysis
3.6. Soil Analysis
3.7. Estimation of Cryptolepine by High-Performance Liquid Chromatography (HPLC)
Cryptolepine content (mg/100 mg of ground dry roots) × 1000 (mg/g).
3.8. Bioefficacy of Cryptolepine Extracts
3.8.1. Antiplasmodial Activity of C. sanguinolenta Extracts
3.8.2. Cytotoxicity of C. Sanguinolenta Extracts
3.8.3. Anti-Cancer Activity of C. sanguinolenta Extracts
3.9. Data Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cimanga, K.; De Bruyne, T.; Pieters, L.; Totte, J.; Tona, L.; Kambu, K.; Berghe, D.V.; Vlietinck, A. Antibacterial and antifungal activities of neocryptolepine, biscryptolepine and cryptoquindoline, alkaloids isolated from Cryptolepis sanguinolenta. Phytomedicine 1998, 5, 209–214. [Google Scholar] [CrossRef]
- Dwuma-Badu, D.; Ayim, J.S.; Fiagbe, N.I.; Knapp, J.E.; Schiff, P.L., Jr.; Slatkin, D.J. Constituents of West African medicinal plants XX: Quindoline from Cryptolepis sanguinolenta. J. Pharm. Sci. 1978, 67, 433–434. [Google Scholar] [CrossRef]
- Paulo, A.; Gomes, E.T.; Houghton, P.J. New alkaloids from Cryptolepis sanguinolenta. J. Nat. Prod. 1995, 58, 1485–1491. [Google Scholar] [CrossRef]
- Tackie, A.N.; Boye, G.L.; Sharaf, M.H.; Schiff, P.L., Jr.; Crouch, R.C.; Spitzer, T.D.; Johnson, R.L.; Dunn, J.; Minick, D.; Martin, G.E. Cryptospirolepine, a unique spiro-nonacyclic alkaloid isolated from Cryptolepis sanguinolenta. J. Nat. Prod. 1993, 56, 653–670. [Google Scholar] [CrossRef]
- Addy, M. Cryptolepis: An African traditional medicine that provides hope for malaria victims. HerbalGram 2003, 60, 54–59. [Google Scholar]
- Tempesta, M.S. The clinical efficacy of Cryptolepis sanguinolenta in the treatment of malaria. Ghana Med. J. 2010, 44, 1. [Google Scholar] [PubMed]
- Borquaye, L.S.; Gasu, E.N.; Ampomah, G.B.; Kyei, L.K.; Amarh, M.A.; Mensah, C.N.; Nartey, D.; Commodore, M.; Adomako, A.K.; Acheampong, P. Alkaloids from Cryptolepis sanguinolenta as potential inhibitors of SARS-CoV-2 viral proteins: An in silico study. BioMed Res. Int. 2020, 2020, 1–14. [Google Scholar] [CrossRef] [PubMed]
- FDA-Ghana. FDA Approves First Herbal Medicine for the Clinical Trial on COVID-19 Treatment. Food and Drugs Authority Ghana, Accra, Ghana. 2021. Available online: https://fdaghana.gov.gh/press.php?page=18 (accessed on 24 May 2021).
- Amissah, J.N.; Gilday, K. Survey on the Importance of Cryptolepis sanguinolenta in the United States of America. Unpublished work. 2018. [Google Scholar]
- Boakye-Yiadom, K. Antimicrobial Properties of Some West African Medicinal Plants II. Antimicrobial Activity of Aqueous Extracts of Cryptolepis sanguinolenta (Lindl.) Schlechter. Quart. J. Crude Drug Res. 2008, 17, 78–80. [Google Scholar] [CrossRef]
- Ansha, C.; Mensah, K. A review of the anticancer potential of the antimalarial herbal Cryptolepis sanguinolenta and its major alkaloid cryptolepine. Ghana Med. J. 2013, 47, 137–147. [Google Scholar]
- Gudivaka, V.R. Synthesis, Analysis and Biological Evaluation of Novel Indoloquinoline Cryptolepine Analogues as Potential Antitumour Agents. Ph.D. Thesis, Kingston University, London, UK, 2014. [Google Scholar]
- Ameyaw, E.O.; Koffuor, G.A.; Asare, K.K.; Konja, D.; Du-bois, A.; Kyei, S.; Forkuo, A.D.; Mensah, R.N.A.O. Cryptolepine, an indoloquinoline alkaloid, in the management of diabetes mellitus and its associated complications. J. Intercult. Ethnopharmacol. 2016, 5, 263. [Google Scholar] [CrossRef]
