
Non-Target Site Mechanisms Endow Resistance to Glyphosate in Saltmarsh Aster (Aster squamatus)

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
2.1. Resistance Confirmation to Glyphosate
2.2. Acummulation of Shikimic Acid
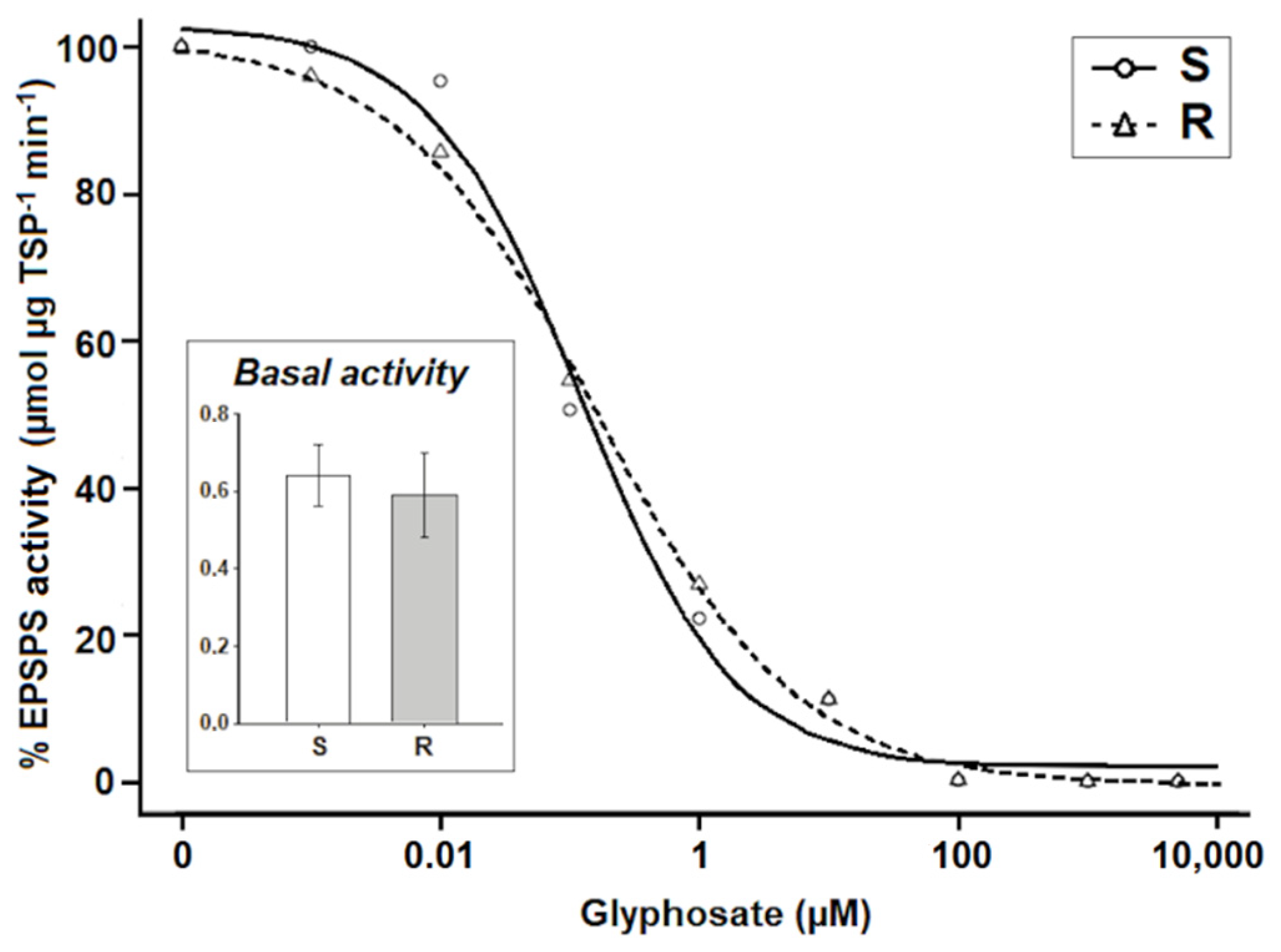
2.3. Concentration and Activity of the EPSPS
2.4. Absorption and Translocation of 14C-Glyphosate
2.5. Glyphosate Metabolism
3. Discussion
4. Materials and Methods
4.1. Biological Material
4.2. Dose-Response Curves
4.3. Shikimic Acid Accumulation
4.4. Enzymatic Activity of EPSPS
4.5. 14C-Glyphosate Absorption and Translocation
4.6. Glyphosate Metabolism
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Antier, C.; Kudsk, P.; Reboud, X.; Ulber, L.; Baret, P.; Messéan, A. Glyphosate use in the European agricultural sector and a framework for its further monitoring. Sustainability 2020, 12, 5682. [Google Scholar] [CrossRef]
- Gravena, R.; Filho, R.V.; Alves, P.L.C.A.; Mazzafera, P.; Gravena, A.R. Glyphosate has low toxicity to citrus plants growing in the field. Can. J. Plant Sci. 2012, 92, 119–127. [Google Scholar] [CrossRef]
- Woodburn, A.T. Glyphosate: Production, pricing and use worldwide. Pest Manag. Sci. 2000, 56, 309–312. [Google Scholar] [CrossRef]
- Alcántara-de la Cruz, R.; Domínguez-Martínez, P.A.; Silveira, H.M.; Cruz-Hipólito, H.E.; Palma-Bautista, C.; Vázquez-García, J.G.; Domínguez-Valenzuela, J.A.; De Prado, R. Management of glyphosate-resistant weeds in Mexican citrus groves: Chemical alternatives and economic viability. Plants 2019, 8, 325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- SADER-SIAP. Anuario Estadístico De La Producción Agrícola. 2019. Available online: https://nube.siap.gob.mx/cierreagricola/ (accessed on 26 February 2021).
