
Brassinolide Enhances the Level of Brassinosteroids, Protein, Pigments, and Monosaccharides in Wolffia arrhiza Treated with Brassinazole


Abstract
:1. Introduction
2. Results
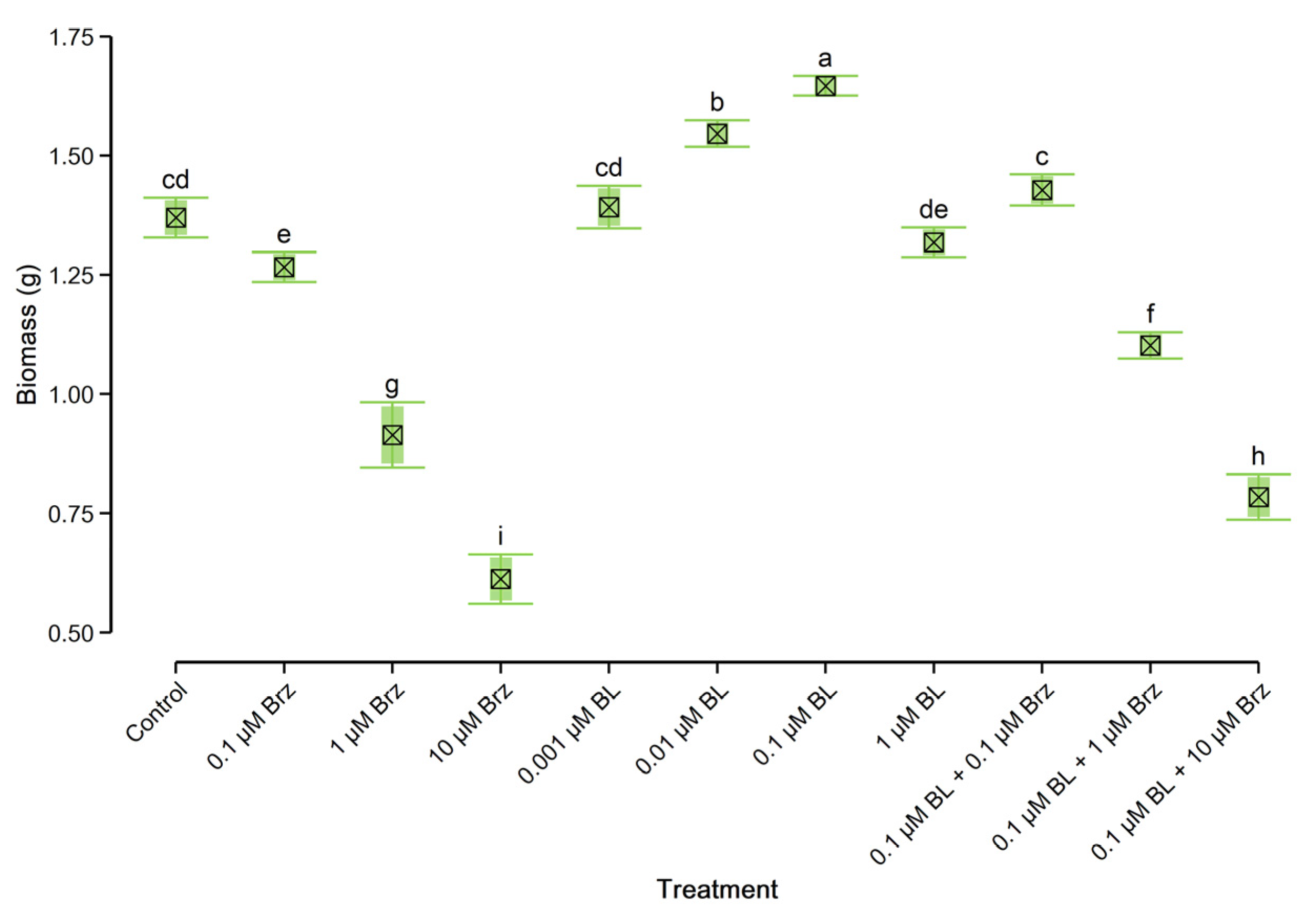
2.1. Growth Rate of W. arrhiza
2.2. Brassinazole and Brassinosteroids’ Content in W. arrhiza
2.3. Photosynthetic Pigments’ Content in W. arrhiza
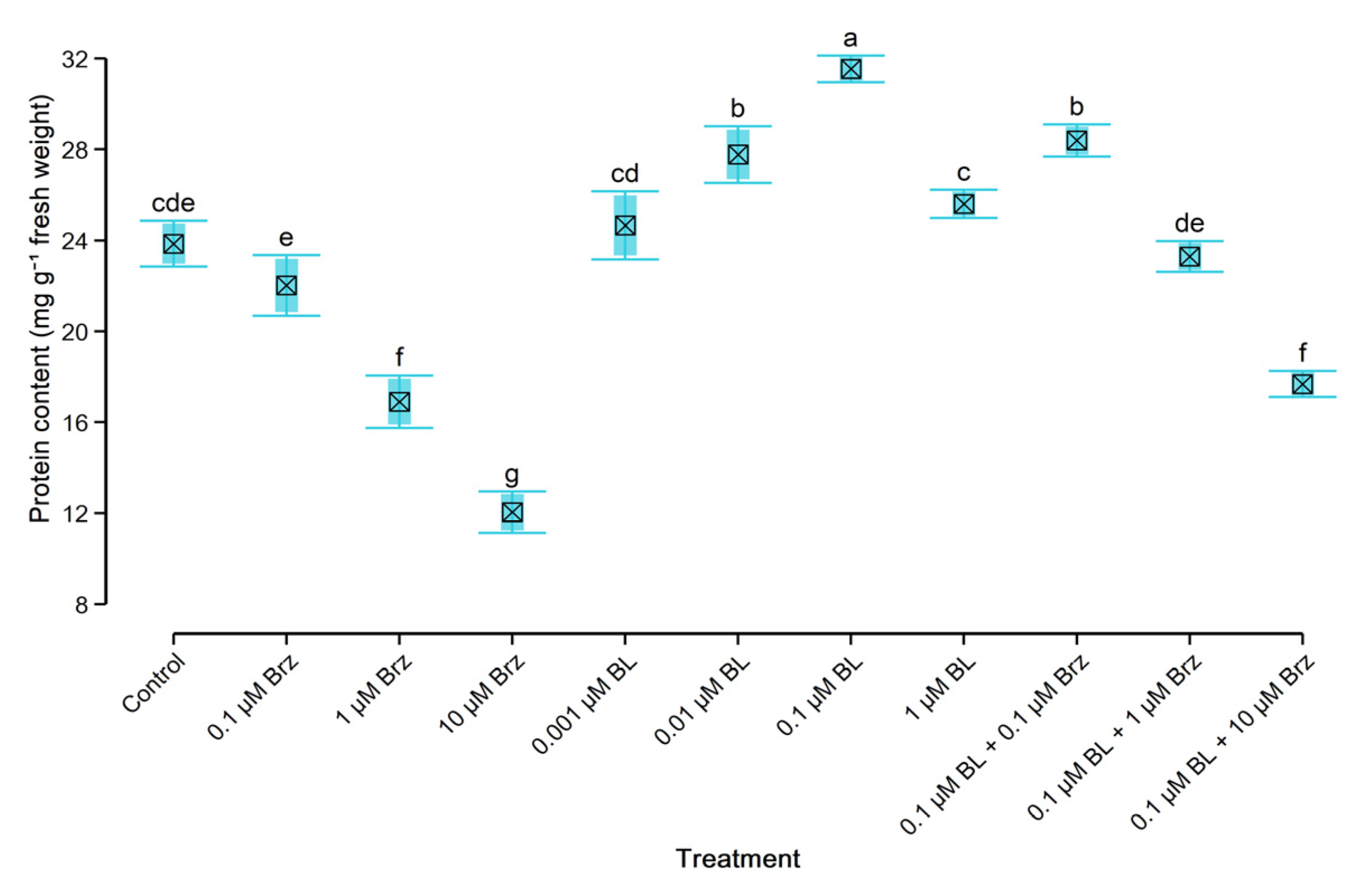
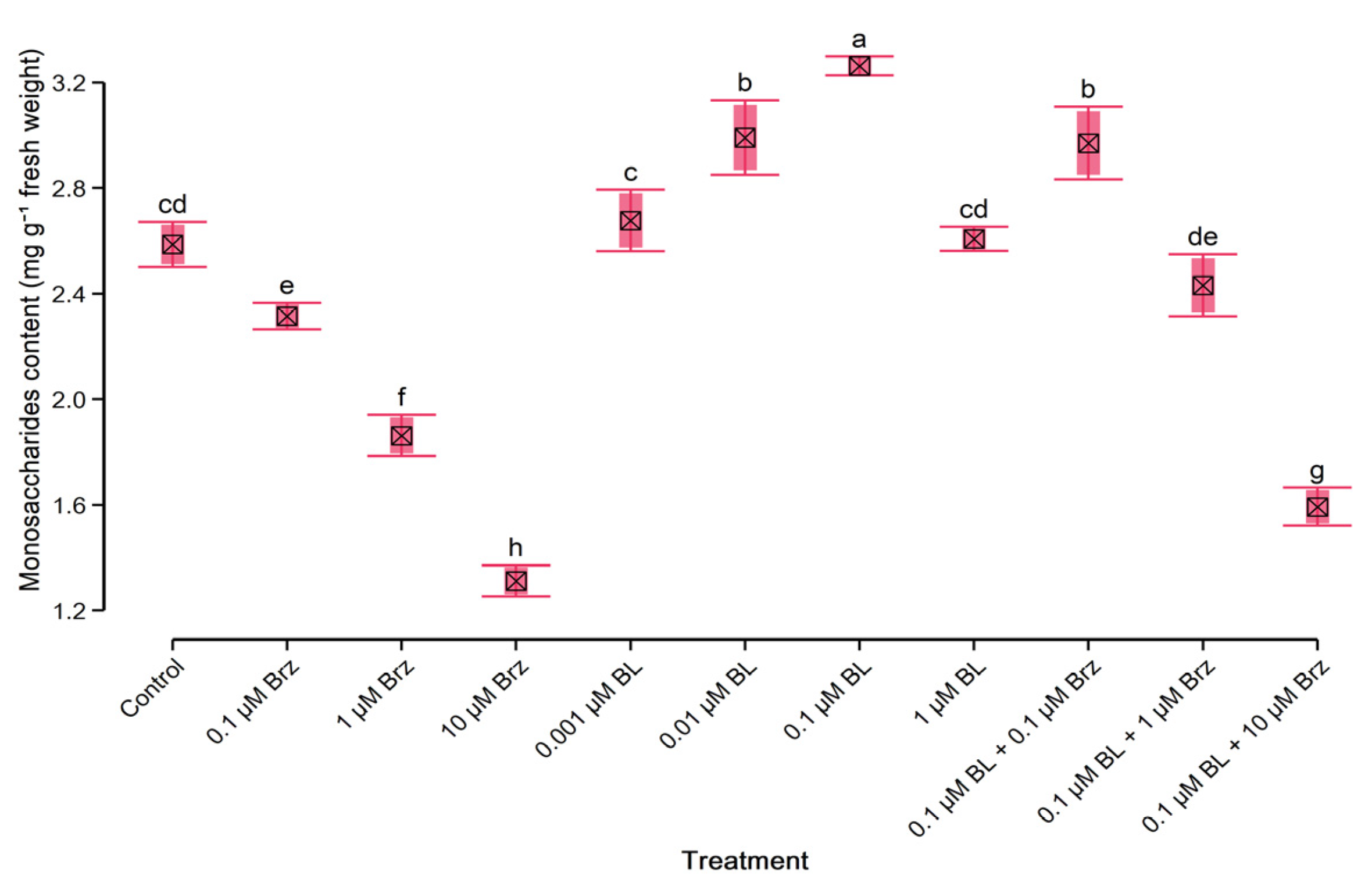
2.4. Protein and Monosaccharides’ Content in W. arrhiza
3. Discussion
3.1. Exogenous Brassinolide Improves the Growth Rate and Content of Endogenous Hormones and Primary Metabolites
3.2. Exogenous Brassinolide Overcame the Negative Effect of Brassinazole
4. Materials and Methods
4.1. Plant Material and Growth Conditions
4.2. Chemicals
4.3. Quantification of Brassinazole and Brassinosteroids
4.4. Quantification of Photosynthetic Pigments
4.5. Determination of Proteins’ Content
4.6. Determination of Monosaccharides’ Content
4.7. Statistical Treatment
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zullo, M.A.T.; Bajguz, A. The brassinosteroids family—structural diversity of natural compounds and their precursors. In Brassinosteroids: Plant Growth and Development; Hayat, S., Yusuf, M., Bhardwaj, R., Bajguz, A., Eds.; Springer: Singapore, 2019; pp. 1–44. [Google Scholar] [CrossRef]