- Ablordeppey, S.Y.; Fan, P.; Li, S.; Clark, A.M.; Hufford, C.D. Substituted indoloquinolines as new antifungal agents. Bioorganic Med. Chem. 2002, 10, 1337–1346. [Google Scholar] [CrossRef]
- Bamgbose, S.; Noamesi, B. Studies on cryptolepine. Planta Med. 1981, 41, 392–396. [Google Scholar] [CrossRef] [PubMed]
- Falconer, J. Non-Timber Forest Products in Southern Ghana; Main Report; Chatham Maritime: Chatham, UK, 1994. [Google Scholar]
- Amissah, J.; Osei-Safo, D.; Asare, C.; Missah-Assihene, B.; Danquah, E.; Addae-Mensah, I. Influence of age and staking on the growth and cryptolepine concentration in cultivated roots of Cryptolepis sanguinolenta (Lindl.) Schlt. J. Med. Plants Res. 2016, 10, 113–121. [Google Scholar]
- Monney, M.A.D.; Amissah, N.; Blay, E. Influence of BA and IBA or NAA Combinations on Micropropagation of Cryptolepis sanguinolenta. Am. J. Plant Sci. 2016, 7, 572–580. [Google Scholar] [CrossRef] [Green Version]
- Lubbe, A.; Verpoorte, R. Cultivation of medicinal and aromatic plants for specialty industrial materials. Ind. Crop. Prod. 2011, 34, 785–801. [Google Scholar] [CrossRef]
- Stutte, G.W. Process and product: Recirculating hydroponics and bioactive compounds in a controlled environment. HortScience 2006, 41, 526–530. [Google Scholar] [CrossRef]
- Erisman, J.W.; Sutton, M.A.; Galloway, J.; Klimont, Z.; Winiwarter, W. How a century of ammonia synthesis changed the world. Nat. Geosci. 2008, 1, 636–639. [Google Scholar] [CrossRef]
- Karamanos, A.J.; Sotiropoulou, D.E. Field studies of nitrogen application on Greek oregano (Origanum vulgare ssp. hirtum (Link) Ietswaart) essential oil during two cultivation seasons. Ind. Crop. Prod. 2013, 46, 246–252. [Google Scholar] [CrossRef]
- Nurzyńska-Wierdak, R. Does mineral fertilization modify essential oil content and chemical composition in medicinal plants. Acta Sci. Pol.-Hortorum Cultus 2013, 12, 3–16. [Google Scholar]
- Gholamhosseinpour, Z.; Hemati, K.; Dorodian, H.; Bashiri-Sadr, Z. Effect of nitrogen fertilizer on yield and amount of alkaloids in periwinkle and determination of vinblastine and vincristine by HPLC and TLC. Plant Sci. Res. 2011, 3, 4–9. [Google Scholar]
- Ruminska, A.; El Gamal, E.S. Effect of nitrogen fertilization on growth, yield and alkaloid content in Datura innoxia mill. ActaHortic. 1978, 73, 173–180. [Google Scholar] [CrossRef]
- Kapoor, R.; Giri, B.; Mukerji, K.G. Improved growth and essential oil yield and quality in Foeniculum vulgare mill on mycorrhizal inoculation supplemented with P-fertilizer. Bioresour. Technol. 2004, 93, 307–311. [Google Scholar] [CrossRef] [PubMed]
- Rao, B.R. Biomass and essential oil yields of rainfed palmarosa (Cymbopogon martinii (Roxb.) Wats. var. motia Burk.) supplied with different levels of organic manure and fertilizer nitrogen in semi-arid tropical climate. Ind. Crop. Prod. 2001, 14, 171–178. [Google Scholar] [CrossRef]
- Sifola, M.I.; Barbieri, G. Growth, yield and essential oil content of three cultivars of basil grown under different levels of nitrogen in the field. Sci. Hortic. 2006, 108, 408–413. [Google Scholar] [CrossRef]
- Bloom, A.J.; Frensch, J.; Taylor, A.R. Influence of inorganic nitrogen and pH on the elongation of maize seminal roots. Ann. Bot. 2006, 97, 867–873. [Google Scholar] [CrossRef] [Green Version]
- Bown, H.E.; Watt, M.S.; Clinton, P.W.; Mason, E.G. Influence of ammonium and nitrate supply on growth, dry matter partitioning, N uptake and photosynthetic capacity of Pinus radiata seedlings. Trees 2010, 24, 1097–1107. [Google Scholar] [CrossRef]