- FAOSTAT. Crops. 2021. Available online: http://www.fao.org/faostat/en/?#data/QC (accessed on 26 February 2020).
- USDA—U.S. Department of Agriculture. Citrus: World Markets and Trade. 2021. Available online: https://apps.fas.usda.gov/psdonline/circulars/citrus.pdf (accessed on 1 April 2021).
- Pérez-López, M.; González-Torralva, F.; Cruz-Hipólito, H.; Santos, F.; Domínguez-Valenzuela, J.A.; De Prado, R. Characterization of glyphosate-resistant tropical sprangletop (Leptochloa virgata) and its alternative chemical control in persian lime orchards in Mexico. Weed Sci. 2014, 62, 441–450. [Google Scholar] [CrossRef]
- Alcántara-de la Cruz, R.; Cruz-Hipolito, H.E.; Domínguez-Valenzuela, J.A.; De Prado, R. Glyphosate ban in Mexico: Potential impacts on agriculture and weed management. Pest Manag. Sci. 2021, 77, 3820–3831. [Google Scholar] [CrossRef] [PubMed]
- Troiani, H.O.; Steibel, P.E. Synopsis of the compositae giseke of La Pampa, Argentina. Rev. Fac. Agron. 1998, 9, 1–86. [Google Scholar]
- CABI. Symphyotrichum Subulatum Var. Squamatum. 2019. Available online: https://www.cabi.org/cpc/datasheet/120910 (accessed on 2 February 2021).
- Nesom, G.L. Taxonomy of the Symphyotrichum (Aster) subulatum group and Symphyotrichum (Aster) tenuifolium (Asteraceae: Astereae). SIDA Contrib. Bot. 2005, 21, 2125–2140. [Google Scholar]
- Heap, I. The International Survey of Herbicide Resistant Weeds. 2021. Available online: www.weedscience.org (accessed on 10 February 2021).
