1. Introduction
The entire Himalayan region is a repository of high value medicinal plants and is therefore, considered as a global biodiversity hotspot. Out of 18,440 plant species found in the Himalayan region, 8298 species are medicinally important [
1,
2].
Berberis lycium is one among them. Commonly known as Indian barberry, it is an evergreen shrub belonging to family Berberidaceae. It is a sub-erect branched shrub growing to a height of 3–4 m. It is native to Nepal and is abundantly distributed in the Himalayas extending through West of Pakistan, North and Central India, Afghanistan and the entirety of Nepal [
3]. Within India, it is frequently distributed in Jammu and Kashmir, Uttar Pradesh, Madhya Pradesh, Himachal Pradesh, Uttrakhand, Sikkim and Nilgiri hills of Tamil Nadu within altitudinal gradients of 850–3500 m.a.s.l. [
3,
4,
5]. This plant is used in polyherbal formulation of organized medicine such as Ayurveda, Siddha and Unani [
6]. Pharmacological investigation of the entire plant body parts has revealed the presence of different secondary metabolites (alkaloids, tannins, saponins, flavonoids etc.) which signifies its potential role in the formulation of antibacterial, antifungal, antidiabetic and cardio-protective drugs [
7,
8]. Active constituents include berberine [
9,
10], an isoquinoline alkaloid which is widely known for its activity against severe diarrhea [
11], cholera [
12], latent malaria and amoebiasis [
12]. Owing to its wide spectrum of medicinal significance this plant is being exploited at an alarming rate. Besides, it is being eradicated from the Himalayan region to reclaim the hill slopes for agriculture or to extract a valuable drug berberidine from its roots and stem [
13,
14]. Decline in the population of this species urges the need for its timely conservation so as to prevent this plant from getting critically endangered or extinct [
15].
To design an effective conservation strategy for any species of plants, information about its reproductive behavior and pollination ecology along with other threatening factors becomes imperative [
16,
17]. Scientific data generated in such studies help in designing conservation strategies on one hand and in exploring the mechanism by which it has become endangered on the other [
18]. In this context, study of the reproductive biology of the plants, particularly endangered and overexploited assumes a paramount significance. Reproductive biology allows us to understand the different phenophases of a species and processes beginning from the production of gametes to the germination of the seeds and constrains that limit the multiplication of the species in nature [
19,
20]. Studies on these aspects of reproductive biology of threatened taxa have provided valuable information regarding the limitation in the process of sexual reproduction and have helped in planning effective conservation and management programs [
20,
21,
22,
23,
24,
25]. Mating and the breeding systems of plant species have found to be detrimental in designing the genetic structure of the populations. Pollination biology is an important parameter in the life history of flowering plants that determines the success of sexual reproduction [
26,
27,
28]. Effective conservation and management programmes based on the result of such studies have been executed for a number of threatened species [
29,
30,
31,
32,
33]. Such studies bring to light the bottlenecks, if any, on account of which the individuals of the species are not able to reproduce in nature [
34,
35]. It also determines the genetic makeup of the populations, the magnitude of variations and therefore, ultimately helps in planning appropriate conservation strategies for the species under different categories of threat [
36,
37].
Berberis lycium Royle is one of the heavily exploited and threatened medicinal plants of the Himalayan region. Every part of the plant is used for medicinal purposes [
38]. The species has been declared endangered in the North-West Himalayan region by FRLHT as per IUCN criteria of 2000 (envis.frlht.org/frlhtenvis.nic.in). Non-availability of basic biological data regarding phenology, reproductive biology, and pollination ecology of
Berberis lycium are a major concern and are detrimental to its recovery plans. To fill these gaps, the present study was conducted to explore and gather the information on
B. lycium with respect to above mentioned parameters, to understand this plant species at its various phenophases and to elucidate potential deficiencies in its reproduction and pollination mechanisms. This study helped to understand the effect of different biotic and abiotic constraints on the phenology and reproductive biology of the plant, including some unique mechanisms displayed by the flower to facilitate pollination and to ensure reproduction. Such a study will not only help to achieve improvement in the existing status of this species but also assist in unravelling the critical events which occur during the life cycle of the species and in turn help with planning strategies for effective conservation and management. The information generated thereof will enable conservation biologists to design appropriate strategies for long-term survival and sustenance of
