
Photorhabdus spp.: An Overview of the Beneficial Aspects of Mutualistic Bacteria of Insecticidal Nematodes


Abstract
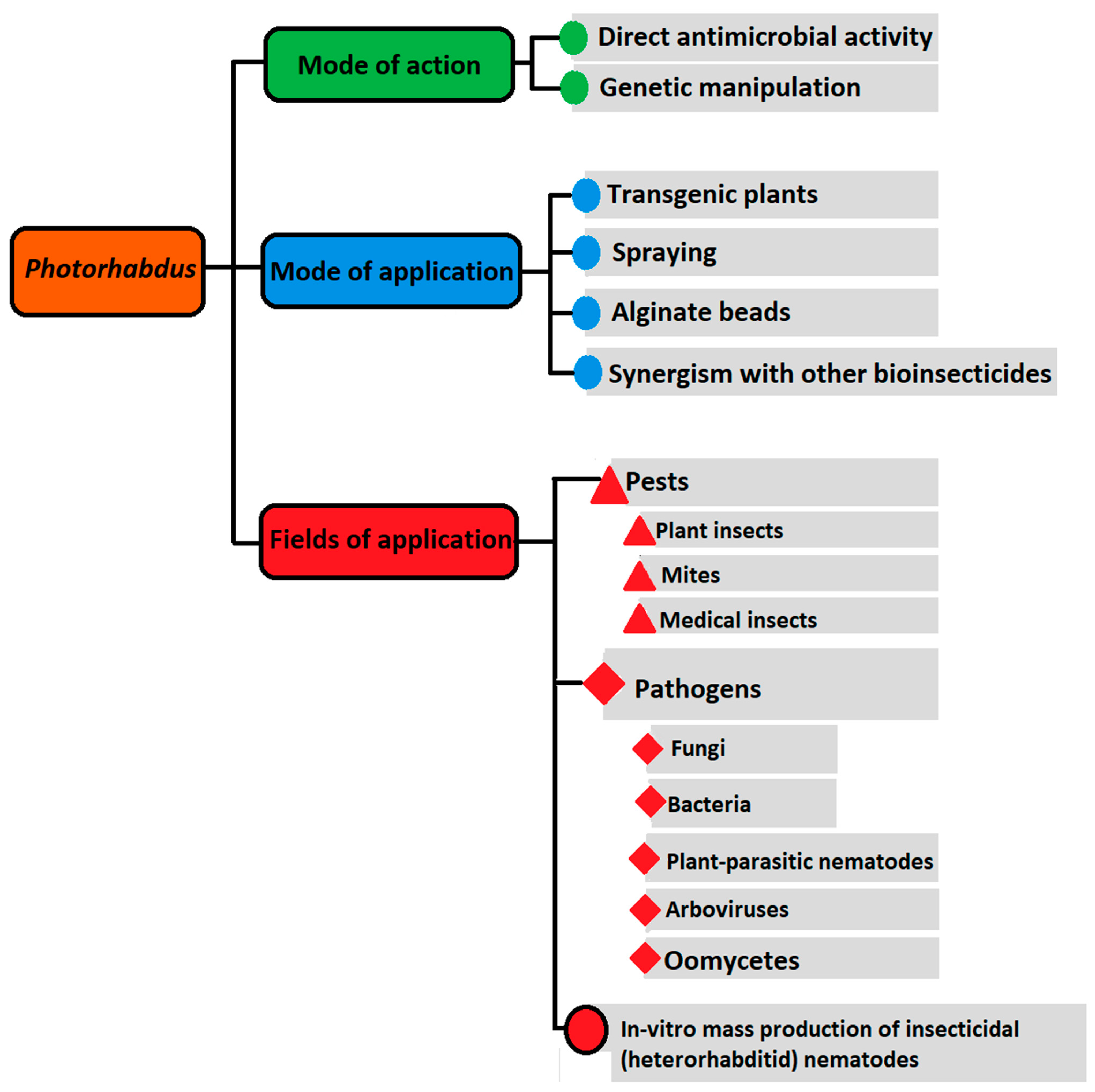
:1. Introduction
2. Taxa, Diversity, and Lifestyle of Photorhabdus spp.
2.1. Their Taxa and Diversity

{kind=link}
| Photorhabduss Species | Heterorhabditis Species | References |
|---|---|---|
| P. akhurstii | H. indica | [18,19] |
| P. asymbiotica | undescribed species | [18,20] |
| P. australis subsp. Thailandensis subsp. australis | H. gerrardi, H. indica | [13,19,20] |
| P. bodei | H. beicherriana | [19] |
| P. caribbeanensis | H. bacteriophora | [19,21] |
| P. cinerea | H. downesi, H. megidis, H. bacteriophora | [19,22] |
| P. hainanensis | undescribed species | [19,21] |
| P. heterorhabditis subsp. Aluminescens subsp. heterorhabditis | H. zealandica | [13,23] |
