
The Effect of Water Level in Rice Cropping System on Phosphorus Uptake Activity of Pup1 in a Pup1+Sub1 Breeding Line

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Genotypic Analysis of IPS
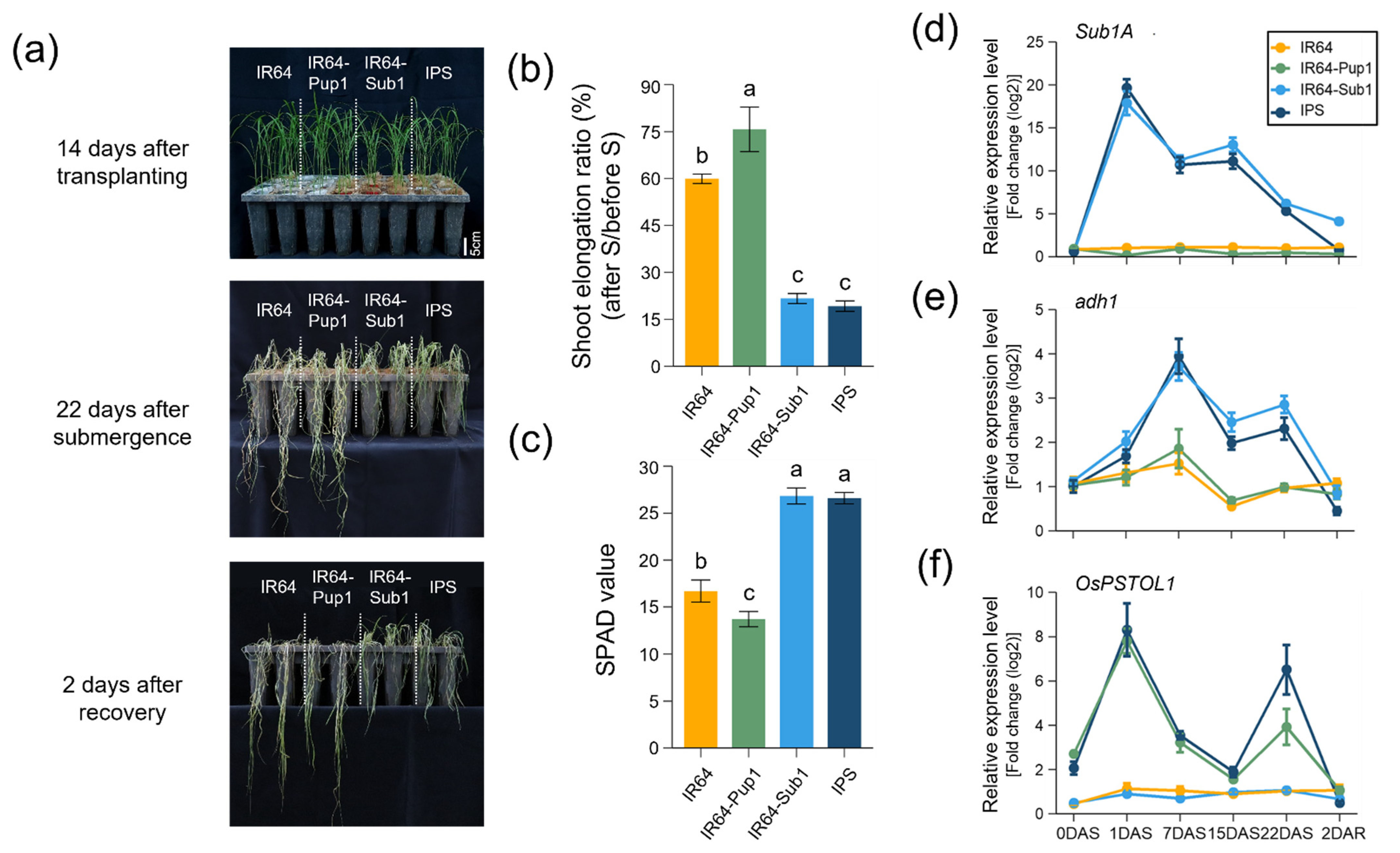
2.2. Phenotypic Evaluation and Gene Expression of IPS in Submerged Condition
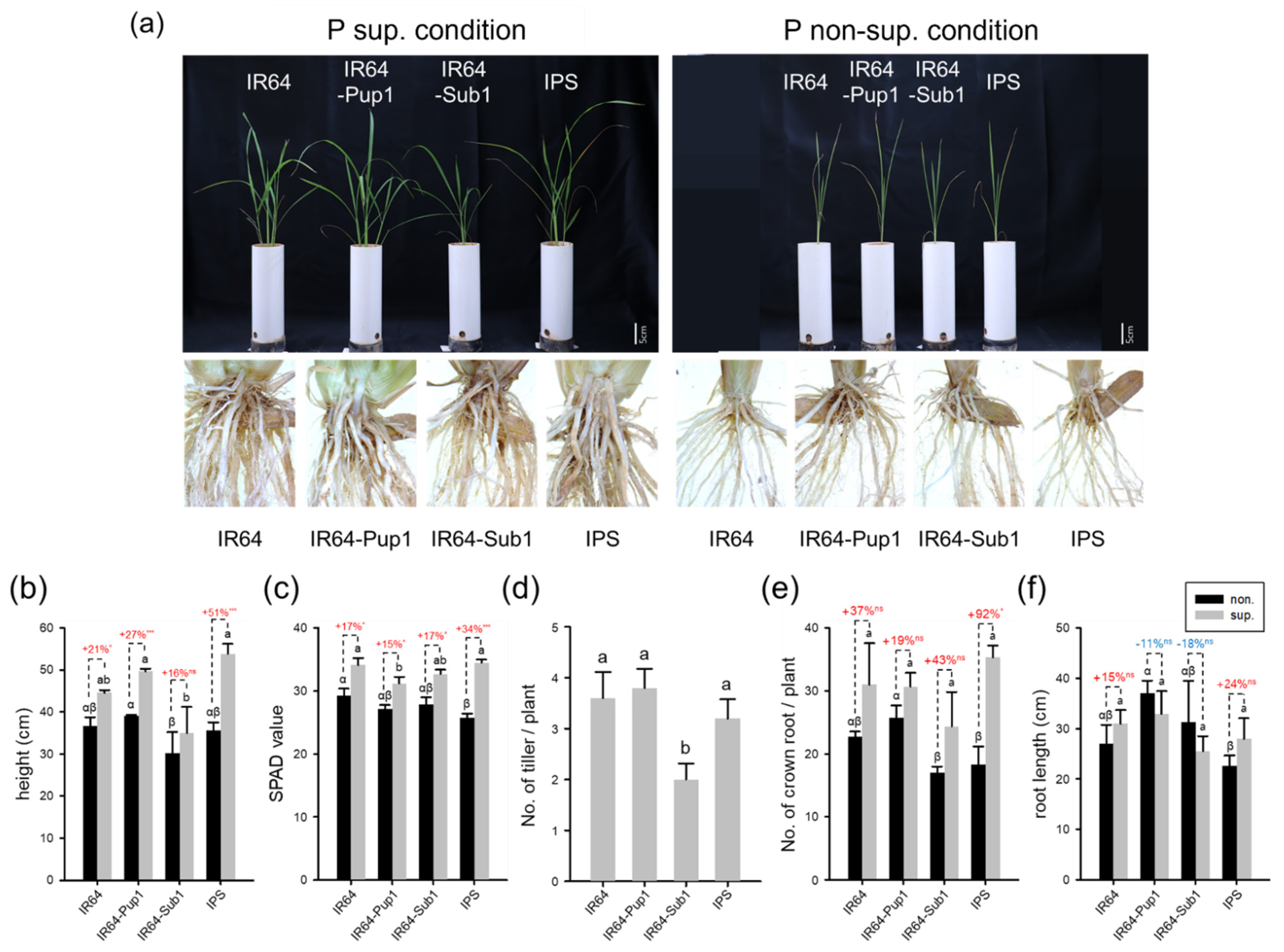
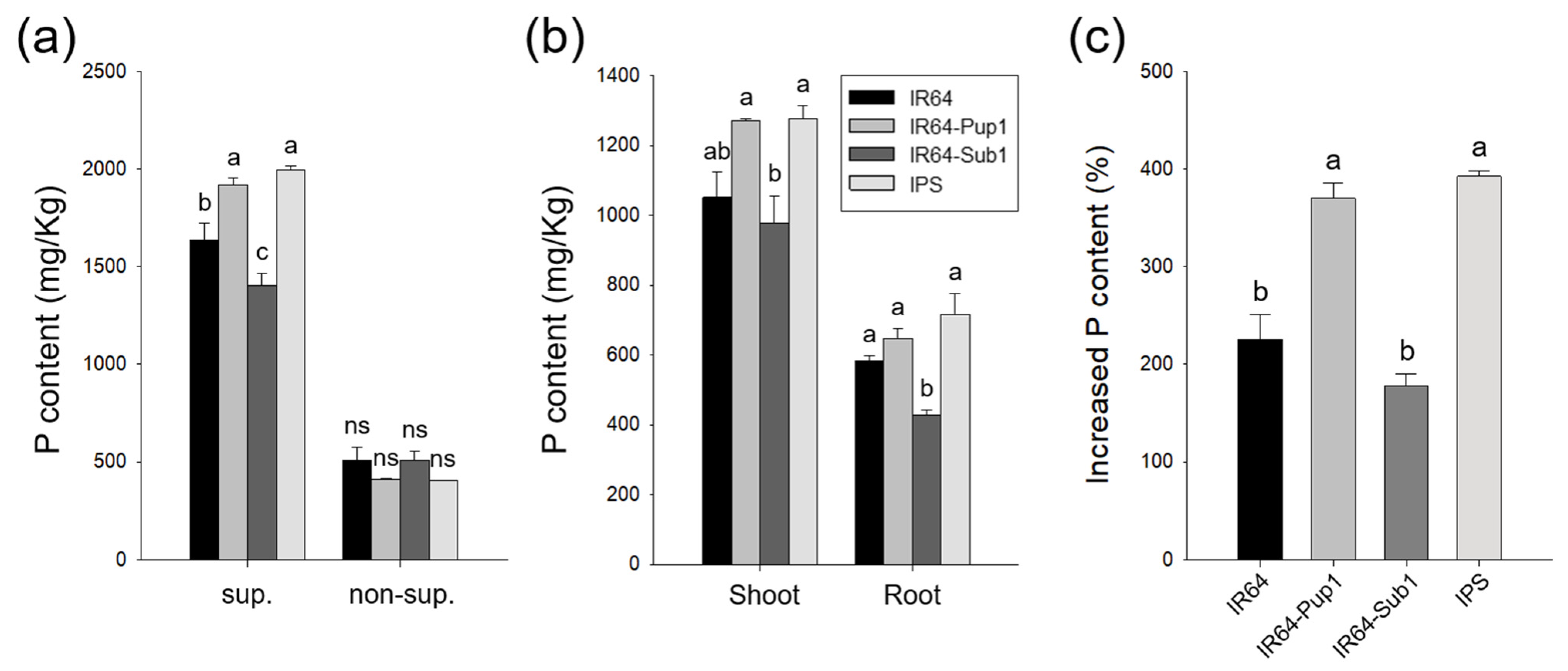
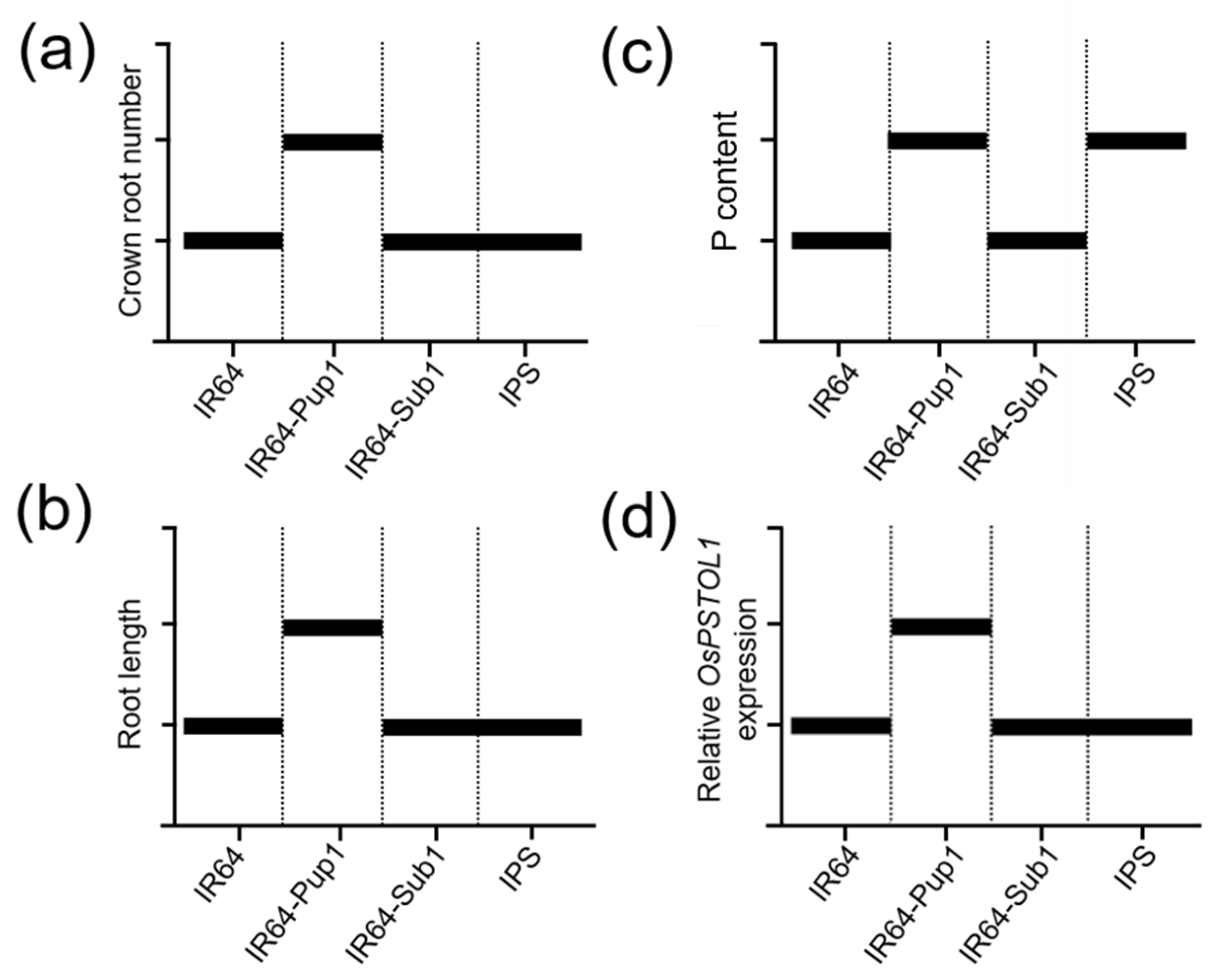
2.3. Pup1 Increased Plant Growth and P Uptake in IPS under P-Supplied Condition
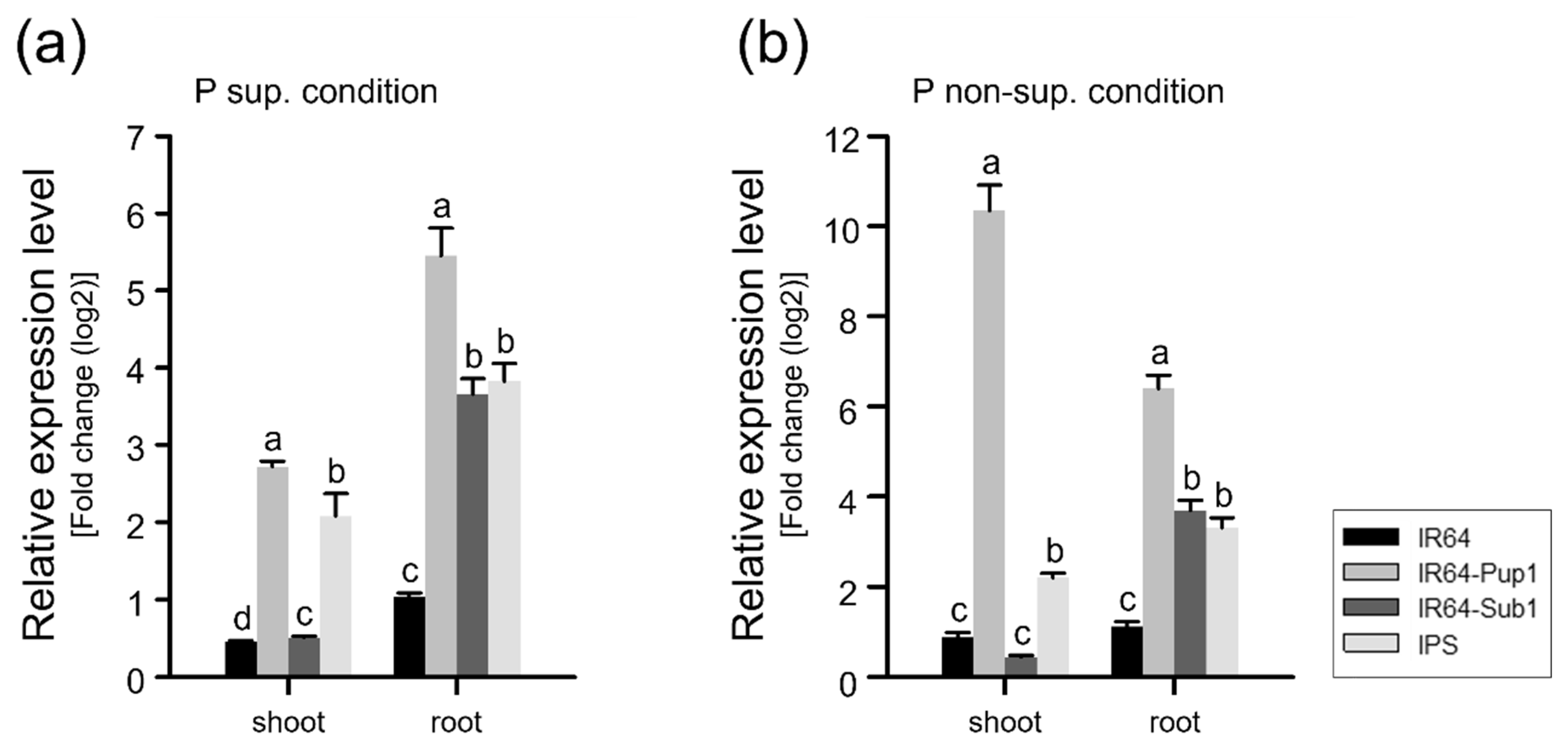
2.4. Interaction of Pup1 with Sub1 Repressed OsPSTOL1 Expression in IPS under Mild Drought Condition
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Plant Materials
5.2. Genomic DNA Extraction and Genomic Analysis
5.3. Submergence Stress Sensitivity Screening System
5.4. Phenotyping under Two Different Phosphorus Concentration Conditions in Soils
5.5. RNA Extraction and Analysis of Transcriptional Levels
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Emerick, K.; Ronald, P.C. Sub1 rice: Engineering rice for climate change. Cold Spring Harb. Perspect. Biol. 2019, 11, a034637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seck, P.A.; Diagne, A.; Mohanty, S.; Wopereis, M.C. Crops that feed the world 7: Rice. Food Secur. 2012, 4, 7–24. [Google Scholar] [CrossRef]
- Heuer, S.; Lu, X.; Chin, J.H.; Tanaka, J.P.; Kanamori, H.; Matsumoto, T.; De Leon, T.; Ulat, V.J.; Ismail, A.M.; Yano, M. Comparative sequence analyses of the major quantitative trait locus phosphorus uptake 1 (Pup1) reveal a complex genetic structure. Plant Biotechnol. J. 2009, 7, 456–471. [Google Scholar] [CrossRef] [PubMed]