B. lycium.
3. Discussion
Occurrence of variants of a species in an area indicates the diversification of the species. All the three variants of
Berberis lycium observed in the study area are characterized by different foliar characters. They differ with respect to size, shape and texture of leaves, inflorescence length, number of flowers per inflorescence, diameter of flowers and number of floral parts. Earlier, Ahrendt [
39] recognized four variants of
Berberis lycium of which three have been reported from Kashmir. The plants of variants I and II enter the flowering phase in the months of February and April, respectively, and the flowering period lasts for just one month whereas the plants of variant III enter the reproductive phase in September and continue producing flowers till January. Variation in flowering time is often considered with a genetic component and has been demonstrated in many studies by different workers [
40,
41,
42,
43,
44].
For the successful initiation of pollen pistil interaction, pollen viability and stigma receptivity are important parameters [
45,
46,
47,
48]. Presence of certain enzymes such as acid phosphatases, peroxidases and esterases mark the receptivity of stigmas [
49,
50]. Extended stigma receptivity assures pollination, fertilization and reproductive success in plants [
51]. The stigma remains receptive for greater periods in
B. lycium to provide proper conditions for pollen to germinate and affect fertilization. This time frame appears to be an approach to ensure reproductive output in this plant species. Mechanism of anther dehiscence is unique and similar in all the three variants. Anther dehiscence occurs simultaneously with anthesis. The large pollen sacs of dehiscing lobes of alternate anthers detach themselves from the filament remaining attached at the top of the connective part and then move in such a way that pollen laden sacs face the stigmatic surface to affect pollination. This pattern of anther dehiscence is believed to be associated with the size difference between the dorsal and ventral side of pollen sacs in each theca, the ventral sacs being much smaller than the dorsal one. The stomium stretches longitudinally between the two pollen sacs and around the base and upward along the dorsal side of the theca which permits the wall of dorsal pollen sac to curve upward at dehiscence [
52]. Alternate anther dehiscence appears to be an adaptation for efficient and extended pollen availability to the pistil.
Like animals, plants also respond to mechanical stimuli, although the signaling machinery is poorly understood [
53]. Flowers and leaves are more commonly responsive to external mechanical stimuli among plant organs [
53]. The first report of thigmo-response of stamens dates long back when Linnaeus described the sensitive stamens of
Berberis vulgaris flowers in his 1755 version of the
Flora Suecica. Stamen movement determines the fate of pollen transfer with pollinators [
54]. Staminal movement is normally correlated with cross-pollination in plants [
55] though it also promotes self-pollination [
36] as has been observed in
B. lycium. A characteristic feature of sensitive stamens is the presence of papillate structures in the sensitive areas which are believed to be related to the water mobility after being irritated and, thus, inducing the thigmonastic response [
56]. In
B. lycium, only the basal part of stamens is sensitive, papillae are localized only at the base of staminal filament.
The floral morphology of different plant species determines their pollination behavior [
57,
58]. Different floral traits such as fewer terminal leaf panicle inflorescences, small perianth, exposed stigmatic surface and raised anthers, delicate fragrance, nectar availability, and dense and showy inflorescences favor insect pollination [
58,
59]. Our observations propose that the pollination syndrome of
B. lycium is entomophilous; i.e., it is pollinated by insects. In all the three variants, pollination is dependent on insect visit. When the visiting insect inserts its proboscis to suck nectar secreted by nectaries at the base of corolla lobes, it invariably touches the base of staminal filaments and induces their movement, thus pushing them towards the interior of flower in such a way that the dehisced anther lobes deposit pollen on the receptive stigmatic surface. The insects with strong proboscis can induce the staminal movement. Morphotype II attract diverse numbers of pollinators as compared to other two morphotypes which is attributed to greater length of inflorescences and large number of flowers. Floral secretions, colorful and attractive inflorescences are more imperative in attracting the pollinators than aroma [
60,
61]. The color of flowers could also be considered as an advertisement of reward when the meaning of flowers is to attract pollinators. Bright yellow petals of
B. lycium have evolved as a means for attraction of pollinators. Pollinators prefer warmer afternoon hours to visit the flowers after which their visitation frequency decreased. Such findings draw their support from the reports of [
42]. Different plant species receive a drastic decrease in the visitation efficiency of pollinators during wind, cloudy periods and rain [
62,
63]. Adverse climatic conditions were found to hamper the visitation frequency and efficiency of the pollinators.