| P. kayaii | H. bacteriophora | [19,24] |
| P. khanii | H. bacteriophora | [19,21] |
| subsp. guanajuatensis | H. atacamensis | [25] |
| P. kleinii | H. georgiana, H. bacteriophora | [19,26] |
| P. laumondii subsp. clarkei subsp. laumondii | H. bacteriophora | [18,19] |
| P. luminescens | H. bacteriophora, H. indica | [27,28] |
| subsp. sonorensis | H. sonorensis | [29] |
| subsp. mexicana | H. mexicana | [25] |
| P. namnaonensis | H. baujardi | [19,30] |
| P. noenieputensis | H. indica, Heterorhabditis sp. | [19,31] |
| P. stackebrandtii | H. bacteriophora, H. georgiana | [19,32] |
| P. tasmanensis | H. zealandica, H. marelatus | [19,21] |
| P. temperata | H. megidis, H. downesi, H. zealandica | [18,19] |
| P. thracensis | H. bacteriophora | [19,21,24] |
| P. aegyptia | H. indica | [13] |
2.2. The Lifestyle of Photorhabdus spp.
3. Pathogenicity of Photorhabdus spp.
3.1. Range and Magnitude of Pathogenicity
3.2. Mode of Action of the Bacteria and Their Secreted Compounds
4. Pros and Cons of Photorhabdus spp.
4.1. The Positive Aspects
4.1.1. Cost-Effective Photorhabdus Mass Culture with Boosting Insecticidal Activity
4.1.2. The Mounting Role of Photorhabdus Bacteria against Pests and Pathogens
4.1.3. The Bacterial Metabolites as New Drugs for Diseases
4.1.4. Photorhabdus-Derived Natural Compounds as a Source for Industrial Products
4.2. Avoiding Negative Aspects
5. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xiao, Y.; Wu, K. Recent progress on the interaction between insects and Bacillus thuringiensis crops. Philos. Trans. R. Soc. 2019, 374, 20180316. [Google Scholar] [CrossRef] [Green Version]
- da Silva, W.J.; Pilz-Júnior, H.L.; Heermann, R.; da Silva, O.S. The great potential of entomopathogenic bacteria Xenorhabdus and Photorhabdus for mosquito control: A review. Parasites Vectors 2020, 13, 376. [Google Scholar] [CrossRef]
- Coupland, J.; Abd-Elgawad, M.M.; Askary, T.H. Beneficial nematodes and the changing scope of crop protection. In Biocontrol Agents: Entomopathogenic and Slug Parasitic Nematodes; Abd-Elgawad, M.M.M., Askary, T.H., Coupland, J., Eds.; CAB International: Wallingford, UK, 2017; pp. 26–42. [Google Scholar]
- Migunova, V.D.; Sasanelli, N. Bacteria as biocontrol tool against phytoparasitic nematodes. Plants 2021, 10, 389. [Google Scholar] [CrossRef] [PubMed]
- Abd-Elgawad, M.M.M. Toxic secretions of Photorhabdus and their efficacy against crop insect pests. In Biocontrol Agents: Entomopathogenic and Slug Parasitic Nematodes; Abd-Elgawad, M.M.M., Askary, T.H., Coupland, J., Eds.; CAB International: Wallingford, UK, 2017; pp. 231–260. [Google Scholar]
- Eroglu, C.; Cimenb, H.; Ulug, D.; Karagoz, M.; Hazir, S.; Cakmaka, I. Acaricidal effect of cell-free supernatants from Xenorhabdus and Photorhabdus bacteria against Tetranychus urticae (Acari: Tetranychidae). J. Inver. Pathol. 2019, 106, 61–66. [Google Scholar] [CrossRef]
- Ffrench-Constant, R.; Waterfield, N.; Daborn, P. Insecticidal toxins from Photorhabdus and Xenorhabdus. In Encyclopedia of Microbiology, 4th ed.; Schmid, T.M., Ed.; Academic Press: New York, NY, USA, 2019; pp. 704–715. [Google Scholar] [CrossRef]
- Muangpat, P.; Suwannaroj, M.; Yimthin, T.; Fukruksa, C.; Sitthisak, S.; Chantratita, N.; Vitta, A.; Thanwisai, A. Antibacterial activity of Xenorhabdus and Photorhabdus isolated from entomopathogenic nematodes against antibiotic-resistant bacteria. PLoS ONE 2020, 15, e0234129. [Google Scholar] [CrossRef]
- Ahuja, A.; Kushwah, J.; Mathur, C.; Chauhan, K.; Dutta, T.K.; Somvanshi, V.S. Identification of Galtox, a new protein toxin from Photorhabdus bacterial symbionts of Heterorhabditis. Toxicon 2021, 194, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Park, H.; Sampathkumar, P.; Perez, C.E.; Lee, J.H.; Tran, J.; Bonanno, J.B.; Hallem, E.A.; Almo, S.C.; Crawford, J.M. Stilbene epoxidation and detoxification in a Photorhabdus luminescens-nematode symbiosis. J. Biol. Chem. 2017, 292, 6680–6694. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Ren, J.; Zhan, J. Identification and characterization of an efficient phenylalanine ammonia-lyase from Photorhabdus luminescens. Appl. Biochem. Biotech. 2012, 193, 1099–1115. [Google Scholar] [CrossRef] [PubMed]
- Kushwah, J.; Kumar, P.; Garg, V.; Somvanshi, V.S. Discovery of a highly virulent strain of Photorhabdus luminescens ssp. akhurstii from Meghalaya, India. Indian J. Microbiol. 2017, 57, 125–128. [Google Scholar] [CrossRef] [Green Version]
- Machado, R.A.R.; Muller, A.; Ghazal, S.; Thanwisai, A.; Pagès, S.; Bode, H.B.; Hussein, M.A.; Khalil, K.M.; Tisa, L.S. Photorhabdus heterorhabditis subsp. aluminescens subsp. nov., Photorhabdus heterorhabditis subsp. heterorhabditis subsp. nov., Photorhabdus australis subsp. thailandensis subsp. nov., Photorhabdus australis subsp. australis subsp. nov., and Photorhabdus aegyptia sp. nov. isolated from Heterorhabditis entomopathogenic nematodes. Int. J. Syst. Evol. Microbiol. 2021, 71. [Google Scholar] [CrossRef]
- Ajnaga, E.; Kazimierczak, W. Evolution and taxonomy of nematode-associated entomopathogenic bacteria of the genera Xenorhabdus and Photorhabdus: An overview. Symbiosis 2020, 80, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Abd-Elgawad, M.M.M. Can rational sampling maximise isolation and fix distribution measure of entomopathogenic nematodes? Nematology 2020, 22, 907–916. [Google Scholar] [CrossRef]