- Chankaew, S.; Monkham, T.; Pinta, W.; Sanitchon, J.; Kaewpradit, W.; Srinives, P. Screening tolerance to phosphorus deficiency and validation of phosphorus uptake 1 (Pup1) gene-linked markers in Thai indigenous upland rice germplasm. Agronomy 2019, 9, 81. [Google Scholar] [CrossRef] [Green Version]
- Kirk, G.; George, T.; Courtois, B.; Senadhira, D. Opportunities to improve phosphorus efficiency and soil fertility in rainfed lowland and upland rice ecosystems. Field Crop. Res. 1998, 56, 73–92. [Google Scholar] [CrossRef]
- Shimizu, A.; Kato, K.; Komatsu, A.; Motomura, K.; Ikehashi, H. Genetic analysis of root elongation induced by phosphorus deficiency in rice (Oryza sativa L.): Fine QTL mapping and multivariate analysis of related traits. Theor. Appl. Genet. 2008, 117, 987–996. [Google Scholar] [CrossRef]
- Shimizu, A.; Yanagihara, S.; Kawasaki, S.; Ikehashi, H. Phosphorus deficiency-induced root elongation and its QTL in rice (Oryza sativa L.). Theor. Appl. Genet. 2004, 109, 1361–1368. [Google Scholar] [CrossRef]
- Cordell, D.; Drangert, J.-O.; White, S. The story of phosphorus: Global food security and food for thought. Glob. Environ. Chang. 2009, 19, 292–305. [Google Scholar] [CrossRef]
- Steen, I. Phosphorus availability in the 21st century: Management of a non-renewable resource. Phosphorus Potassium 1998, 217, 25–31. [Google Scholar]