The mechanisms of anther dehiscence, staminal movement and pollination syndrome appear to be an adaptation to affect the self-pollination in
B. lycium. It is further strengthened by low fruit set in flowers whose self-pollen is foraged by predatory larvae. Results of pollination experiments in
B. lycium point towards the mixed breeding strategy followed by the species to ensure reproductive output. This kind of strategy not only ensures sexual reproduction but also generates variation and distributes it effectively [
64]. Low fruit set in variants I and III in open pollination can be due to non-availability of pollen grains to receptive stigmata. Both pollinator and pollen limitation appear to be the reasons for low reproductive output in open pollination in variants I and III. Seed set was high in variants I and II. However, the seeds are predated by larvae of some insects in both cases. Variant I is more susceptible to predation leading to 10% loss in reproductive output. Majority of fruits of variant III were seedless and those with seeds get infected and damaged. Seeds in such fruits thus turn nonviable leading to low reproductive output. Low reproductive output of this variant appears to be the reason for its low representation in the study area.
The percentage seed germination varied greatly on wet filter paper and in soil conditions. Seeds fail to germinate and emerge out of the soil when sown in soil. Low germination of seeds sown in soil can be attributed to non availability of light to the germinating seeds. Many workers have reported this in different weed species [
65,
66,
67]. It has been reported that very little light is reached to the seeds below a depth of 4mm in all soil types [
68]. Increase in the storage period and aging of seeds results in decreased vigor and germination index [
69]. Decrease in the germination percentage with respect to their increased storage period reflects the decreased longevity of the seeds in
B. lycium. These results are in agreement with the findings of Arif [
70] who reported inverse relation between seed longevity and storage period. The differences in the germination percentage of seeds may be due to decline in phospholipids and polyunsaturated fatty acids [
71]. Our results are equally supported by the findings of Kandil [
71] and Muhammadi [
72] who also reported significant decrease in the germination percentage of seeds with respect to increased storage periods.
4. Materials and Methods
4.1. Phenology, Floral Organization, Traits and Reproductive Biology
Mature and healthy plants in full bloom were randomly selected and tagged to study floral traits and biology. A detailed study regarding different quantitative characteristics of floral parts, viz., length and width of the flower, porophylls size, length of pistil, length of stamen, was carried out under a stereo zoom microscope (Nikon- BX51). The number of flowers per inflorescence, number of anthers per flower, length of inflorescence was noted directly in the field with applied standard deviations.
Data on different aspects of reproductive biology like plant and floral morphology, phenology, pollen viability, pollen–ovule ratio, anther dehiscence and pollination mechanism, pollinator behavior, pollen–pistil interaction, reproductive output and seed germination were collected for the plant under study.
4.2. Floral Biology
Data on flower opening (anthesis) were collected, both in nature and under lab conditions. Time and pattern of anther dehiscence was determined by periodic examination of anthers. Stigma receptivity was determined by manually pollinating the stigmata and examining them for pollen germination at different time intervals either directly or after fixing them in Carnoy’s fixative (three parts absolute alcohol and one part glacial acetic acid) and then shifting them to 70% alcohol. Pistils were then stained in Lewis’ stain (mixture of 2mL of aqueous acid fuschin, 2mL of 1% aq. Lightgreen, 40 mL of lactic acid and 46 mL distilled water) [
38] for microscopic studies. Some of the pistils were stained with Aniline blue for fluorescence microscopy [
73].
4.3. Pollen–Ovule Ratio
Pollen output was calculated by first counting the number of pollen grains per anther and multiplying this figure by the number of stamens per flower. The ovule count was determined by putting the ovaries in 4N NaOH for 12–14 h at 60 °C in a hot air oven, washing them thoroughly in water to remove every trace of NaOH and then gently squashing the ovaries in a drop of Lewis stain on a slide and then counting ovules by observing these under the microscope. The amount of pollen divided by number of ovules per flower gave pollen–ovule ratio. Pollen–ovule ratio was calculated following the method of Kumari [
18].