- Abd-Elgawad, M.M.M. Optimizing sampling and extraction methods for plant-parasitic and entomopathogenic nematodes. Plants 2021, 10, 629. [Google Scholar] [CrossRef]
- Yooyangket, T.; Muangpat, P.; Polseela, R.; Tandhavanant, S.; Thanwisai, A.; Vitta, A. Identification of entomopathogenic nematodes and symbiotic bacteria from Nam Nao National Park in Thailand and larvicidal activity of symbiotic bacteria against Aedes aegypti and Aedes albopictus. PLoS ONE 2018, 13, e0195681. [Google Scholar] [CrossRef]
- Fischer-Le Saux, M.; Viallard, V.; Brunel, B.; Normand, P.; Boemare, N.E. Polyphasic classification of the genus Photorhabdus and proposal of new taxa: P. luminescens subsp. luminescens subsp. nov., P. luminescens subsp. akhurstii subsp. nov., P. luminescens subsp. laumondii subsp. nov., P. temperata sp. nov., P. temperate subsp. temperatas ubsp. nov. and P. asymbiotica sp. nov. Int. J. Syst. Bacteriol. 1999, 49, 1645–1656. [Google Scholar]
- Machado, R.A.R.; Wüthrich, D.; Kuhnert, P.; Arce, C.C.M.; Thönen, L.; Ruiz, C.; Zhang, X.; Robert, C.A.M.; Karimi, J.; Kamali, S.; et al. Whole-genome-based revisit of Photorhabdus phylogeny: Proposal for the elevation of most Photorhabdus subspecies to the species level and description of one novel species Photorhabdus bodei sp. nov., and one novel subspecies Photorhabdus laumondii subsp. clarkei subsp. nov. Int. J. Syst. Evol. Microbiol. 2018, 68, 2664–2681. [Google Scholar]
- Akhurst, R.J.; Boemare, N.E.; Janssen, P.H.; Peel, M.M.; Alfredson, D.A.; Beard, C.E. Taxonomy of Australian clinical isolates of the genus Photorhabdus and proposal of Photorhabdus asymbiotica subsp. asymbiotica subsp. nov. and P. asymbiotica subsp. australis subsp. nov. Int. J. Syst. Evol. Microbiol. 2004, 54, 1301–1310. [Google Scholar] [CrossRef] [Green Version]
- Tailliez, P.; Laroui, C.; Ginibre, N.; Paule, A.; Pages, S.; Boemare, N. Phylogeny of Photorhabdus and Xenorhabdus based on universally conserved protein-coding sequences and implications for the taxonomy of these two genera. Proposal of new taxa. X. vietnamensis sp. nov., P. luminescens subsp. caribbeanensis subsp. nov., P. luminescens subsp. hainanensis subsp. nov., P. temperata subsp. khanii subsp. nov., P. temperata subsp. tasmaniensis subsp. nov., and the reclassification of P. luminescens subsp. thracensis as P. temperata subsp. thracensis comb. nov. Int. J. Syst. Evol. Microbiol. 2010, 60, 1921–1937. [Google Scholar] [PubMed] [Green Version]
- Tóth, T.; Lakatos, T. Photorhabdus temperata subsp. cinerea subsp. nov., isolated from Heterorhabditis nematodes. Int. J. Syst. Evol. Microbiol. 2008, 58, 2579–2581. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, T.; Reenen, C.A.; Endo, A.; Tailliez, P.; Pages, S.; Sproer, C.; Malan, A.P.; Dicks, L.M.T. Photorhabdus heterorhabditis sp. nov., a symbiont of the entomopathogenic nematode Heterorhabditis zealandica. Int. J. Syst. Evol. Microbiol. 2014, 64, 1540–1545. [Google Scholar] [CrossRef] [Green Version]
- Hazir, S.; Stackebrandt, E.; Lang, E.; Schumann, P.; Ehlers, R.U.; Keskin, N. Two new subspecies of Photorhabdus luminescens, isolated from Heterorhabditis bacteriophora (Nematoda: Heterorhabditidae): Photorhabdus luminescens subsp. Kayaii subsp. nov. and Photorhabdus luminescens subsp. thracensis subsp. nov. Syst. Appl. Microbiol. 2004, 27, 36–42. [Google Scholar] [CrossRef]
- Machado, R.A.R.; Bruno, P.; Arce, C.C.M.; Liechti, N.; Köhler, A.; Bernal, J.; Bruggmann, R.; Turlings, T.C.J. Photorhabdus khanii subsp. guanajuatensis subsp. nov., isolated from Heterorhabditis atacamensis, and Photorhabdus luminescens subsp. Mexicana subsp. nov., isolated from Heterorhabditis mexicana entomopathogenic nematodes. Int. J. Syst. Evol. Microbiol. 2019, 69, 652–661. [Google Scholar] [CrossRef]
- An, R.; Grewal, P.S. Photorhabdus luminescens subsp. kleinii subsp. nov. (Enterobacteriales: Enterobacteriaceae). Curr. Microbiol. 2011, 62, 539–543. [Google Scholar] [CrossRef]
- Thomas, G.M.; Poinar, G.O. Xenorhabdus gen. nov., a genus of entomopathogenic nematophilic bacteria of the family Enterobacteriaceae. Int. J. Syst. Bacteriol. 1979, 29, 352–360. [Google Scholar] [CrossRef] [Green Version]
- Boemare, N.E.; Akhurst, R.J.; Mourant, R.G. DNA relatedness between Xenorhabdus spp. (Enterobacteriaceae), symbiotic bacteria of entomopathogenic nematodes, and a proposal to transfer Xenorhabdus luminescens to a new genus, Photorhabdus gen. nov. Int. J. Syst. Bacteriol. 1993, 43, 249–255. [Google Scholar] [CrossRef] [Green Version]
- Orozco, R.A.; Hill, T.; Stock, S.P. Characterization and phylogenetic relationships of Photorhabdus luminescens subsp. sonorensis (γ-Proteobacteria: Enterobacteriaceae), the bacterial symbiont of the entomopathogenic nematode Heterorhabditis sonorensis (Nematoda: Heterorhabditidae). Curr. Microbiol. 2013, 66, 30–39. [Google Scholar] [CrossRef]
- Glaeser, S.P.; Tobias, N.J.; Thanwisai, A.; Chantratita, N.; Bode, H.B.; Kampfer, P. Photorhabdus luminescens subsp. namnaonensis subsp. nov., isolated from Heterorhabditis baujardi nematodes. Int. J. Syst. Evol. Microbiol. 2017, 67, 1046–1051. [Google Scholar] [CrossRef]