- Bajguz, A. Brassinosteroids in microalgae: Application for growth improvement and protection against abiotic stresses. In Brassinosteroids: Plant Growth and Development; Hayat, S., Yusuf, M., Bhardwaj, R., Bajguz, A., Eds.; Springer: Singapore, 2019; pp. 45–58. [Google Scholar] [CrossRef]
- Tarkowska, D.; Strnad, M. Isoprenoid-derived plant signaling molecules: Biosynthesis and biological importance. Planta 2018, 247, 1051–1066. [Google Scholar] [CrossRef]
- Bajguz, A.; Tretyn, A. The chemical characteristic and distribution of brassinosteroids in plants. Phytochemistry 2003, 62, 1027–1046. [Google Scholar] [CrossRef]
- Kanwar, M.K.; Bajguz, A.; Zhou, J.; Bhardwaj, R. Analysis of brassinosteroids in plants. J. Plant Growth Regul. 2017, 36, 1002–1030. [Google Scholar] [CrossRef]
- Grove, M.D.; Spencer, G.F.; Rohwedder, W.K.; Mandava, N.; Worley, J.F.; Warthen, J.D., Jr.; Steffens, G.L.; Flippen-Anderson, J.L.; Cook, J.C., Jr. Brassinolide, a plant growth-promoting steroid isolated from Brassica napus pollen. Nature 1979, 281, 216–217. [Google Scholar] [CrossRef]
