
Efficient CRISPR/Cas-Mediated Targeted Mutagenesis in Spring and Winter Wheat Varieties

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Target Sites
2.2. Plasmid Construction
2.2.1. TaBAK1-2
2.2.2. Ta-eIF4E/Ta-eIF(iso)4E
2.3. Growth of Donor Plants
2.4. Transformation
2.5. Analysis of CRISPR/Cas-Induced Mutations
2.5.1. TaBAK1-2
2.5.2. Ta-eIF4E and Ta-eIF(iso)4E
3. Results
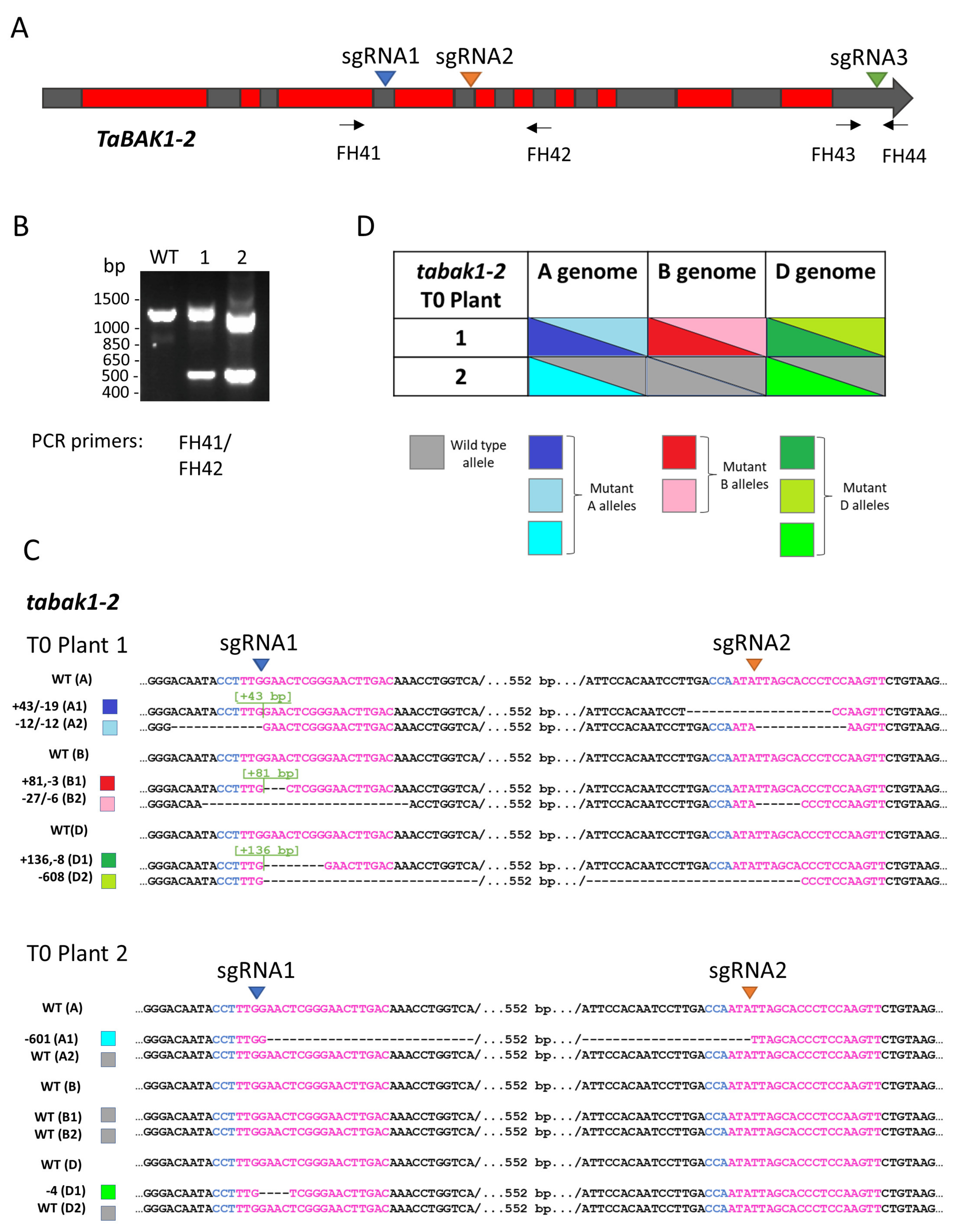
3.1. Targeted Mutagenesis of the TaBAK1-2 Gene
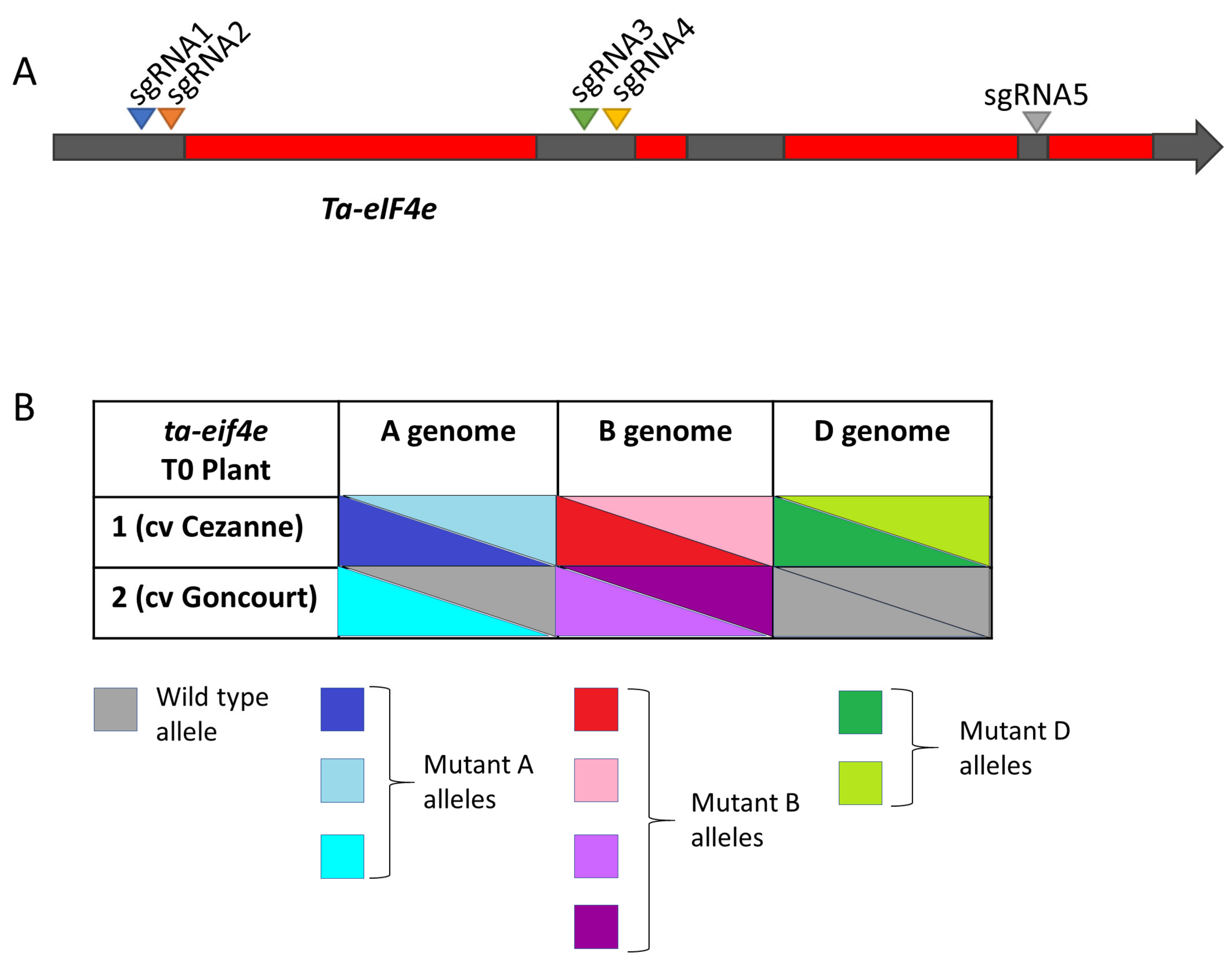
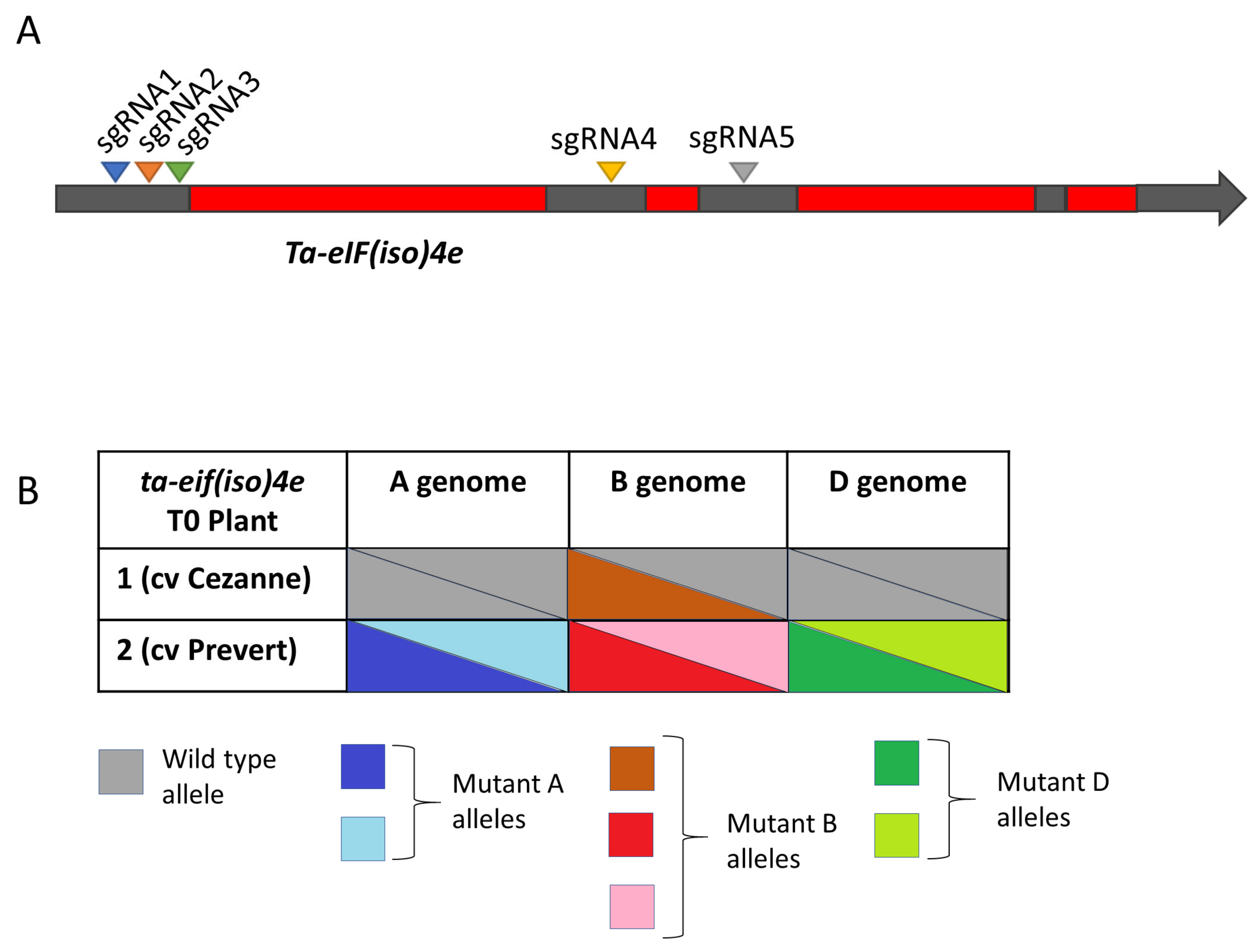
3.2. Targeted Mutagenesis of the Ta-eIF4E and Ta-eIF(iso)4E Genes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Reynolds, M.; Atkin, O.K.; Bennett, M.; Cooper, M.; Dodd, I.C.; Foulkes, M.J.; Frohberg, C.; Hammer, G.; Henderson, I.R.; Huang, B.; et al. Addressing Research Bottlenecks to Crop Productivity. Trends Plant Sci. 2021, 26, 607–630. [Google Scholar] [CrossRef]
- Holme, I.B.; Gregersen, P.L.; Brinch-Pedersen, H. Induced Genetic Variation in Crop Plants by Random or Targeted Mutagenesis: Convergence and Differences. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef]
- Kumar, S.; Rymarquis, L.A.; Ezura, H.; Nekrasov, V. Editorial: CRISPR-Cas in Agriculture: Opportunities and Challenges. Front. Plant Sci. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Zhang, C.; Li, J.; Yan, L.; Wang, N.; Xia, L. Present and Future Prospects for Wheat Improvement through Genome Editing and Advanced Technologies. Plant Commun. 2021, 2, 100211. [Google Scholar] [CrossRef]