- Ferreira, T.; Reenen, C.A.; Pages, S.; Tailliez, P.; Malan, A.P.; Dicks, L.M.T. Photorhabdus luminescens subsp. noenieputensis subsp. nov., a symbiotic bacterium associated with a novel Heterorhabditis species related to Heterorhabditis indica. Int. J. Syst. Evol. Microbiol. 2013, 63, 1853–1858. [Google Scholar] [CrossRef]
- An, R.; Grewal, P.S. Molecular mechanisms of persistence of mutualistic bacteria Photorhabdus in the entomopathogenic nematode host. PLoS ONE 2010, 5, e13154. [Google Scholar] [CrossRef] [Green Version]
- Boemare, N. Biology, taxonomy and systematics of Photorhabdus and Xenorhabdus. In Entomopathogenic Nematology; Gaugler, R., Ed.; CAB International: Wallingford, UK, 2002; pp. 35–56. [Google Scholar]
- Plichta, K.L.; Joyce, S.A.; Clarke, D.; Waterfield, N.; Stock, S.P. Heterorhabditis gerrardi n. sp. (Nematoda: Heterorhabditidae): The hidden host of Photorhabdus asymbiotica (Enterobacteriaceae:g-Proteobacteria). J. Helminthol. 2009, 83, 309–320. [Google Scholar] [CrossRef]
- Gerrard, J.; Waterfield, N.; Vohra, R.; ffrench-Constant, R. Human infection with Photorhabdus asymbiotica: An emerging bacterial pathogen. Microbes Infect./Inst. Pasteur. 2004, 6, 229–237. [Google Scholar] [CrossRef]
- Piedra-Buena, A.; López-Cepero, J.; Campos-Herrera, R. Entomopathogenic nematode production and application: Regulation, ecological impact and non-target effects. In Nematodes Pathogenesis of Insects and Other Pests; Campos-Herrera, R., Ed.; Springer International Publishing: Cham, Switzerland, 2015; pp. 255–282. [Google Scholar]
- Askary, T.H.; Abd-Elgawad, M.M.M. Opportunities and challenges of entomopathogenic nematodes as bio control agents in their tripartite interactions. Egypt. J. Biol. Pest Cont. 2021, 31, 42. [Google Scholar] [CrossRef]
- Goodrich-Blair, H.; Clarke, D.J. Mutualism and pathogenesis in Xenorhabdus and Photorhabdus: Two roads to the same destination. Molecul. Microbiol. 2007, 64, 260–268. [Google Scholar] [CrossRef] [PubMed]
- Ciche, T.A.; Kim, K.S.; Kaufmann-Daszczuk, B.; Nguyen, K.C.Q.; Hall, D.H. Cell invasion and matricide during Photorhabdus luminescens transmission by Heterorhabditis bacteriophora nematodes. Appl. Environ. Microbiol. 2008, 74, 2275–2287. [Google Scholar] [CrossRef] [Green Version]
- Chaston, J.M.; Suen, G.; Tucker, S.L.; Andersen, A.W.; Bhasin, A.; Bode, E. The entomopathogenic bacterial endosymbionts Xenorhabdus and Photorhabdus: Convergent lifestyles from divergent genomes. PLoS ONE 2011, 6, e27909. [Google Scholar] [CrossRef]
- Maneesakorn, P.; An, R.S.; Daneshvar, H.; Taylor, K.; Bai, X.D.; Adams, B.J.; Grewal, P.S.; Chandrapatya, A. Phylogenetic and cophylogenetic relationships of entomopathogenic nematodes (Heterorhabditis: Rhabditida) and their symbiotic bacteria (Photorhabdus: Enterobacteriaceae). Mol. Phylogenet. Evol. 2011, 59, 271–280. [Google Scholar] [CrossRef]
- Kazimierczak, W.; Skrzypek, H.; Sajnaga, E.; Skowronek, M.; Wasko, A.; Kreft, A. Strains of Photorhabdus spp. associated with polish Heterorhabditis isolates: Their molecular and phenotypic characterization and symbiont exchange. Arch. Microbiol. 2017, 199, 979–989. [Google Scholar] [CrossRef]
- Maher, A.M.D.; Asaiyah, M.A.M.; Brophy, C.; Griffin, C.T. An entomopathogenic nematode extends its niche by associating with different symbionts. Microb. Ecol. 2017, 73, 211–223. [Google Scholar] [CrossRef]
- Adams, B.J.; Fodor, A.; Koppenhofer, H.S.; Stackebrandt, E.; Patricia, S.; Klein, M.G. Biodiversity and systematics of nematode-bacterium entomopathogens. Biol. Control 2006, 37, 32–49. [Google Scholar] [CrossRef]
- Maher, A.M.D.; Asaiyah, M.; Quinn, S.; Burke, R.; Wolff, H.; Bod, H.B.; Griffin, C.T. Competition and co-existence of two Photorhabdus symbionts with a nematode host. Microb. Ecol. 2021, 81, 223–239. [Google Scholar] [CrossRef]
- Abd-Elgawad, M.M.M. Towards optimization of entomopathogenic nematodes for more service in the biological control of insect pests. Egypt. J. Biol. Pest Cont. 2019, 29, 77. [Google Scholar] [CrossRef]
- Koppenhöfer, A.M.; Shapiro-Ilan, D.I.; Hiltpold, I. Entomopathogenic nematodes in sustainable food production. Front. Sustain. Food Syst. 2020, 4, 125. [Google Scholar] [CrossRef]
- Shapiro-Ilan, D.; Hazir, S.; Glazer, I. Advances in use of entomopathogenic nematodes in integrated pest management. In Integrated Management of Insect Pests: Current and Future Developments; Kogan, M., Heinrichs, E.A., Eds.; Burleigh Dodds Science Publication: Cambridge, UK, 2020; pp. 1–30. [Google Scholar]
- Clarke, D.J. Photorhabdus: A model for the analysis of pathogenicity and mutualism. Cell Microbiol. 2008, 10, 2159–2167. [Google Scholar] [CrossRef]
- Morgan, J.A.W.; Kuntzelmann, V.; Tavernor, S.; Ousley, M.A.; Winstanley, C. Survival of Xenorhabdus nematophilus and Photorhabdus luminescens in water and soil. J. Appl. Microbiol. 1997, 83, 665–670. [Google Scholar] [CrossRef]
- Mohan, S.; Raman, R.; Gaur, H.S. Foliar application of Photorhabdus luminescens, symbiotic bacteria from entomopathogenic nematode Heterorhabditis indica, to kill cabbage butterfly Pieris brassicae. Curr. Sci. 2003, 84, 1397. [Google Scholar]
- Bhatnagar, R.K.; Rajagopal, R.; Rao, N.G.V. Biopesticidal Compositions. World Intellectual Property. Patent WO 2005/055724 A1. Available online: https://patentscope.wipo.int/search/en/detail.jsf?docId=WO2005055724 (accessed on 23 May 2021).