- Gaines, T.A.; Patterson, E.L.; Neve, P. Molecular mechanisms of adaptive evolution revealed by global selection for glyphosate resistance. New Phytol. 2019, 223, 1770–1775. [Google Scholar] [CrossRef] [Green Version]
- Dayan, F.E.; Barker, A.; Takano, H.; Bough, R.; Ortiz, M.; Duke, S.O. Herbicide mechanisms of action and resistance. In Comprensive Biotechnology Vol. 4, 3rd ed.; Moo-Young M, Ed.; Pergamon: Oxford, UK, 2019; pp. 36–48. [Google Scholar]
- Gaines, T.A.; Duke, S.O.; Morran, S.; Rigon, C.A.G.; Tranel, P.J.; Küpper, A.; Dayan, F.E. Mechanisms of evolved herbicide resistance. J. Biol. Chem. 2020, 295, 10307–10330. [Google Scholar] [CrossRef]
- Osuna, M.D.; Fischer, A.J.; De Prado, R. Herbicide resistance in Aster squamatus conferred by a less sensitive form of acetolactate synthase. Pest Manag. Sci. 2003, 59, 1210–1216. [Google Scholar] [CrossRef]
- Duke, S.O. Enhanced metabolic degradation: The last evolved glyphosate resistance mechanism of weeds? Plant Physiol. 2019, 181, 1401–1403. [Google Scholar] [CrossRef] [Green Version]
- La Cruz, R.A.-D.; Fernández-Moreno, P.T.; Ozuna, C.V.; Rojano-Delgado, A.M.; Cruz-Hipolito, H.E.; Valenzuela, J.A.D.; Barro, F.; De Prado, R. Target and non-target site mechanisms developed by glyphosate-resistant hairy beggarticks (Bidens pilosa L.) populations from Mexico. Front. Plant Sci. 2016, 7, 1492. [Google Scholar]
- Palma-Bautista, C.; Gherekhloo, J.; Domínguez-Martínez, P.A.; Domínguez-Valenzuela, A.; Cruz-Hipolito, H.E.; Alcántara-de la Cruz, R.; Rojano-Delgado, A.M.; De Prado, R. Characterization of three glyphosate resistant Parthenium hysterophorus populations collected in citrus groves from Mexico. Pestic. Biochem. Physiol. 2019, 155, 1–7. [Google Scholar] [CrossRef]
- Moretti, M.L.; Van Horn, C.R.; Robertson, R.; Segobye, K.; Weller, S.C.; Young, B.G.; Johnson, W.G.; Sammons, R.D.; Wang, D.; Ge, X.; et al. Glyphosate resistance in Ambrosia trifida: Part 2. Rapid response physiology and non-target-site resistance. Pest Manag. Sci. 2018, 74, 1079–1088. [Google Scholar] [CrossRef]
- Van Horn, C.R.; Moretti, M.L.; Robertson, R.R.; Segobye, K.; Weller, S.C.; Young, B.G.; Johnson, W.; Schulz, B.; Green, A.C.; Jeffery, T.; et al. Glyphosate resistance in Ambrosia trifida: Part 1. Novel rapid cell death response to glyphosate. Pest Manag. Sci. 2018, 74, 1071–1078. [Google Scholar] [CrossRef]
- De Queiroz, A.R.S.; Delatorre, C.A.; Lucio, F.R.; Rossi, C.V.S.; Zobiole, L.H.S.; Merotto, A. Rapid necrosis: A novel plant resistance mechanism to 2,4-D. Weed Sci. 2020, 68, 6–18. [Google Scholar] [CrossRef]
- Shaner, D.L.; Nadler-Hassar, T.; Henry, W.B.; Koger, C.H. A rapid in vivo shikimate accumulation assay with excised leaf discs. Weed Sci. 2005, 53, 769–774. [Google Scholar] [CrossRef]