- Swamy, H.M.; Anila, M.; Kale, R.R.; Bhadana, V.; Anantha, M.; Brajendra, P.; Hajira, S.; Balachiranjeevi, C.; Prasanna, B.L.; Pranathi, K. Phenotypic and molecular characterization of rice germplasm lines and identification of novel source for low soil phosphorus tolerance in rice. Euphytica 2019, 215, 118. [Google Scholar] [CrossRef]
- Chin, J.H.; Gamuyao, R.; Dalid, C.; Bustamam, M.; Prasetiyono, J.; Moeljopawiro, S.; Wissuwa, M.; Heuer, S. Developing rice with high yield under phosphorus deficiency: Pup1 sequence to application. Plant Physiol. 2011, 156, 1202–1216. [Google Scholar] [CrossRef] [Green Version]
- Wissuwa, M.; Wegner, J.; Ae, N.; Yano, M. Substitution mapping of Pup1: A major QTL increasing phosphorus uptake of rice from a phosphorus-deficient soil. Theor. Appl. Genet. 2002, 105, 890–897. [Google Scholar] [CrossRef] [PubMed]
- Wissuwa, M.; Yano, M.; Ae, N. Mapping of QTLs for phosphorus-deficiency tolerance in rice (Oryza sativa L.). Theor. Appl. Genet. 1998, 97, 777–783. [Google Scholar] [CrossRef]
- Shin, N.-H.; Han, J.-H.; Jang, S.; Song, K.; Koh, H.-J.; Lee, J.-H.; Yoo, S.; Chin, J.H. Early vigor of a pyramiding line containing two quantitative trait loci, phosphorus uptake 1 (Pup1) and anaerobic germination 1 (ag1) in rice (O. sativa L.). Agriculture 2020, 10, 453. [Google Scholar] [CrossRef]
- Chin, J.H.; Lu, X.; Haefele, S.M.; Gamuyao, R.; Ismail, A.; Wissuwa, M.; Heuer, S. Development and application of gene-based markers for the major rice QTL Phosphorus uptake 1. Theor. Appl. Genet. 2010, 120, 1073–1086. [Google Scholar] [CrossRef]