4.4. Pollen Stainability and Viability
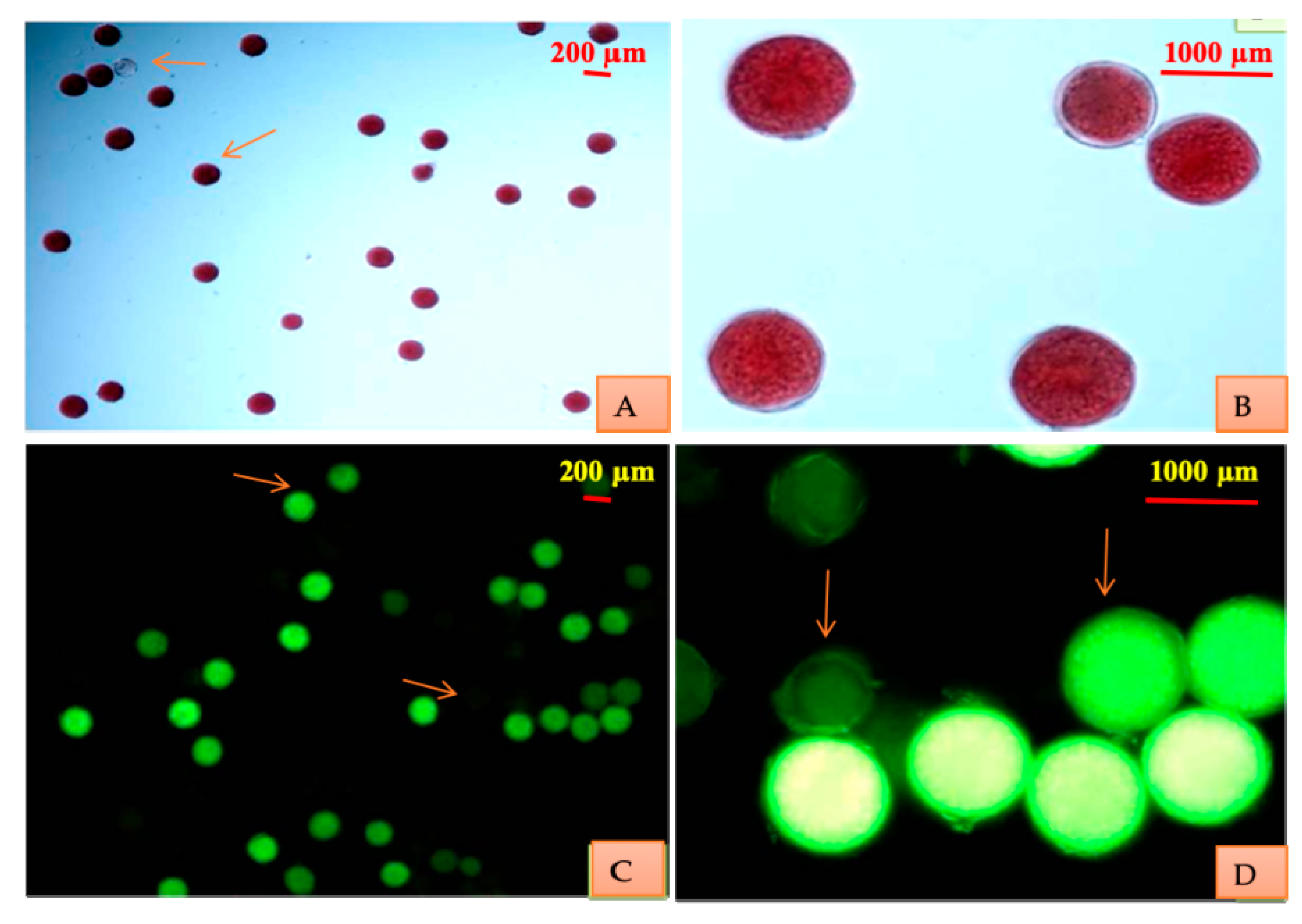
Pollen stainability and viability was checked by conducting a stainability test in 1% acetocarmine and FCR (a mixture of FDA in acetone and sucrose) test, respectively. The concentration of sucrose used was 10%. In the stainability test, pollen with stained cytoplasm were considered viable while shriveled and unstained were taken as nonviable. In the FCR test, pollen with fluorescent and non-fluorescent cytoplasm were treated as viable and nonviable, respectively.