- Yasuda, S.; Okada, K.; Saijo, Y. A Look at Plant Immunity through the Window of the Multitasking Coreceptor BAK1. Curr. Opin. Plant Biol. 2017, 38, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Nolan, T.M.; Vukašinović, N.; Liu, D.; Russinova, E.; Yin, Y. Brassinosteroids: Multidimensional Regulators of Plant Growth, Development, and Stress Responses. Plant Cell 2020, 32, 295–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, C.; Kan, J.; Ordon, F.; Perovic, D.; Yang, P. Bymovirus-Induced Yellow Mosaic Diseases in Barley and Wheat: Viruses, Genetic Resistances and Functional Aspects. Theor. Appl. Genet. 2020, 133, 1623–1640. [Google Scholar] [CrossRef] [PubMed]
- Revers, F.; García, J.A. Chapter Three—Molecular Biology of Potyviruses. In Advances in Virus Research; Maramorosch, K., Mettenleiter, T.C., Eds.; Academic Press: Cambridge, MA, USA, 2015; Volume 92, pp. 101–199. [Google Scholar]
- Chandrasekaran, J.; Brumin, M.; Wolf, D.; Leibman, D.; Klap, C.; Pearlsman, M.; Sherman, A.; Arazi, T.; Gal-On, A. Development of Broad Virus Resistance in Non-Transgenic Cucumber Using CRISPR/Cas9 Technology. Mol. Plant Pathol. 2016, 17, 1140–1153. [Google Scholar] [CrossRef] [Green Version]
- Pyott, D.E.; Sheehan, E.; Molnar, A. Engineering of CRISPR/Cas9-Mediated Potyvirus Resistance in Transgene-Free Arabidopsis Plants. Mol. Plant Pathol. 2016, 17, 1276–1288. [Google Scholar] [CrossRef] [Green Version]