- Nolan, T.M.; Vukašinović, N.; Liu, D.; Russinova, E.; Yin, Y. Brassinosteroids: Multidimensional regulators of plant growth, development, and stress responses. Plant Cell 2020, 32, 295–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siddiqui, H.; Hayat, S.; Bajguz, A. Regulation of photosynthesis by brassinosteroids in plants. Acta Physiol. Plant. 2018, 40, 59. [Google Scholar] [CrossRef]
- Hayat, S.; Mori, M.; Fariduddin, Q.; Bajguz, A.; Ahmad, A. Physiological role of brassinosteroids: An update. Indian J. Plant Physiol. 2010, 15, 99–109. [Google Scholar]
- Li, Z.; He, Y. Roles of brassinosteroids in plant reproduction. Int. J. Mol. Sci. 2020, 21, 872. [Google Scholar] [CrossRef] [Green Version]
- Hussain, M.A.; Fahad, S.; Sharif, R.; Jan, M.F.; Mujtaba, M.; Ali, Q.; Ahmad, A.; Ahmad, H.; Amin, N.; Ajayo, B.S.; et al. Multifunctional role of brassinosteroid and its analogues in plants. Plant Growth Regul. 2020, 92, 141–156. [Google Scholar] [CrossRef]
- Bajguz, A.; Hayat, S. Effects of brassinosteroids on the plant responses to environmental stresses. Plant Physiol. Biochem. 2009, 47, 1–8. [Google Scholar] [CrossRef]
- Rajewska, I.; Talarek, M.; Bajguz, A. Brassinosteroids and response of plants to heavy metals action. Front. Plant Sci. 2016, 7, 629. [Google Scholar] [CrossRef] [Green Version]