4.5. Pollen–Pistil Interaction
Pollen–pistil interaction was studied by following the Aniline blue fluorescence method [
18]. Pistils were cleared by keeping them in 4N NaOH at 60 °C for 3 h. They were then stained in 0.005% aniline blue (discolor) for 30 min. Later on, they were mounted in 1:1
v/v mixture of aniline blue and glycerin and observed under microscope (Nikon 80 i Eclipse (Nikon, Tokyo, Japan)). Under fluorescence microscope, germination of pollen on the stigma and path of pollen tubes through the stylar tissue to the ovule was studied.
4.6. Pollination Mechanism
To understand the pollination mechanism, different structural features of the flower like type of corolla, position of the reproductive parts in a fully opened flower and kind of pollen and stigma were studied carefully. To check anemophily, slides smeared with Mayer’s albumin were suspended at varying distances from plants. These slides were then examined after 24 h by staining in Lewis stain for presence or absence of the pollen of the plant under study. To check entomophily as the possible mode of pollination, observations were made whether or not the flowers are visited by the insects. The rewards offered by the flowers to the visiting insects were also noted. Behavior of the insects visiting the flowers was monitored to determine how they affect pollination.
Different pollination indices were calculated for all the pollinators to determine their contribution to pollen transfer. The foraging behavior (FB) of insect visitors was ascertained through direct field visitations at regular intervals of time. It was determined as the time spent by a particular pollinator per inflorescence per visit [
19,
74]. Foraging speed (FS) was calculated as the average number of flowers visited per minute of time [
19,
75]. Insect visitation efficiency (IVE) and insect visitation frequency (IVF) were determined following the methodology put forward by Yaqoob and Nawchoo [
19].
4.7. Mating or Breeding System
Different pollination experiments were carried out to determine the nature of breeding system. Inflorescences were subjected to forced selfing by bagging them. Compatibility status was determined by manual pollinations (manual self and crosspollination). In any pollination experiment, fruit set was taken as the end product of effective pollination, thereby confirming the compatibility. In order to ascertain breeding behavior for B. lycium the following experiments were placed:
Control
Flowers were kept undisturbed and monitored for fruit set.
Agamospermy or apomixis
During bud condition flowers were emasculated and bagged to check fruit set.
Xenogamy
Flowers were emasculated, followed by manual cross pollination and bagging.
Autonomous selfing or unassisted selfing
Flowers were bagged during bud condition and monitored regularly for fruit set.
Facilitated autogamy (selfing)
Plants were emasculated, followed by manual self-pollination and bagging.
Geitonogamy
Stigmas of the flowers were pollinated with pollen from different flowers of the same plant.
4.8. Reproductive Output
For calculating reproductive output, percent fruit set per plant and percent seed set per fruit were calculated. Both these aspects were studied for open pollination in nature. To determine percent fruit set, first the average number of flowers per plant and then average number of fruits formed per plant was determined. Percentage fruit set was calculated using the following formula:
For determining percentage seed set, first the average no. of ovules per ovary and then average number of seeds per fruit was calculated. Percentage seed set was determined using the following formula:
4.9. Seed Germination
The potential of the seeds to germinate was determined by keeping the seeds for germination under lab conditions and in substrata like moist silver paper and mixture of sand, manure and garden soil in the ratio of 1:1:1. Total number of germinated seeds by total number of seeds kept for germination yielded the percentage seed germination.
4.10. Data Analysis Using Statistical Software
The data analysis was performed using R software v.4.0.3 (R Core Team, 2021). We performed one-way ANOVA (analysis of variance) test to study whether the studied variants differed in terms of their morphological characters, pollen biology and floral anthesis. Two-way ANOVA test was carried out to determine the simultaneous effect of different variants and treatment on fruit and seed development. Redundancy analysis (RDA) was carried out to study the similarity between pollinators in terms of their pollination indices separately using the Biodiversity R package. Additionally, we searched for the most contributing factors, which explained 40% or more of variation in the foraging behavior and pollen pollinator efficiency, respectively, and thus were most effective in separating different pollinators.


 ,
, 










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}