- Gomez, M.A.; Lin, Z.D.; Moll, T.; Chauhan, R.D.; Hayden, L.; Renninger, K.; Beyene, G.; Taylor, N.J.; Carrington, J.C.; Staskawicz, B.J.; et al. Simultaneous CRISPR/Cas9-Mediated Editing of Cassava EIF4E Isoforms NCBP-1 and NCBP-2 Reduces Cassava Brown Streak Disease Symptom Severity and Incidence. Plant Biotechnol. J. 2019, 17, 421–434. [Google Scholar] [CrossRef] [Green Version]
- Schmitt-Keichinger, C. Manipulating Cellular Factors to Combat Viruses: A Case Study From the Plant Eukaryotic Translation Initiation Factors EIF4. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Kanyuka, K.; Druka, A.; Caldwell, D.G.; Tymon, A.; Mccallum, N.; Waugh, R.; Adams, M.J. Evidence That the Recessive Bymovirus Resistance Locus Rym4 in Barley Corresponds to the Eukaryotic Translation Initiation Factor 4E Gene. Mol. Plant Pathol. 2005, 6, 449–458. [Google Scholar] [CrossRef] [PubMed]
- Concordet, J.-P.; Haeussler, M. CRISPOR: Intuitive Guide Selection for CRISPR/Cas9 Genome Editing Experiments and Screens. Nucleic Acids Res. 2018, 46, W242–W245. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Liu, B.; Weeks, D.P.; Spalding, M.H.; Yang, B. Large Chromosomal Deletions and Heritable Small Genetic Changes Induced by CRISPR/Cas9 in Rice. Nucleic Acids Res. 2014, 42, 10903–10914. [Google Scholar] [CrossRef]