- El Gendy, A.; El Gohary, A.; Omer, E.; Hendawy, S.; Hussein, M.; Petrova, V.; Stancheva, I. Effect of nitrogen and potassium fertilizer on herbage and oil yield of chervil plant (Anthriscus cerefolium L.). Ind. Crop. Prod. 2015, 69, 167–174. [Google Scholar] [CrossRef]
- Pandey, V.; Patel, A.; Patra, D. Amelioration of mineral nutrition, productivity, antioxidant activity and aroma profile in marigold (Tagetes minuta L.) with organic and chemical fertilization. Ind. Crop. Prod. 2015, 76, 378–385. [Google Scholar] [CrossRef]
- Waraich, E.A.; Ahmad, Z.; Ahmad, R.; Saifullah; Ashraf, M. Foliar applied phosphorous enhanced growth, chlorophyll contents, gas exchange attributes and PUE in wheat (Triticum aestivum L.). J. Plant Nutr. 2015, 38, 1929–1943. [Google Scholar] [CrossRef]
- Zapata, F.; Zaharah, A. Phosphorus availability from phosphate rock and sewage sludge as influenced by the addition of water soluble phosphate fertilizer. Nutr. Cycl. Agroecosystems 2002, 63, 43–48. [Google Scholar] [CrossRef]
- Barku, V.; Opoku-Boahen, Y.; Dzotsi, E. Isolation and pharmacological activities of alkaloids from Cryptolepis sanguinolenta (Lindl.) schlt. Int. Res. J. Biochem. Bioinform. 2012, 2, 58–61. [Google Scholar]
- Kandil, M.A.M.; Khatab, M.E.; Ahmed, S.S.; Schnug, E. Herbal and essential oil yield of Genovese basil (Ocimum basilicum L.) grown with mineral and organic fertilizer sources in Egypt. J. Für Kult. 2009, 61, 443–449. [Google Scholar]
- Ehsanipour, A.; Razmjoo, J.; Zeinali, H. Effect of nitrogen rates on yield and quality of fennel (Foeniculum vulgare Mill.) accessions. Ind. Crop. Prod. 2012, 35, 121–125. [Google Scholar] [CrossRef]
- Habibi, A.; Heidari, G.; Sohrabi, Y.; Badakhshan, H.; Mohammadi, K. Influence of bio, organic and chemical fertilizers on medicinal pumpkin traits. J. Med. Plants Res. 2011, 5, 5590–5597. [Google Scholar]
- Azizi, A.; Yan, F.; Honermeier, B. Herbage yield, essential oil content and composition of three oregano (Origanum vulgare L.) populations as affected by soil moisture regimes and nitrogen supply. Ind. Crop. Prod. 2009, 29, 554–561. [Google Scholar] [CrossRef]
- Abdelaziz, M.E.; Pokluda, R.; Abdelwahab, M.M. Influence of compost, microorganisms and NPK fertilizer upon growth, chemical composition and essential oil production of Rosmarinus officinalis L. Not. Bot. Horti Agrobot. Cluj-Napoca 2007, 35, 86. [Google Scholar]
- Salisbury, F.; Ross, C. Chap. Lipids and aromatic compounds. In Plant Physiology, 2nd ed.; Wadsworth Publishing Co.: Belmont, CA, USA, 1978. [Google Scholar]
- Hatwar, G.; Gondane, S.; Urkade, S. Effect of micronutrients on growth and yield of chilli. J. Soil Crops 2003, 13, 123–125. [Google Scholar]
- Ramezani, S.; Rezaei, M.R.; Sotoudehnia, P. Improved growth, yield and essential oil content of basil grown under different levels of phosphorus sprays in the field. J. Appl. Biol. Sci. 2009, 3, 105–110. [Google Scholar]
- Jeshni, M.G.; Mousavinik, M.; Khammari, I.; Rahimi, M. The changes of yield and essential oil components of German Chamomile (Matricaria recutita L.) under application of phosphorus and zinc fertilizers and drought stress conditions. J. Saudi Soc. Agric. Sci. 2017, 16, 60–65. [Google Scholar] [CrossRef] [Green Version]