- Maeda, H.; Dudareva, N. The shikimate pathway and aromatic amino acid biosynthesis in plants. Annu. Rev. Plant Biol. 2012, 63, 73–105. [Google Scholar] [CrossRef] [PubMed]
- Bracamonte, E.; Silveira, H.M.; Alcántara-de la Cruz, R.; Domínguez-Valenzuela, J.A.; Cruz-Hipolito, H.E.; De Prado, R. From tolerance to resistance: Mechanisms governing the differential response to glyphosate in Chloris barbata. Pest Manag Sci. 2018, 74, 1118–1124. [Google Scholar] [CrossRef] [PubMed]
- Dayan, F.E.; Owens, D.K.; Corniani, N.; Silva, F.M.L.; Watson, S.B.; Howell, J.; Shaner, D.L. Biochemical markers and enzyme assays for herbicide mode of action and resistance studies. Weed Sci. 2015, 63, 23–63. [Google Scholar] [CrossRef] [Green Version]
- Cruz-Hipolito, H.; Rojanodelgado, A.M.; Valenzuela, J.A.D.; Heredia, A.; de Castro, M.D.L.; De Prado, R. Glyphosate tolerance by Clitoria ternatea and Neonotonia wightii plants involves differential absorption and translocation of the herbicide. Plant Soil 2011, 347, 221–230. [Google Scholar] [CrossRef]
- Menendez, J.; Rojano-Delgado, M.A.; De Prado, R. Differences in herbicide uptake, translocation, and distribution as sources of herbicide resistance in weeds. In Retention, Uptake, and Translocation of Agrochemicals in Plants, 1st ed.; Myung, K., Satchivi, N.M., Kingston, C.K., Eds.; ACS Publications: Washington, DC, USA, 2014; pp. 141–157. [Google Scholar]
- Nandula, V.K.; Ray, J.D.; Ribeiro, D.N.; Pan, Z.; Reddy, K.N. Glyphosate resistance in tall waterhemp (Amaranthus tuberculatus) from Mississippi is due to both altered target-site and nontarget-site mechanisms. Weed Sci. 2013, 61, 374–383. [Google Scholar] [CrossRef]
- Michitte, P.; De Prado, R.; Espinoza, N.; Ruiz-Santaella, J.P.; Gauvrit, C. Mechanisms of resistance to glyphosate in a ryegrass (Lolium multiflorum) biotype from Chile. Weed Sci. 2007, 55, 435–440. [Google Scholar] [CrossRef]
- Vila-Aiub, M.M.; Balbi, M.C.; Distéfano, A.J.; Fernandez, L.; Hopp, H.E.; Yu, Q.; Powles, S.B. Glyphosate resistance in perennial Sorghum halepense (Johnsongrass), endowed by reduced glyphosate translocation and leaf uptake. Pest Manag. Sci. 2012, 68, 430–436. [Google Scholar] [CrossRef] [PubMed]
- Svyantek, A.W.; Aldahir, P.; Chen, S.; Flessner, M.L.; McCullough, P.E.; Sidhu, S.S.; McElroy, J.S.; Aldahir, P. Target and nontarget resistance mechanisms induce annual bluegrass (Poa annua) resistance to atrazine, amicarbazone, and diuron. Weed Technol. 2016, 30, 773–782. [Google Scholar] [CrossRef]
- De La Cruz, R.A.; Amaral, G.D.S.; Mendes, K.F.; Rojano-Delgado, A.M.; De Prado, R.; Silva, M.F.D.G.F.D. Absorption, translocation, and metabolism studies of herbicides in weeds and crops. In Radioisotopes in Weed Research, 1st ed.; Mendes, K.F., Ed.; CRC Press: Boca Raton, FL, USA, 2021; pp. 127–154. [Google Scholar]