- Rajagopal, R.; Mohan, S.; Bhatnagar, R.K. Direct infection of Spodoptera litura by Photorhabdus luminescens encapsulated in alginate beads. J. Invert. Pathol. 2006, 93, 50–53. [Google Scholar] [CrossRef] [PubMed]
- Kushwah, J.; Somvanshi, V.S. Photorhabdus: A microbial factory of insect-killing toxins. In Microbial Factories: Biodiversity, Biopolymers, Bioactive Molecules; Kalia, V.C., Ed.; Springer: New Delhi, India, 2015; Volume 2, pp. 235–240. [Google Scholar] [CrossRef]
- Antonello, A.M.; Sartori, T.; Folmer, C.A.P.; Brandelli, A.; Heermann, R.; Rodrigues, J.L.C.; Peres, A.; Romão, P.R.T.; Da Silva, O.S. Entomopathogenic bacteria Photorhabdus luminescens as drug source against Leishmania amazonensis. Parasitology 2018, 145, 1065–1074. [Google Scholar] [CrossRef] [PubMed]
- Antonello, A.M.; Sartori, T.; Silva, M.B.; Prophiro, J.S.; Pinge-Filho, P.; Heermann, R. Anti-Trypanosoma activity of bioactive metabolites from Photorhabdus luminescens and Xenorhabdus nematophila. Exp. Parasitol. 2019, 204, 107724. [Google Scholar] [CrossRef]
- An, R.; Grewal, P.S. Photorhabdus temperata subsp. Stackebrandtii subsp. nov. (Enterobacteriales: Enterobacteriaceae). Curr. Microbiol. 2011, 61, 291–297. [Google Scholar] [CrossRef]
- Pidot, S.J.; Coyne, S.; Kloss, F.; Hertweck, C. Antibiotics from neglected bacterial sources. Int. J. Med. Microbiol. 2014, 304, 14–22. [Google Scholar] [CrossRef]
- Ffrench-Constant, R.; Waterfield, N.; Daborn, P.; Joyce, S.; Bennett, H.; Au, C.; Dowling, A.; Boundy, S.; Reynolds, S.; Clarke, D. Photorhabdus: Towards a functional genomic analysis of a symbiont and pathogen. FEMS Microbiol. Rev. 2003, 26, 433–456. [Google Scholar] [CrossRef]
- Regaiolo, A.; Dominelli, N.; Andresen, K.; Heermann, R. The biocontrol agent and insect pathogen Photorhabdus luminescens interacts with plant roots. Appl. Environ. Microbiol. 2020, 86, e00891-20. [Google Scholar] [CrossRef] [PubMed]
- Eckstein, S.; Brehm, J.; Seidel, M.; Lechtenfeld, M.; Heermann, R. Two novel XRE-like transcriptional regulators control phenotypic heterogeneity in Photorhabdus luminescens cell populations. BMC Microbiol. 2021, 21, 63. [Google Scholar] [CrossRef] [PubMed]
- Eliceche, D.P.; Rusconi, J.M.; Rosales, M.N.; Salas, A.; Achinelly, M.F. Infectivity by nematode-bacteria association on the potato weevil Phyrdenus muriceus. Rev. Argent. Microbiol. 2020, 52, 256–257. [Google Scholar] [CrossRef]
- Mahar, A.N.; Munir, M.; Gowen, S.R.; Hague, N.G.M. Role of entomopathogenic bacteria, Photorhabdus luminescens and its toxic secretions against Galleria mellonella larvae. J. Entomol. 2005, 2, 69–76. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Razek, A.S. Pathogenic effects of Xenorhabdus nematophilus and Photorhabdus luminescens (Enterobacteriaceae) against pupae of the Diamondback Moth, Plutella xylostella (L.). J. Pest Sci. 2003, 76, 108–111. [Google Scholar] [CrossRef]
- Blackburn, M.B.; Domek, J.M.; Gelman, D.B.; Hu, J.S. The broadly insecticidal Photorhabdus luminescens toxin complex a (Tca): Activity against the Colorado potato beetle, Leptinotarsa decemlineata, and sweet potato whitefly, Bemisia tabaci. J. Insect Sci. 2005, 5, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Salazar-Gutiérrez, J.D.; Castelblanco, A.; Rodríguez-Bocanegra, M.X.; Teran, W.; Sáenz-Aponte, A. Photorhabdus luminescens subsp. akhurstii SL0708 pathogenicity in Spodoptera frugiperda (Lepidoptera: Noctuidae) and Galleria mellonella (Lepidoptera: Pyralidae). J. Asia-Pacific Entomol. 2017, 20, 1112–1121. [Google Scholar] [CrossRef]
- Tounsi, S.; Aoun, A.E.; Blight, M.; Rebai, A.; Jaoua, S. Evidence of oral toxicity of Photorhabdus temperate strain K122 against Prays oleae and its improvement by heterologous expression of Bacillus thuringiensis cry 1Aa and cry 1Ia genes. J. Invert. Pathol. 2006, 91, 131–135. [Google Scholar] [CrossRef] [PubMed]
- Jallouli, W.; Abdelkefi-Mesrati, L.; Tounsi, S.; Jaoua, S.; Zouari, N. Potential of Photorhabdus temperate K122 bioinsecticide in protecting wheat flour against Ephestia kuehniella. J. Stored Products Res. 2013, 53, 61–66. [Google Scholar] [CrossRef]
- Bock, C.H.; Shapiro-Ilan, D.I.; Wedge, D.E.; Cantrell, C.L. Identification of the antifungal compound, trans-cinnamic acid, produced by Photorhabdus luminescens, a potential bio-pesticide against pecan scab. J. Pest Sci. 2014, 87, 155–162. [Google Scholar] [CrossRef]