- Gamuyao, R.; Chin, J.H.; Pariasca-Tanaka, J.; Pesaresi, P.; Catausan, S.; Dalid, C.; Slamet-Loedin, I.; Tecson-Mendoza, E.M.; Wissuwa, M.; Heuer, S. The protein kinase Pstol1 from traditional rice confers tolerance of phosphorus deficiency. Nature 2012, 488, 535–539. [Google Scholar] [CrossRef] [PubMed]
- Wissuwa, M.; Ae, N. Genotypic variation for tolerance to phosphorus deficiency in rice and the potential for its exploitation in rice improvement. Plant Breed. 2001, 120, 43–48. [Google Scholar] [CrossRef]
- Nandi, S.; Subudhi, P.; Senadhira, D.; Manigbas, N.; Sen-Mandi, S.; Huang, N. Mapping QTLs for submergence tolerance in rice by AFLP analysis and selective genotyping. Mol. Gen. Genet. MGG 1997, 255, 1–8. [Google Scholar] [CrossRef]
- Toojinda, T.; Siangliw, M.; Tragoonrung, S.; Vanavichit, A. Molecular genetics of submergence tolerance in rice: QTL analysis of key traits. Ann. Bot. 2003, 91, 243–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, K.; Mackill, D.J. A major locus for submergence tolerance mapped on rice chromosome 9. Mol. Breed. 1996, 2, 219–224. [Google Scholar] [CrossRef]
- Xu, K.; Xu, X.; Ronald, P.; Mackill, D. A high-resolution linkage map of the vicinity of the rice submergence tolerance locus Sub1. Mol. Gen. Genet. MGG 2000, 263, 681–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Septiningsih, E.M.; Pamplona, A.M.; Sanchez, D.L.; Neeraja, C.N.; Vergara, G.V.; Heuer, S.; Ismail, A.M.; Mackill, D.J. Development of submergence-tolerant rice cultivars: The Sub1 locus and beyond. Ann. Bot. 2008, 103, 151–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, K.; Xu, X.; Fukao, T.; Canlas, P.; Maghirang-Rodriguez, R.; Heuer, S.; Ismail, A.M.; Bailey-Serres, J.; Ronald, P.C.; Mackill, D.J. Sub1A