- Ge, X.; d’Avignon, D.A.; Ackerman, J.J.H.; Sammons, D.R. Rapid vacuolar sequestration: The horseweed glyphosate resistance mechanism. Pest Manag. Sci. 2010, 66, 345–348. [Google Scholar] [CrossRef] [Green Version]
- Kleinman, Z.; Rubin, B. Non-target-site glyphosate resistance in Conyza bonariensis is based on modified subcellular distribution of the herbicide. Pest Manag. Sci. 2017, 73, 246–253. [Google Scholar] [CrossRef]
- Palma-Bautista, C.; Torra, J.; Garcia, M.J.; Bracamonte, E.; Rojano-Delgado, A.M.; la Cruz, R.A.-D.; De Prado, R. Reduced absorption and impaired translocation endows glyphosate resistance in Amaranthus palmeri harvested in glyphosate-resistant soybean from Argentina. J. Agric. Food Chem. 2019, 67, 1052–1060. [Google Scholar] [CrossRef] [Green Version]
- Ghanizadeh, H.; Harrington, K.C. Restricted glyphosate translocation in Lolium multiflorum is controlled by a single incomplete dominant nuclear gene. N. Z. J. Crop Hortic. Sci. 2018, 46, 346–353. [Google Scholar] [CrossRef]
- La Cruz, R.A.-D.; Rojano-Delgado, A.M.; Giménez, M.J.; Cruz-Hipolito, H.E.; Domínguez-Valenzuela, J.A.; Barro, F.; De Prado, R. First resistance mechanisms characterization in glyphosate-resistant Leptochloa virgata. Front. Plant Sci. 2016, 7, 1742. [Google Scholar]
- Gherekhloo, J.; Fernández-Moreno, P.T.; La Cruz, R.A.-D.; Sánchez-González, E.; Cruz-Hipolito, H.E.; Domínguez-Valenzuela, J.A.; De Prado, R. Pro-106-Ser mutation and EPSPS overexpression acting together simultaneously in glyphosate-resistant goosegrass (Eleusine indica). Sci. Rep. 2017, 7, 6702. [Google Scholar] [CrossRef] [PubMed]
- De Carvalho, L.B.; Alves, P.L.D.C.A.; González-Torralva, F.; Cruz-Hipolito, H.E.; Rojano-Delgado, A.M.; De Prado, R.; Gil Humanes, J.; Barro, F.; De Castro, M.D.L. Pool of resistance mechanisms to glyphosate in Digitaria insularis. J. Agric. Food Chem. 2012, 60, 615–622. [Google Scholar] [CrossRef]
- Duke, S.O. Glyphosate degradation in glyphosate-resistant and -susceptible crops and weeds. J. Agric. Food Chem. 2011, 59, 5835–5841. [Google Scholar] [CrossRef] [PubMed]
- Pan, L.; Yu, Q.; Han, H.; Mao, L.; Nyporko, A.; Fan, L.; Bai, L.; Powles, S. Aldo-keto reductase metabolizes glyphosate and confers glyphosate resistance in Echinochloa colona. Plant Physiol. 2019, 181, 1519–1534. [Google Scholar] [CrossRef] [PubMed]
- McElroy, J.S.; Hall, N.D. Echinochloa colona with reported resistance to glyphosate conferred by aldo-keto reductase also contains a pro-106-Thr EPSPS target site mutation. Plant Physiol. 2020, 183, 447–450. [Google Scholar] [CrossRef] [Green Version]
- Baek, Y.; Bobadilla, L.K.; Giacomini, D.A.; Montgomery, J.S.; Murphy, B.P.; Tranel, P.J. Evolution of glyphosate-resistant weeds. In Reviews of Environmental Contamination and Toxicology; Springer: New York, NY, USA, 2021; pp. 1–36. [Google Scholar] [CrossRef]