- Chrysargyris, A.; Panayiotou, C.; Tzortzakis, N. Nitrogen and phosphorus levels affected plant growth, essential oil composition and antioxidant status of lavender plant (Lavandula angustifolia Mill.). Ind. Crop. Prod. 2016, 83, 577–586. [Google Scholar] [CrossRef]
- Baskaran, K.; Srinivas, K.; Kulkarni, R. Two induced macro-mutants of periwinkle with enhanced contents of leaf and root alkaloids and their inheritance. Ind. Crop. Prod. 2013, 43, 701–703. [Google Scholar] [CrossRef]
- Siribel, A.; El Hassan, G.; Muddathir, A.; Alla, A. Effect of Season and Plant Age on Growth and Alkaloids Content of Two Strains of Catharanthus roseus LG Don. Grown in Sudan. Khartoum Univ. J. Agric. Sci. 2003, 11, 11–27. [Google Scholar]
- Sreevalli, Y.; Baskaran, K.; Chandrashekara, R.; Kulkarni, R. Preliminary observations on the effect of irrigation frequency and genotypes on yield and alkaloid concentration in periwinkle. J. Med. Aromat. Plant Sci. 2000, 22, 356–358. [Google Scholar]
- Yousaf, M.; Li, J.; Lu, J.; Ren, T.; Cong, R.; Fahad, S.; Li, X. Effects of fertilization on crop production and nutrient-supplying capacity under rice-oilseed rape rotation system. Sci. Rep. 2017, 7, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Dibb, D.; Thompson, W., Jr. Interaction of potassium with other nutrients. Potassium Agric. 1985, 515–533. [Google Scholar]
- Brady, N.C.; Weil, R.R.; Weil, R.R. The Nature and Properties of Soils, 14th ed.; Prentice Hall: Upper Saddle River, NJ, USA, 2008; Volume 13, pp. 662–710. [Google Scholar]
- Ansah, C.; Gooderham, N.J. The popular herbal antimalarial, extract of Cryptolepis sanguinolenta, is potently cytotoxic. Toxicol. Sci. 2002, 70, 245–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laryea, D.; Isaksson, A.; Wright, C.W.; Larsson, R.; Nygren, P. Characterization of the cytotoxic activity of the indoloquinoline alkaloid cryptolepine in human tumour cell lines and primary cultures of tumour cells from patients. Investig. New Drugs 2009, 27, 402–411. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Gooderham, N.J. Mechanisms of induction of cell cycle arrest and cell death by cryptolepine in human lung adenocarcinoma a549 cells. Toxicol. Sci. 2006, 91, 132–139. [Google Scholar] [CrossRef]
- Owusu-Bennoah, E.; Awadzi, T.; Boateng, E.; Krogh, L.; Breuning-Madsen, H.; Borggaard, O.K. Soil properties of a toposequence in the moist semi-deciduous forest zone of Ghana. W. Afr. J. Appl. Ecol. 2000, 1, 1–10. [Google Scholar] [CrossRef]
- Carballas, T.; Macias, F.; Diaz-Fierros, F.; Carballas, M.; Fernandez-Urrutia, J. FAO-UNESCO soil map of the world. Revised legend. Inf. Sobre Recur. Mund. De Suelos (FAO) 1990, 60, 146. [Google Scholar]
- Page, A.; Miller, R.; Keeney, D. Methods of Soil Analysis, Part 2: Chemical and Microbiological Properties, 2nd ed.; American Society of Agronomy: Madison, WI, USA, 1982. [Google Scholar]
- Bouyoucos, G.J. The hydrometer as a new method for the mechanical analysis of soils. Soil Sci. 1927, 23, 343–354. [Google Scholar] [CrossRef]
- Black, C.A. Methods of soil analysis. Part 2. In Chemical and Microbiological Properties; American Society of Agronomy: Madison, WI, USA, 1965. [Google Scholar]
- ISO. ISO 11261: Soil Quality Determination of Total Nitrogen-Modified Kjeldahl Method; International Organization for Standardization: Geneva, Switzerland, 1995. [Google Scholar]
- ISO. ISO 14263: Soil Quality Determination of Phosphorus-Spectrometric Determination of Phosphorus Soluble in Sodium Hydrogen Carbonate Solution; International Standard Organisation: Geneva, Switzerland, 1994. [Google Scholar]