is an ethylene-response-factor-like gene that confers submergence tolerance to rice. Nature 2006, 442, 705–708. [Google Scholar] [CrossRef] [Green Version]
- Kuanar, S.R.; Molla, K.A.; Chattopadhyay, K.; Sarkar, R.K.; Mohapatra, P.K. Introgression of Sub1 (SUB1) QTL in mega rice cultivars increases ethylene production to the detriment of grain-filling under stagnant flooding. Sci. Rep. 2019, 9, 18567. [Google Scholar] [CrossRef] [Green Version]
- Panda, D.; Barik, J.; Sarkar, R.K. Recent advances of genetic resources, genes and genetic approaches for flooding tolerance in rice. Curr. Genom. 2021, 22, 41–58. [Google Scholar] [CrossRef] [PubMed]
- Leridon, H. World population outlook: Explosion or implosion? Popul. Soc. 2020, 573, 1–4. [Google Scholar]
- Chen, T.; Shabala, S.; Niu, Y.; Chen, Z.-H.; Shabala, L.; Meinke, H.; Venkataraman, G.; Pareek, A.; Xu, J.; Zhou, M. Molecular mechanisms of salinity tolerance in rice. Crop J. 2021, 9, 506–520. [Google Scholar] [CrossRef]
- Das, K.K.; Panda, D.; Sarkar, R.K.; Reddy, J.; Ismail, A.M. Submergence tolerance in relation to variable floodwater conditions in rice. Environ. Exp. Bot. 2009, 66, 425–434. [Google Scholar] [CrossRef]
- Singh, H.P.; Singh, B.B.; Ram, P.C. Submergence tolerance of rainfed lowland rice: Search for physiological marker traits. J. Plant Physiol. 2001, 158, 883–889. [Google Scholar] [CrossRef]
- Wissuwa, M.; Gamat, G.; Ismail, A.M. Is root growth under phosphorus deficiency affected by source or sink limitations? J. Exp. Bot. 2005, 56, 1943–1950. [Google Scholar] [CrossRef] [Green Version]