- Palma-Bautista, C.; Tahmasebi, B.K.; Fernández-Moreno, P.T.; Rojano-Delgado, A.M.; Alcántara de la Cruz, R.; De Prado, R. First Case of Conyza canadensis from Hungary with Multiple Resistance to Glyphosate and Flazasulfuron. Agronomy 2018, 8, 157. [Google Scholar] [CrossRef] [Green Version]
- Yanniccari, M.; Vázquez-García, J.G.; Gómez-Lobato, M.E.; Rojano-Delgado, A.M.; Alves, P.L.D.C.A.; De Prado, R. First case of glyphosate resistance in Bromus catharticus Vahl.: Examination of endowing resistance mechanisms. Front. Plant Sci. 2021, 12, 617945. [Google Scholar] [CrossRef]
- Rojano-Delgado, A.M.; Ruiz-Jiménez, J.; De Castro, M.D.L.; De Prado, R. Determination of glyphosate and its metabolites in plant material by reversed-polarity CE with indirect absorptiometric detection. Electrophoresis 2010, 31, 1423–1430. [Google Scholar] [CrossRef]
- Ritz, C.; Baty, F.; Streibig, J.C.; Gerhard, D. Dose-response analysis using R. PLoS ONE 2015, 10, e0146021. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dose-Response | Population | b | d | g | RF |
|---|---|---|---|---|---|
| Weight reduction (g ae ha−1) | S | 1.2 | 99.9 | 14.7 ± 1.2 | 13.7 |
| R | 1.3 | 101.1 | 202.1 ± 17.2 | ||
| Plant mortality (g ae ha−1) | S | 5.5 | 100.0 | 214.3 ± 11.4 | 9.7 |
| R | 7.1 | 99.9 | 2078.1 ± 60.9 | ||
| Enzyme inhibition (µM) | S | 0.2 | 99.6 | 0.27 ± 0.04 | 0.85 |
| R | 4.2 | 100.5 | 0.23 ± 0.02 |
| HAT | Population | Glyphosate | AMPA * | Glyoxylate |
|---|---|---|---|---|
| 48 | S | 53.3 ± 4.2 (87) a | 4.4 ± 0.8 (7) b | 3.9 ± 0.6 (6) b |
| R | 45.5 ± 3.8 (65) b | 12.6 ± 2.0 (18) a | 11.7 ± 1.9 (17) a | |
| 72 | S | 57.7 ± 4.7 (73) a | 11.2 ± 1.4 (14) b | 10.1 ± 1.2 (13) b |
| R | 43.6 ± 3.6 (50) b | 22.3 ± 2.2 (26) a | 21.1 ± 3.1 (24) a | |
| 96 | S | 59.8 ± 4.6 (64) a | 18.1 ± 1.8 (19) b | 16.3 ± 2.3 (17) b |
| R | 30.1 ± 2.6 (28) b | 37.3 ± 4.1 (35) a | 39.8 ± 4.3 (37) a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Domínguez-Valenzuela, J.A.; Alcántara-de la Cruz, R.; Palma-Bautista, C.; Vázquez-García, J.G.; Cruz-Hipolito, H.E.; De Prado, R. Non-Target Site Mechanisms Endow Resistance to Glyphosate in Saltmarsh Aster (Aster squamatus). Plants 2021, 10, 1970. https://doi.org/10.3390/plants10091970
Domínguez-Valenzuela JA, Alcántara-de la Cruz R, Palma-Bautista C, Vázquez-García JG, Cruz-Hipolito HE, De Prado R. Non-Target Site Mechanisms Endow Resistance to Glyphosate in Saltmarsh Aster (Aster squamatus). Plants. 2021; 10(9):1970. https://doi.org/10.3390/plants10091970
Chicago/Turabian StyleDomínguez-Valenzuela, José Alfredo, Ricardo Alcántara-de la Cruz, Candelario Palma-Bautista, José Guadalupe Vázquez-García, Hugo E. Cruz-Hipolito, and Rafael De Prado. 2021. "Non-Target Site Mechanisms Endow Resistance to Glyphosate in Saltmarsh Aster (Aster squamatus)" Plants 10, no. 9: 1970. https://doi.org/10.3390/plants10091970