- Chapman, H.; Pratt, P. Methods of analysis for soils, plants and waters. Soil Sci. 1961, 93, 169–176. [Google Scholar] [CrossRef] [Green Version]
- Trager, W.; Jensen, J.B. Human malaria parasites in continuous culture. Science 1976, 193, 673–675. [Google Scholar] [CrossRef] [PubMed]
- Bennett, T.N.; Paguio, M.; Gligorijevic, B.; Seudieu, C.; Kosar, A.D.; Davidson, E.; Roepe, P.D. Novel, rapid, and inexpensive cell-based quantification of antimalarial drug efficacy. Antimicrob. Agents Chemother. 2004, 48, 1807–1810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayisi, N.K.; Appiah-Opong, R.; Gyan, B.; Bugyei, K.; Ekuban, F. Plasmodium falciparum: Assessment of selectivity of action of chloroquine, Alchornea cordifolia, Ficus polita, and other drugs by a tetrazolium-based colorimetric assay. Malar. Res. Treat. 2011, 2011, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Appiah-Opong, R.; Asante, I.K.; Safo, D.O.; Tuffour, I.; Ofori-attah, E.; Uto, T.; Nyarko, A.K. Cytotoxic effects of Albizia zygia (DC) JF Macbr, a Ghanaian medicinal plant, against human T-lymphoblast-like leukemia, prostate and breast cancer cell lines. Int. J. Pharm. Pharm. Sci. 2016, 8, 392–396. [Google Scholar]
- VSN. GenStat Edition 12.1 for Windows. VSN International Ltd.; UK, 2009. Available online: https://www.vsni.co.uk/software/genstat/ (accessed on 27 November 2021).



{kind=link}
{kind=link}
| Treatment | Field Experiment | Pot Experiment | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Fertilizer (kg/ha) | Plant Age (in MAT) | No of Vines | Vine Girth (cm) | No of Vines | Vine Girth (cm) | ||||
| CK | 3 | 2.33 ± 0.33 | a | 2.90 ± 0.35 | abc | 2.33 ± 0.33 | ab | 1.47 ± 0.46 | a |
| K at 150 | 3 | 3.33 ± 0.33 | abc | 3.27 ± 0.29 | bcd | 2.33 ± 0.33 | ab | 1.57 ± 0.37 | a |
| N at 100 | 3 | 3.67 ± 0.33 | abcd | 2.49 ± 0.33 | ab | 3.00 ± 0.58 | ab | 2.07 ± 0.34 | abc |
| NPK at 150 | 3 | 4.33 ± 0.33 | bcd | 3.97 ± 0.17 | cde | 3.67 ± 0.33 | bc | 2.20 ± 0.58 | abc |
| P at 100 | 3 | 3.00 ± 0.12 | ab | 3.23 ± 0.88 | bcd | 3.33 ± 0.33 | abc | 1.63 ± 0.34 | ab |
| CK | 6 | 3.00 ± 0.12 | ab | 2.63 ± 0.36 | abc | 3.67 ± 0.33 | bc | 2.04 ± 0.09 | abc |
| K at 187.5 | 6 | 3.33 ± 0.33 | abc | 3.54 ± 0.58 | bcd | 4.67 ± 0.33 | cd | 3.79 ± 0.58 | de |
| N at 150 | 6 | 4.67 ± 0.33 | bcd | 3.64 ± 0.37 | bcd | 5.33 ± 0.33 | d | 3.73 ± 0.58 | de |
| NPK at 187.5 | 6 | 5.00 ± 0.17 | cd | 4.50 ± 0.66 | de | 7.00 ± 0.58 | e | 3.70 ± 0.58 | de |
| P at 150 | 6 | 3.67 ± 0.33 | abcd | 2.82 ± 0.29 | abc | 6.00 ± 0.58 | de | 2.81 ± 0.58 | abcde |
| CK | 9 | 3.67 ± 0.33 | abcd | 2.70 ± 0.11 | abc | 3.33 ± 0.33 | abc | 2.69 ± 0.46 | abcde |
| K at 225 | 9 | 3.67 ± 0.33 | abcd | 2.63 ± 0.17 | abc | 4.67 ± 0.88 | cd | 2.45 ± 0.65 | abcd |
| N at 200 | 9 | 3.33 ± 0.33 | abc | 1.81 ± 0.27 | a | 3.33 ± 0.33 | abc | 2.67 ± 0.12 | abcde |
| NPK at 225 | 9 | 3.00 ± 0.58 | ab | 2.35 ± 0.23 | ab | 5.33 ± 0.67 | d | 3.88 ± 1.01 | e |
| P at 200 | 9 | 5.33 ± 0.58 | d | 3.23 ± 0.35 | bcd | 3.00 ± 0.25 | ab | 2.06 ± 0.25 | abc |
| CK | 12 | 4.00 ± 1.00 | bcd | 2.58 ± 0.33 | ab | 2.00 ± 0.25 | a | 2.62 ± 0.42 | abcde |
| K at 262.5 | 12 | 4.00 ± 0.58 | bcd | 2.69 ± 0.20 | abc | 3.00 ± 1.00 | ab | 2.80 ± 0.44 | abcde |
| N at 250 | 12 | 4.67 ± 1.20 | cd | 2.62 ± 0.06 | abc | 3.67 ± 0.33 | bc | 3.00 ± 0.43 | bcde |
| NPK at 262.5 | 12 | 5.00 ± 1.53 | cd | 5.15 ± 1.41 | e | 3.67 ± 0.33 | bc | 3.41 ± 0.20 | cde |