- Jacob, J.; Lawlor, D. In vivo photosynthetic electron transport does not limit photosynthetic capacity in phosphate-deficient sunflower and maize leaves. Plant Cell Environ. 1993, 16, 785–795. [Google Scholar] [CrossRef]
- Gahoonia, T.S.; Nielsen, N.E. Variation in root hairs of barley cultivars doubled soil phosphorus uptake. Euphytica 1997, 98, 177–182. [Google Scholar] [CrossRef]
- Lynch, J.P.; Brown, K.M. Topsoil foraging–an architectural adaptation of plants to low phosphorus availability. Plant Soil 2001, 237, 225–237. [Google Scholar] [CrossRef]
- Assuero, S.; Mollier, A.; Pellerin, S. The decrease in growth of phosphorus-deficient maize leaves is related to a lower cell production. Plant Cell Environ. 2004, 27, 887–895. [Google Scholar] [CrossRef]
- Huguenin-Elie, O.; Kirk, G.; Frossard, E. Phosphorus uptake by rice from soil that is flooded, drained or flooded then drained. Eur. J. Soil Sci. 2003, 54, 77–90. [Google Scholar] [CrossRef]
- Kumar, A.; Sandhu, N.; Dixit, S.; Yadav, S.; Swamy, B.; Shamsudin, N.A.A. Marker-assisted selection strategy to pyramid two or more QTLs for quantitative trait-grain yield under drought. Rice 2018, 11, 35. [Google Scholar] [CrossRef] [Green Version]
- Dixit, S.; Swamy, B.M.; Vikram, P.; Ahmed, H.; Cruz, M.S.; Amante, M.; Atri, D.; Leung, H.; Kumar, A. Fine mapping of QTLs for rice grain yield under drought reveals sub-QTLs conferring a response to variable drought severities. Theor. Appl. Genet. 2012, 125, 155–169. [Google Scholar] [CrossRef]