- Bowen, D.J.; Ensign, J.C. Purification and characterization of a high molecular- weight insecticidal protein complex produced by the entomopathogenic bacterium Photorhabdus luminescens. Appl. Environ. Microbiol. 1998, 64, 3029–3035. [Google Scholar] [CrossRef] [Green Version]
- Dunphy, G.B.; Webster, J.M. Virulence mechanisms of Heterorhabditis heliothidis and its bacterial associate, Xenorhabdus luminescens, in nonimmune larvae of the greater wax moth, Galleria mellonella. Int. J. Parasitol. 1988, 18, 729–737. [Google Scholar] [CrossRef]
- English, L.; Slatin, S.L. Mode of action of delta-endotoxins from Bacillus thuringiensis: A comparison with other bacterial toxins. Insect Biochem. Mol. Biol. 1992, 22, 1–7. [Google Scholar] [CrossRef]
- Williamson, V.M.; Kaya, H.K. Sequence of a symbiont. Nat. Biotech. 2003, 21, 1924–1925. [Google Scholar] [CrossRef]
- Duchaud, E.; Rusniok, C.; Frangeul, L.; Buchrieser, C.; Givaudan, A.; Taourit, S. The genome sequence of the entomopathogenic bacterium Photorhabdus luminescens. Nat. Biotechnol. 2003, 21, 1307–1313. [Google Scholar] [CrossRef]
- Kronenwerth, M.; Brachmann, A.O.; Kaiser, M.; Bode, H.B. Bioactive derivatives of isopropylstilbene from mutasynthesis and chemical synthesis. ChemBioChem 2014, 15, 2689–2691. [Google Scholar] [CrossRef] [PubMed]
- Challinor, V.L.; Bode, H.B. Bioactive natural products from novel microbial sources. Ann. N.Y. Acad. Sci. 2015, 1354, 82–97. [Google Scholar] [CrossRef]
- Jallouli, W.; Zouari, N.; Jaoua, S. Involvement of oxidative stress and growth at high cell density in the viable but nonculturable state of Photorhabdus temperata ssp. temperata strain K122. Process. Biochem. 2010, 45, 706–713. [Google Scholar] [CrossRef]
- Vlisidou, I.; Hapeshi, A.; Healey, J.R.; Smart, K.; Yang, G.; Waterfield, N.R. The Photorhabdus asymbiotica virulence cassettes deliver protein effectors directly into target eukaryotic cells. eLife 2019, 8, e46259. [Google Scholar] [CrossRef]
- Daborn, P.J.; Waterfield, N.; Silva, C.P.; Au, C.P.Y.; Sharma, S.; Ffrench-Constant, R.H. A single Photorhabdus gene, makes caterpillars floppy (mcf), allows Escherichia coli to persist within and kill insects. Fixed Point Theory Appl. 2002, 99, 10742–10747. [Google Scholar] [CrossRef] [Green Version]
- Dutta, T.K.; Mathur, C.; Mandal, A.; Somvanshi, V.S. The differential strain virulence of the candidate toxins of Photorhabdus akhurstii can be correlated with their inter-strain gene sequence diversity. Biotech 2020, 10, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; De Souza, R.F.; Anantharaman, V.; Iyer, L.M.; Aravind, L. Polymorphic toxin systems: Comprehensive characterization of trafficking modes, processing, mechanisms of action, immunity and ecology using comparative genomics. Biol. Direct 2012, 7, 18. [Google Scholar] [CrossRef] [Green Version]
- Ffrench-Constant, R.H.; Dowling, A.J. Photorhabdus toxins. In Advances in Insect Physiology; Dhadialla, T.S., Gill, S.S., Eds.; Elsevier Academic Press: London, UK, 2014; Volume 47, pp. 343–388. [Google Scholar]
- Zhou, X.; Kaya, H.K.; Heungens, K.; Goodrich-Blair, H. Response of ants to a deterrent factor(s) produced by the symbiotic bacteria of entomopathogenic nematodes. Appl. Environ. Microbiol. 2002, 68, 6202–6209. [Google Scholar] [CrossRef] [Green Version]
- Shapiro-Ilan, D.I.; Reilly, C.C.; Hotchkiss, M.W. Suppressive effects of metabolites from Photorhabdus and Xenorhabdus spp. on phytopathogens of peach and pecan. Arch. Phytopathol. Plant Protect. 2009, 42, 715–728. [Google Scholar] [CrossRef]
- Shahina, F.; Tabassum, K.A.; Salma, J.; Mahreen, G. Biopesticidal affect of Photorhabdus luminescens against Galleria mellonella larvae and subteranean termite (Termitidae: Macrotermis). Pak. J. Nematol. 2011, 29, 35–43. [Google Scholar]
- Erler, S.; Popp, M.; Lattorff, M. Dynamics of immune system gene expression upon bacterial challenge and wounding in a social insect (Bombus terrestris). PLoS ONE 2011, 6, e18126. [Google Scholar] [CrossRef]
- Abd El-Zaher, F.H.; Abd-Elgawad, M.M.M.; Abd El-Maksoud, H.K. Use of the entomopathogenic nematode symbiont Photorhabdus luminescens as a biocontrol agent. B- Factors affecting the cell-free filtrates from the bacterium. J. Appl. Sci. Res. 2012, 8, 4600–4614. [Google Scholar]
- Shapiro-Ilan, D.I.; Bock, C.H.; Hotchkiss, M.W. Suppression of pecan and peach pathogens on different substrates using Xenorhabdus bovienii and Photorhabdus luminescens. Biol. Cont. 2014, 77, 1–6. [Google Scholar] [CrossRef]