| P at 250 | 12 | 5.33 ± 0.88 | d | 2.96 ± 0.12 | abc | 3.33 ± 0.67 | abc | 3.11 ± 0.19 | cde |
| Treatment | Field Experiment | Pot Experiment | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Fertilizer (kg/ha) | Plant Age (in MAT) | Dry Root Biomass (g) | Aboveground Dry Biomass (g) | Shoot/Root Ratio | Dry Root Biomass (g) | Aboveground Dry Biomass (g) | Shoot/Root Ratio | ||||||
| CK | 3 | 6.63 ± 0.88 | ab | 16.10 ± 0.50 | a | 2.53 ± 0.39 | abcd | 3.33 ± 0.55 | a | 4.90 ± 1.15 | a | 1.44 ± 0.11 | ab |
| K at 150 | 3 | 10.87 ± 0.41 | bcde | 21.37 ± 0.91 | ab | 1.98 ± 0.14 | ab | 6.54 ± 1.43 | abc | 9.73 ± 0.88 | ab | 1.68 ± 0.50 | abc |
| N at 100 | 3 | 13.53 ± 0.54 | def | 22.80 ± 1.02 | abc | 1.70 ± 0.14 | a | 11.18 ± 1.85 | defg | 15.57 ± 0.33 | cd | 1.46 ± 0.19 | ab |
| NPK at 150 | 3 | 18.97 ± 0.38 | ghi | 29.57 ± 1.45 | cd | 1.56 ± 0.07 | a | 10.94 ± 1.80 | defg | 25.03 ± 1.15 | ghi | 2.48 ± 0.59 | cde |
| P at 100 | 3 | 6.14 ± 0.29 | a | 21.00 ± 0.40 | ab | 3.43 ± 0.12 | bcd | 6.73 ± 0.33 | abc | 12.07 ± 1.20 | bc | 1.81 ± 0.22 | abcd |
| CK | 6 | 10.60 ± 1.01 | bcd | 27.73 ± 0.60 | bcd | 2.66 ± 0.23 | abcd | 6.07 ± 1.98 | ab | 9.53 ± 1.72 | ab | 1.71 ± 0.21 | abc |
| K at 187.5 | 6 | 12.73 ± 0.19 | cdef | 35.00 ± 1.25 | def | 2.75 ± 0.14 | abcd | 13.37 ± 1.85 | fg | 15.8 ± 0.50 | cd | 1.18 ± 0.11 | a |
| N at 150 | 6 | 16.27 ± 0.29 | fgh | 40.97 ± 0.87 | ef | 2.52 ± 0.01 | abcd | 13.73 ± 0.24 | fg | 25.47 ± 0.39 | ghi | 1.91 ± 0.06 | abcd |
| NPK at 187.5 | 6 | 20.67 ± 0.47 | i | 44.00 ± 0.87 | f | 2.13 ± 0.01 | abc | 14.40 ± 1.73 | g | 41.47 ± 0.94 | k | 2.95 ± 0.29 | e |
| P at 150 | 6 | 12.27 ± 0.70 | cdef | 31.63 ± 0.95 | de | 2.60 ± 0.18 | abcd | 11.40 ± 0.62 | defg | 16.57 ± 0.45 | cde | 1.46 ± 0.04 | ab |
| CK | 9 | 8.56 ± 1.66 | abc | 44.75 ± 4.39 | fg | 5.43 ± 0.52 | f | 6.23 ± 0.50 | ab | 15.85 ± 1.18 | cd | 2.55 ± 0.10 | cde |
| K at 225 | 9 | 11.50 ± 1.80 | cde | 61.11 ± 8.62 | h | 5.44 ± 0.47 | f | 8.39 ± 1.18 | bcde | 19.32 ± 2.38 | def | 2.43 ± 0.13 | cde |
| N at 200 | 9 | 22.40 ± 2.50 | i | 75.88 ± 4.66 | i | 3.45 ± 0.61 | bcd | 10.22 ± 1.39 | cdef | 23.77 ± 2.61 | fgh | 2.42 ± 0.08 | cde |
| NPK at 225 | 9 | 22.68 ± 2.88 | i | 77.67 ± 5.79 | i | 3.52 ± 0.33 | cd | 12.17 ± 1.20 | efg | 25.80 ± 1.18 | ghi | 2.14 ± 0.45 | bcde |
| P at 200 | 9 | 16.07 ± 1.91 | fgh | 60.19 ± 1.45 | h | 3.89 ± 0.81 | de | 8.91 ± 0.97 | bcde | 21.74 ± 1.59 | fg | 2.46 ± 0.58 | cde |
| CK | 12 | 10.45 ± 1.17 | abcd | 56.00 ± 0.40 | gh | 5.57 ± 0.92 | f | 8.19 ± 0.98 | bcd | 21.55 ± 2.23 | efg | 2.66 ± 0.23 | de |
| K at 262.5 | 12 | 12.12 ± 1.63 | cdef | 74.47 ± 8.11 | i | 6.51 ± 0.29 | f | 10.76 ± 1.11 | defg | 23.98 ± 2.82 | fghi | 2.22 ± 0.35 | bcde |
| N at 250 | 12 | 14.98 ± 1.84 | efg | 95.10 ± 5.23 | j | 6.52 ± 0.67 | f | 12.02 ± 1.30 | defg | 28.38 ± 3.33 | hij | 2.46 ± 0.46 | cde |
| NPK at 262.5 | 12 | 21.01 ± 2.77 | i | 111.68 ± 12.24 | j | 5.36 ± 0.73 | ef | 13.48 ± 1.88 | fg | 31.27 ± 2.12 | j | 2.41 ± 0.52 | cde |
| P at 250 | 12 | 19.59 ± 1.44 | hi | 109.47 ± 7.10 | j | 5.68 ± 1.38 | f | 11.12 ± 1.68 | defg | 29.10 ± 2.95 | ij | 2.72 ± 0.06 | de |
| Plant Age at Harvest (in MAT) | Field Experiment | Pot Experiment | ||
|---|---|---|---|---|
| Cryptolepine Content (mg) in 100 mg of Dry Roots | Cryptolepine Yield (mg/plant) | Cryptolepine Content (mg) in 100 mg of Dry Roots | Cryptolepine Yield (mg/plant) | |
| 3 | 0.88 ± 0.14 a | 106.70 ± 33.86 a | 0.63 ± 0.07 a | 49.94 ± 11.98 a |