- Dixit, S.; Swamy, B.M.; Vikram, P.; Bernier, J.; Cruz, M.S.; Amante, M.; Atri, D.; Kumar, A. Increased drought tolerance and wider adaptability of qDTY 12.1 conferred by its interaction with qDTY 2.3 and qDTY 3.2. Mol. Breed. 2012, 30, 1767–1779. [Google Scholar] [CrossRef]
- Venuprasad, R.; Bool, M.; Quiatchon, L.; Cruz, M.S.; Amante, M.; Atlin, G. A large-effect QTL for rice grain yield under upland drought stress on chromosome 1. Mol. Breed. 2012, 30, 535–547. [Google Scholar] [CrossRef]
- Venuprasad, R.; Dalid, C.; Del Valle, M.; Zhao, D.; Espiritu, M.; Cruz, M.S.; Amante, M.; Kumar, A.; Atlin, G. Identification and characterization of large-effect quantitative trait loci for grain yield under lowland drought stress in rice using bulk-segregant analysis. Theor. Appl. Genet. 2009, 120, 177–190. [Google Scholar] [CrossRef]
- Vikram, P.; Swamy, B.M.; Dixit, S.; Singh, R.; Singh, B.P.; Miro, B.; Kohli, A.; Henry, A.; Singh, N.; Kumar, A. Drought susceptibility of modern rice varieties: An effect of linkage of drought tolerance with undesirable traits. Sci. Rep. 2015, 5, 14799. [Google Scholar] [CrossRef] [Green Version]
- Vikram, P.; Swamy, B.M.; Dixit, S.; Trinidad, J.; Sta Cruz, M.T.; Maturan, P.C.; Amante, M.; Kumar, A. Linkages and interactions analysis of major effect drought grain yield QTLs in rice. PLoS ONE 2016, 11, e0151532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.; Crouch, J.H. Marker-assisted selection in plant breeding: From publications to practice. Crop Sci. 2008, 48, 391–407. [Google Scholar] [CrossRef] [Green Version]