- Abd El-Zaher, F.H.; Abd El-Maksoud, H.K.; Abd-Elgawad, M.M.M. Use of Photorhabdus as a biopesticides. A-Cell suspension from Photorhabdus sp. against Galleria mellonella insect. Catrina 2008, 3, 125–129. [Google Scholar]
- Keskes, S.; Jallouli, W.; Atitallah, I.B.; Driss, F.; Sahli, E.; Chamkha, M.; Touns, S. Development of a cost-effective medium for Photorhabdus temperata bioinsecticide production from wastewater and exploration of performance kinetic. Sci. Rep. 2021, 11, 779. [Google Scholar] [CrossRef]
- Wang, Y.; Bilgrami, A.L.; Shapiro-Ilan, D.; Gaugler, R. Stability of entomopathogenic bacteria, Xenorhabdus nematophila and Photorhabdus luminescens, during in vitro culture. J. Int. Microbiol. Biotechnol. 2007, 34, 73–81. [Google Scholar] [CrossRef]
- Orozco-Hidalgo, M.T.; Quevedo-Hidalgo, B.; Sáenz-Aponte, A. Growth kinetics and pathogenicity of Photorhabdus luminescens subsp. Akhurstii SL0708. Egypt. J. Biol. Pest Cont. 2019, 29, 71. [Google Scholar] [CrossRef]
- Jallouli, W.; Hammami, W.; Zouari, N.; Jaoua, S. Medium optimization for biomass production and morphology variance overcome of Photorhabdus temperata ssp. temperata strain K122. Process. Biochem. 2008, 43, 1338–1344. [Google Scholar] [CrossRef]
- Hinchliffe, S.J. Insecticidal toxins from the Photorhabdus and Xenorhabdus bacteria. Open Toxinol. J. 2010, 3, 101–118. [Google Scholar] [CrossRef]
- Tobias, N.J.; Wolff, H.; Djahanschiri, B.; Grundmann, F.; Kronenwerth, M.; Shi, Y.M. Natural product diversity associated with the nematode symbionts Photorhabdus and Xenorhabdus. Nat. Microbiol. 2017, 2, 1676–1685. [Google Scholar] [CrossRef]
- Bode, H.B. Entomopathogenic bacteria as a source of secondary metabolites. Curr. Opin. Chem. Biol. 2009, 13, 224–230. [Google Scholar] [CrossRef]
- Bussaman, P.; Sermswan, R.W.; Grewal, P.S. Toxicity of the entomopathogenic bacteria Photorhabdus and Xenorhabdus to the mushroom mite (Luciaphorus sp.; Acari: Pygmephoridae). Biocont. Sci. Tech. 2006, 16, 245–256. [Google Scholar] [CrossRef]
- Hazir, S.; Shapiro-Ilan, D.I.; Bock, C.H.; Hazir, C.; Leite, L.G.; Hotchkiss, M.W. Relative potency of culture supernatants of Xenorhabdus and Photorhabdus spp. on growth of some fungal phytopathogens. Eur. J. Plant Pathol. 2016, 146, 369–381. [Google Scholar] [CrossRef]
- Shehata, I.E.; Hammam, M.M.A.; El-Borai, F.E.; Duncan, L.W.; Abd-Elgawad, M.M.M. Comparison of virulence, reproductive potential, and persistence among local Heterorhabditis indica populations for the control of Temnorhynchus baal (Reiche & Saulcy) (Coleoptera: Scarabaeidae) in Egypt. Egy. J. Biol. Pest Cont. 2019, 29, 32. [Google Scholar] [CrossRef]
- Shehata, I.E.; Hammam, M.M.A.; El-Borai, F.E.; Duncan, L.W.; Abd-Elgawad, M.M.M. Traits of the entomopathogenic nematode, Heterorhabditis bacteriophora (Hb-EG strain), for potential biocontrol in strawberry fields. Egy. J. Biol. Pest Cont. 2020, 30, 40. [Google Scholar] [CrossRef] [Green Version]
- Abd El-Zaher, F.H.; Abd-Elgawad, M.M.M. Assessment of the antagonistic potential for the bacterial symbionts of entomopathogenic nematodes. J. Appl. Sci. Res. 2012, 8, 4590–4599. [Google Scholar]
- Jung, S.; Kim, Y. Synergistic effect of entomopathogenic bacteria (Xenorhabdus sp. and Photorhabdus temperata ssp. temperata) on the pathogenicity of Bacillus thuringiensis ssp. Aizawai against Spodoptera exigua (Lepidoptera: Noctuidae). Environ. Entomol. 2006, 35, 1584–1589. [Google Scholar] [CrossRef]
- Benfarhat-Touzri, D.; Ben Amira, A.; Khedher, B.S.; Givaudan, A.; Jaoua, S.; Tounsi, S. Combinatorial effect of Bacillus thuringiensis kurstaki and Photorhabdus luminescens against Spodoptera littoralis (Lepidoptera: Noctuidae). J. Basic Microbiol. 2014, 54, 1160–1165. [Google Scholar] [CrossRef] [PubMed]
- Gulcu, B.; Hazir, S.; Kaya, H.K. Scavenger deterrent factor (SDF) from symbiotic bacteria of entomopathogenic nematodes. J. Invertebr. Pathol. 2012, 110, 326–333. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Burton, S.; Glancy, T.; Li, Z.S.; Hampton, R.; Meade, T.; Merlo, D.J. Insect resistance conferred by 283-kDa Photorhabdus luminescens protein TcdA in Arabidopsis thaliana. Nat. Biotechnol. 2003, 21, 1222–1228. [Google Scholar] [CrossRef]
- Petell, J.K.; Merlo, D.J.; Herman, R.A.; Roberts, J.L.; Guo, L.; Schafer, B.W.; Sukhapinda, K.; Merlo, A.O. Transgenic Plants Expressing Photorhabdus Toxin. US Patent No. 2004/6717035 B2. Available online: https://www.google.com/patents/US6717035 (accessed on 3 June 2021).