| 6 | 1.26 ± 0.08 b | 186.00 ± 29.33 a | 0.78 ± 0.12 a | 91.50 ± 18.56 b |
| 9 | 2.63 ± 0.27 c | 431.06 ± 91.36 b | 1.88 ± 0.17 c | 177.10 ± 30.95 c |
| 12 | 2.61 ± 0.28 c | 413.90 ± 81.98 b | 1.28 ± 0.16 b | 146.60 ± 31.44 c |
| mean | 1.84 ± 0.19 | 284.39 ± 42.91 | 1.14 ± 0.12 | 116.26 ± 14.42 |
| Root Biomass | Shoot Biomass | Vine Number | Vine Girth | Cryptolepine Content | Cryptolepine Yield | |
|---|---|---|---|---|---|---|
| Root biomass | 0.78 ** | 0.57 ** | 0.83 ** | 0.16 | 0.53 * | |
| Shoot biomass | 0.61 ** | 0.46 * | 0.71 ** | 0.39 | 0.65 ** | |
| Vine number | 0.53 * | 0.57 ** | 0.65 ** | 0.01 | 0.19 | |
| Vine girth | 0.25 | 0.05 | 0.49 * | 0.25 | 0.54 * | |
| Cryptolepine content | 0.50 * | 0.88 ** | 0.50 * | −0.20 | 0.89 ** | |
| Cryptolepine yield | 0.82 ** | 0.88 ** | 0.50 * | −0.03 | 0.87 ** |
| Source of Variation | Degrees of Freedom | Sum of Squares | Mean Squares | F-Value | p-Value | Variance Component | % of Total |
|---|---|---|---|---|---|---|---|
| Field Experiment | |||||||
| Growth period (GP) | 3 | 1167.58 | 389.19 | 8.53 | 0.001 | 22.90 | 55.67 |
| Block | 2 | 3.44 | 1.72 | 0.37 | 0.696 | 0.00 | 0.00 |
| Fertilizer (GP) | 16 | 730.07 | 45.63 | 9.72 | 0.000 | 13.70 | 33.29 |
| Error | 38 | 178.32 | 4.69 | 4.54 | 11.04 | ||
| Total | 59 | 2079.41 | 41.14 | 100.00 | |||
| Pot Experiment | |||||||
| Growth period (GP) | 3 | 347.20 | 115.73 | 9.26 | 0.001 | 6.76 | 50.63 |
| Fertilizer (GP) | 16 | 200.10 | 12.50 | 3.61 | 0.000 | 3.90 | 29.18 |
| Error | 40 | 138.40 | 3.46 | 2.70 | 20.19 | ||
| Total | 59 | 685.70 | 13.36 | 100.00 | |||
| pH(H2O) | Avail. P (mg/kg) | % N | % OC | Exchangeable Cations (cmol/kg) | Particle Size (%) | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| K+ | Na+ | Ca2+ | Mg2+ | Sand | Silt | Clay | ||||
| 5.50 | 9.72 | 0.09 | 0.60 | 0.14 | 0.70 | 2.60 | 2.00 | 52.90 | 22.10 | 25.00 |
| Treatment | Field Experiment | Pot Experiment | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Fertilizer (kg/ha) | Plant Age (months) | Total N (%) | P (mg/kg) | K (cmol/kg) | Total N (%) | P (mg/kg) | K (cmol/kg) | ||||||
| CK | 3 | 0.84 ± 0.37 | b | 9.62 ± 0.58 | ab | 1.10 ± 0.58 | cd | 1.40 ± 0.20 | b | 17.54 ± 0.58 | a | 1.50 ± 0.33 | b |
| K at 150 | 3 | 1.42 ± 0.33 | c | 29.77 ± 0.66 | f | 3.02 ± 0.33 | f | 1.64 ± 0.39 | b | 16.17 ± 0.58 | a | 1.71 ± 0.33 | b |
| N at 100 | 3 | 1.93 ± 0.42 | d | 16.29 ± 0.61 | bcd | 1.09 ± 0.58 | bcd | 1.68 ± 0.31 | b | 16.36 ± 0.58 | a | 1.46 ± 0.33 | b |
| NPK at 150 | 3 | 1.15 ± 0.58 | cd | 72.50 ± 0.58 | i | 2.12 ± 0.37 | e | 1.43 ± 0.33 | b | 65.28 ± 0.58 | g | 1.69 ± 0.33 | b |
| P at 100 | 3 | 1.50 ± 0.30 | cd | 39.24 ± 0.58 | g | 1.48 ± 0.30 | d | 1.34 ± 0.33 | b | 42.02 ± 0.58 | cd | 1.46 ± 0.33 | b |
| CK | 6 | 0.07 ± 0.00 | a | 29.00 ± 0.73 | ef | 0.10 ± 0.01 | a | 0.07 ± 0.03 | a | 14.43 ± 0.38 | a | 0.13 ± 0.00 | a |
| K at 187.5 | 6 | 0.06 ± 0.00 | a | 19.78 ± 0.85 | cde | 0.50 ± 0.02 | ab | 0.07 ± 0.01 | a | 14.49 ± 1.31 | a | 0.31 ± 0.01 | a |
| N at 150 | 6 | 0.07 ± 0.00 | a | 25.11 ± 0.83 | def | 0.09 ± 0.01 | a | 0.07 ± 0.00 | a | 12.20 ± 0.23 | a | 0.11 ± 0.10 | a |
| NPK at 187.5 | 6 | 0.09 ± 0.00 | a | 54.51 ± 1.06 | h | 0.33 ± 0.02 | a | 0.09 ± 0.00 | a | 56.03 ± 2.63 | ef | 0.19 ± 0.01 | a |
| P at 150 | 6 | 0.06 ± 0.00 | a | 75.13 ± 4.07 | i | 0.10 ± 0.01 | a | 0.06 ± 0.03 | a | 34.90 ± 1.07 | bc | 0.13 ± 0.00 | a |