- Zhang, Z.; Hua, L.; Gupta, A.; Tricoli, D.; Edwards, K.J.; Yang, B.; Li, W. Development of an Agrobacterium-Delivered CRISPR/Cas9 System for Wheat Genome Editing. Plant Biotechnol. J. 2019, 17, 1623–1635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alotaibi, S.S.; Sparks, C.A.; Parry, M.A.J.; Simkin, A.J.; Raines, C.A. Identification of Leaf Promoters for Use in Transgenic Wheat. Plants 2018, 7, 27. [Google Scholar] [CrossRef] [Green Version]
- Xie, K.; Minkenberg, B.; Yang, Y. Boosting CRISPR/Cas9 Multiplex Editing Capability with the Endogenous TRNA-Processing System. Proc. Natl. Acad. Sci. USA 2015, 112, 3570–3575. [Google Scholar] [CrossRef] [Green Version]
- Dang, Y.; Jia, G.; Choi, J.; Ma, H.; Anaya, E.; Ye, C.; Shankar, P.; Wu, H. Optimizing SgRNA Structure to Improve CRISPR-Cas9 Knockout Efficiency. Genome Biol. 2015, 16, 280. [Google Scholar] [CrossRef] [Green Version]
- Hahn, F.; Korolev, A.; Sanjurjo Loures, L.; Nekrasov, V. A Modular Cloning Toolkit for Genome Editing in Plants. BMC Plant Biol. 2020, 20, 179. [Google Scholar] [CrossRef] [Green Version]
- Sparks, C.A.; Doherty, A. Genetic Transformation of Common Wheat (Triticum aestivum L.) Using Biolistics. In Biolistic DNA Delivery in Plants: Methods and Protocols; Rustgi, S., Luo, H., Eds.; Methods in Molecular Biology; Springer US: New York, NY, USA, 2020; pp. 229–250. ISBN 978-1-07-160356-7. [Google Scholar]
- Nekrasov, V.; Wang, C.; Win, J.; Lanz, C.; Weigel, D.; Kamoun, S. Rapid Generation of a Transgene-Free Powdery Mildew Resistant Tomato by Genome Deletion. Sci. Rep. 2017, 7, 482. [Google Scholar] [CrossRef] [Green Version]