- Rasmann, S.; Köllner, T.G.; Degenhardt, J.; Hiltpold, I.; Toepfer, S.; Kuhlmann, U.; Gershenzon, J.; Turlings, T.C.J. Recruitment of entomopathogenic nematodes by insect-damaged maize roots. Nature 2005, 434, 732–737. [Google Scholar] [CrossRef]
- Wilschut, R.A.; Geisen, S. Nematodes as drivers of plant performance in natural systems. Trends Plant Sci. 2020, 26, 237–247. [Google Scholar] [CrossRef]
- Caccia, M.; Marro, N.; Dueñas, J.R.; Doucet, M.E.; Lax, P. Effect of the entomopathogenic nematode-bacterial symbiont complex on Meloidogyne hapla and Nacobbus aberrans in short-term greenhouse trials. Crop Prot. 2018, 114, 162–166. [Google Scholar] [CrossRef] [Green Version]
- Abd-Elgawad, M.M.M. Optimizing biological control agents for controlling nematodes of tomato in Egypt. Egypt. J. Biol. Pest Cont. 2020, 30, 58. [Google Scholar] [CrossRef]
- Abd-Elgawad, M.M.M. Biological control agents in the integrated nematode management of pepper in Egypt. Egypt. J. Biol. Pest Cont. 2020, 30, 70. [Google Scholar] [CrossRef]
- Abd-Elgawad, M.M.M. Biological control agents in the integrated nematode management of potato in Egypt. Egypt. J. Biol. Pest Cont. 2020, 30, 121. [Google Scholar] [CrossRef]
- Abd-Elgawad, M.M.M. Biological control of nematodes infecting eggplant in Egypt. Bull. Natl. Res. Cent. 2021, 45, 6. [Google Scholar] [CrossRef]
- Stock, S.P.; Kusakabe, A.; Orozco, R.A. Secondary metabolites produced by Heterorhabditis symbionts and their application in agriculture: What we know and what to do next. J. Nematol. 2017, 49, 373–383. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.B.; Tabashnik, B.; Dennehy, T.; Patin, A.; Bartlet, A. Development time and resistance to Bt crops. Nature 1999, 400, 519. [Google Scholar] [CrossRef]
- Maithri, S.K.; Ramesh, K.V.; Mutangana, D. Theoretical structure prediction of TcaA from Photorhabdus luminescens and aminopeptidase N receptor from Helicoverpa armigera. Res. J. Recent Sci. 2013, 2, 40–49. [Google Scholar]
- Kumar, K.S.V.; Vendan, K.T.; Nagaraj, S.B. Isolation and characterization of entomopathogenic symbiotic bacterium, Photorhabdus luminescens of Heterorhabditis indica from soils of five agro climatic zones of Karnataka. Biosci. Biotech. Res. Asia 2014, 11, 129–139. [Google Scholar] [CrossRef]
- Kim, E.; Jeoung, S.; Park, Y.; Kim, K.; Kim, Y. A novel formulation of Bacillus thuringiensis for the control of brassica leaf beetle, Phaedon brassicae (Coleoptera: Chrysomelidae). J. Econ. Entomol. 2015, 108, 2556–2565. [Google Scholar] [CrossRef] [PubMed]
- Mohan, S.; Sabir, N. Biosafety concerns on the use of Photorhabdus luminescens as biopesticide: Experimental evidence of mortality in egg parasitoid Trichogramma spp. Curr. Sci. 2005, 89, 1268–1272. [Google Scholar]
- Fand, B.B.; Gautam, R.D.; Kamra, A.; Suroshe, S.S.; Mohan, S. Bioefficacy of aqueous garlic extract and a symbiotic bacterium, Photorhabdus luminescens against Phenacoccus solenopsis Tinsley (Homoptera: Pseudococcidae). Biopestic. Int. 2012, 8, 38–48. [Google Scholar]
- Nielsen-LeRoux, C.; Gaudriault, S.; Ramarao, N.; Lereclus, D.; Givaudan, A. How the insect pathogen bacteria Bacillus thuringiensis and Xenorhabdus/Photorhabdus occupy their hosts. Curr. Opin. Microbiol. 2012, 15, 220–231. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abd-Elgawad, M.M.M. Photorhabdus spp.: An Overview of the Beneficial Aspects of Mutualistic Bacteria of Insecticidal Nematodes. Plants 2021, 10, 1660. https://doi.org/10.3390/plants10081660
Abd-Elgawad MMM. Photorhabdus spp.: An Overview of the Beneficial Aspects of Mutualistic Bacteria of Insecticidal Nematodes. Plants. 2021; 10(8):1660. https://doi.org/10.3390/plants10081660
Chicago/Turabian StyleAbd-Elgawad, Mahfouz M. M. 2021. "Photorhabdus spp.: An Overview of the Beneficial Aspects of Mutualistic Bacteria of Insecticidal Nematodes" Plants 10, no. 8: 1660. https://doi.org/10.3390/plants10081660