| CK | 9 | 0.16 ± 0.01 | a | 8.03 ± 0.64 | ab | 0.09 ± 0.03 | a | 0.10 ± 0.01 | a | 17.87 ± 1.34 | a | 0.15 ± 0.01 | a |
| K at 225 | 9 | 0.13 ± 0.01 | a | 6.72 ± 1.93 | a | 0.55 ± 0.05 | abc | 0.09 ± 0.01 | a | 48.80 ± 5.21 | de | 0.20 ± 0.03 | a |
| N at 200 | 9 | 0.12 ± 0.02 | a | 7.78 ± 1.49 | ab | 0.09 ± 0.01 | a | 0.10 ± 0.03 | a | 54.31 ± 5.71 | ef | 0.13 ± 0.01 | a |
| NPK at 225 | 9 | 0.15 ± 0.01 | a | 93.35 ± 1.70 | j | 0.44 ± 0.05 | a | 0.09 ± 0.00 | a | 76.68 ± 3.02 | h | 0.20 ± 0.01 | a |
| P at 200 | 9 | 0.13 ± 0.02 | a | 70.40 ± 12.93 | i | 0.09 ± 0.01 | a | 0.10 ± 0.01 | a | 57.20 ± 5.09 | f | 0.14 ± 0.01 | a |
| CK | 12 | 0.12 ± 0.01 | a | 13.10 ± 1.31 | abc | 0.05 ± 0.01 | a | 0.15 ± 0.01 | a | 11.97 ± 1.06 | a | 0.05 ± 0.03 | a |
| K at 262.5 | 12 | 0.12 ± 0.01 | a | 16.36 ± 1.81 | bcd | 0.35 ± 0.07 | a | 0.15 ± 0.00 | a | 10.55 ± 0.91 | a | 0.15 ± 0.02 | a |
| N at 250 | 12 | 0.12 ± 0.01 | a | 9.32 ± 0.85 | ab | 0.06 ± 0.00 | a | 0.20 ± 0.01 | a | 14.31 ± 1.39 | a | 0.07 ± 0.01 | a |
| NPK at 262.5 | 12 | 0.09 ± 0.01 | a | 43.20 ± 1.12 | g | 0.12 ± 0.01 | a | 0.16 ± 0.01 | a | 51.53 ± 3.77 | ef | 0.08 ± 0.03 | a |
| P at 250 | 12 | 0.10 ± 0.00 | a | 20.12 ± 1.72 | cde | 0.07 ± 0.01 | a | 0.16 ± 0.03 | a | 31.15 ± 3.17 | b | 0.06 ± 0.01 | a |
| Growth Period (in MAT) | Antiplasmodial Activity | Antiplasmodial Selectivity Index (SI) | Anti-Cancer Activity (Cell Viability) | |||
|---|---|---|---|---|---|---|
| IC50 Values for Pot Expt. (µg/mL) | IC50 Values for Field Expt. (µg/mL) | SI for Pot Expt. (*CC50/IC50) | SI for Field Expt. (*CC50/IC50) | CC50 Values for Pot Expt. (µg/mL) | CC50 Values for Field Expt. (µg/mL) | |
| 3 | 9.23 ± 0.05 | 8.12 ± 0.07 | 1.08 | 1.23 | 6.83 ± 1.03 | 12.25 ± 1.41 |
| 6 | 7.37 ± 0.06 | 6.44 ± 0.56 | 1.36 | 1.55 | 30.91 ± 1.95 | 8.81 ± 0.76 |
| 9 | 6.30 ± 0.30 | 4.65 ± 0.02 | 1.59 | 2.15 | 11.7 ± 2.52 | 8.68 ± 0.46 |
| 12 | 2.56 ± 0.04 | 5.06 ± 1.28 | 3.91 | 1.98 | – | 62.53 ± 6.68 |
| Control | 3.01 ± 0.03 x | 3.01 ± 0.03 x | 0.05 | 0.05 | 8.56 ± 1.67 y | 8.56 ± 1.67 y |
| Fertilizer | Fertilizer Rates (kg/ha) Used in Each Growth Period (Months after Transplanting) | |||
|---|---|---|---|---|
| 3 Months 2* | 6 Months 4 | 9 Months 6 | 12 Months 8 | |
| Urea (N) | 100.0 | 150.0 | 200.0 | 250.0 |
| Triple Super Phosphate (P) | 100.0 | 150.0 | 200.0 | 250.0 |
| Muriate of Potash (K) | 150.0 | 187.5 | 225.0 | 262.5 |
| NPK | 150.0 | 187.5 | 225.0 | 262.5 |
| Control (CK) | 0.0 | 0.0 | 0.0 | 0.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amissah, J.N.; Alorvor, F.E.; Okorley, B.A.; Asare, C.M.; Osei-Safo, D.; Appiah-Opong, R.; Addae-Mensah, I. Mineral Fertilization Influences the Growth, Cryptolepine Yield, and Bioefficacy of Cryptolepis sanguinolenta (Lindl.) Schlt. Plants 2022, 11, 122. https://doi.org/10.3390/plants11010122
Amissah JN, Alorvor FE, Okorley BA, Asare CM, Osei-Safo D, Appiah-Opong R, Addae-Mensah I. Mineral Fertilization Influences the Growth, Cryptolepine Yield, and Bioefficacy of Cryptolepis sanguinolenta (Lindl.) Schlt. Plants. 2022; 11(1):122. https://doi.org/10.3390/plants11010122
Chicago/Turabian StyleAmissah, Jacqueline Naalamle, Forgive Enyonam Alorvor, Benjamin Azu Okorley, Chris Mpere Asare, Dorcas Osei-Safo, Regina Appiah-Opong, and Ivan Addae-Mensah. 2022. "Mineral Fertilization Influences the Growth, Cryptolepine Yield, and Bioefficacy of Cryptolepis sanguinolenta (Lindl.) Schlt." Plants 11, no. 1: 122. https://doi.org/10.3390/plants11010122