- Andersson, M.; Turesson, H.; Nicolia, A.; Falt, A.S.; Samuelsson, M.; Hofvander, P. Efficient Targeted Multiallelic Mutagenesis in Tetraploid Potato (Solanum Tuberosum) by Transient CRISPR-Cas9 Expression in Protoplasts. Plant Cell Rep. 2017, 36, 117–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersson, M.; Turesson, H.; Olsson, N.; Fält, A.-S.; Ohlsson, P.; Gonzalez, M.N.; Samuelsson, M.; Hofvander, P. Genome Editing in Potato via CRISPR-Cas9 Ribonucleoprotein Delivery. Physiol. Plant. 2018, 164, 378–384. [Google Scholar] [CrossRef] [Green Version]
- Banakar, R.; Eggenberger, A.L.; Lee, K.; Wright, D.A.; Murugan, K.; Zarecor, S.; Lawrence-Dill, C.J.; Sashital, D.G.; Wang, K. High-Frequency Random DNA Insertions upon Co-Delivery of CRISPR-Cas9 Ribonucleoprotein and Selectable Marker Plasmid in Rice. Sci. Rep. 2019, 9, 19902. [Google Scholar] [CrossRef]
- Li, J.; Jiao, G.; Sun, Y.; Chen, J.; Zhong, Y.; Yan, L.; Jiang, D.; Ma, Y.; Xia, L. Modification of Starch Composition, Structure and Properties through Editing of TaSBEIIa in Both Winter and Spring Wheat Varieties by CRISPR/Cas9. Plant Biotechnol. J. 2020, 19, 937–951. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Luo, W.; Linghu, Q.; Abe, F.; Hisano, H.; Sato, K.; Kamiya, Y.; Kawaura, K.; Onishi, K.; Endo, M.; et al. In Planta Genome Editing in Commercial Wheat Varieties. Front. Plant Sci. 2021, 12. [Google Scholar] [CrossRef]