- Bernier, J.; Kumar, A.; Ramaiah, V.; Spaner, D.; Atlin, G. A large-effect QTL for grain yield under reproductive-stage drought stress in upland rice. Crop Sci. 2007, 47, 507–516. [Google Scholar] [CrossRef]
- Murray, M.; Thompson, W.F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Res. 1980, 8, 4321–4326. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.-W.; Nam, J.; Chu, S.-H.; Phitaktansakul, R.; Yoo, J.-M.; Kang, J.-S.; Min, M.-H.; Cheng, L.; Cao, Y.; Aung, K.M. Development of KNU Axiom Oryza 580K Genotyping Array. In Proceedings of the KSBS& SABRAO International Conference on Plant Breeding for Sustainable Development, Gwangju, Korea, 2–5 July 2019; p. 158. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shin, N.-H.; Lee, O.N.; Han, J.-H.; Song, K.; Koh, H.-J.; Yoo, S.-C.; Chin, J.H. The Effect of Water Level in Rice Cropping System on Phosphorus Uptake Activity of Pup1 in a Pup1+Sub1 Breeding Line. Plants 2021, 10, 1523. https://doi.org/10.3390/plants10081523
Shin N-H, Lee ON, Han J-H, Song K, Koh H-J, Yoo S-C, Chin JH. The Effect of Water Level in Rice Cropping System on Phosphorus Uptake Activity of Pup1 in a Pup1+Sub1 Breeding Line. Plants. 2021; 10(8):1523. https://doi.org/10.3390/plants10081523
Chicago/Turabian StyleShin, Na-Hyun, O New Lee, Jae-Hyuk Han, Kihwan Song, Hee-Jong Koh, Soo-Cheul Yoo, and Joong Hyoun Chin. 2021. "The Effect of Water Level in Rice Cropping System on Phosphorus Uptake Activity of Pup1 in a Pup1+Sub1 Breeding Line" Plants 10, no. 8: 1523. https://doi.org/10.3390/plants10081523