- Grützner, R.; Martin, P.; Horn, C.; Mortensen, S.; Cram, E.J.; Lee-Parsons, C.W.T.; Stuttmann, J.; Marillonnet, S. High-Efficiency Genome Editing in Plants Mediated by a Cas9 Gene Containing Multiple Introns. Plant Commun. 2021, 2, 100135. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TaBAK1-2 | sgRNA1 | GTCAAGTTCCCGAGTTCCAA |
| sgRNA2 | AACTTGGAGGGTGCTAATAT | |
| sgRNA3 | GATCCAGTCGTTGTTTCGCG | |
| Ta-eIF4E | sgRNA1 | GCTCCCACATTCAACTTGCT |
| sgRNA2 | GTTGTCGAACCAGAAGGTCC | |
| sgRNA3 | GAAGGTGTGGATGGGGTGGA | |
| sgRNA4 | GATGGTCCATTTACCGCCAT | |
| sgRNA5 | GAAGGAGTTTCTGGACTACA | |
| Ta-eIF(iso)4E | sgRNA1 | GAACTCTTCGACGGTGTCGA |
| sgRNA2 | GGCTGGGGTAGAACCAAAGT | |
| sgRNA3 | GACAGGATAAGCTTTCATTA | |
| sgRNA4 | GGTCTGGATGTCGTACCAGA | |
| sgRNA5 | GGTCGAAGCTGCGCTCCCGG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hahn, F.; Sanjurjo Loures, L.; Sparks, C.A.; Kanyuka, K.; Nekrasov, V. Efficient CRISPR/Cas-Mediated Targeted Mutagenesis in Spring and Winter Wheat Varieties. Plants 2021, 10, 1481. https://doi.org/10.3390/plants10071481
Hahn F, Sanjurjo Loures L, Sparks CA, Kanyuka K, Nekrasov V. Efficient CRISPR/Cas-Mediated Targeted Mutagenesis in Spring and Winter Wheat Varieties. Plants. 2021; 10(7):1481. https://doi.org/10.3390/plants10071481
Chicago/Turabian StyleHahn, Florian, Laura Sanjurjo Loures, Caroline A. Sparks, Kostya Kanyuka, and Vladimir Nekrasov. 2021. "Efficient CRISPR/Cas-Mediated Targeted Mutagenesis in Spring and Winter Wheat Varieties" Plants 10, no. 7: 1481. https://doi.org/10.3390/plants10071481