- Ahanger, M.A.; Ashraf, M.; Bajguz, A.; Ahmad, P. Brassinosteroids regulate growth in plants under stressful environments and crosstalk with other potential phytohormones. J. Plant Growth Regul. 2018, 37, 1007–1024. [Google Scholar] [CrossRef]
- Li, S.; Zheng, H.; Lin, L.; Wang, F.; Sui, N. Roles of brassinosteroids in plant growth and abiotic stress response. Plant Growth Regul. 2020, 93, 29–38. [Google Scholar] [CrossRef]
- Bajguz, A.; Chmur, M.; Gruszka, D. Comprehensive overview of the brassinosteroid biosynthesis pathways: Substrates, products, inhibitors, and connections. Front. Plant Sci. 2020, 11, 1034. [Google Scholar] [CrossRef]
- Wei, Z.; Li, J. Regulation of brassinosteroid homeostasis in higher plants. Front. Plant Sci. 2020, 11, 583622. [Google Scholar] [CrossRef]
- Rozhon, W.; Akter, S.; Fernandez, A.; Poppenberger, B. Inhibitors of brassinosteroid biosynthesis and signal transduction. Molecules 2019, 24, 4372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asami, T.; Min, Y.K.; Nagata, N.; Yamagishi, K.; Takatsuto, S.; Fujioka, S.; Murofushi, N.; Yamaguchi, I.; Yoshida, S. Characterization of brassinazole, a triazole-type brassinosteroid biosynthesis inhibitor. Plant Physiol. 2000, 123, 93–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asami, T.; Yoshida, S. Brassinosteroid biosynthesis inhibitors. Trends Plant Sci. 1999, 4, 348–353. [Google Scholar] [CrossRef]
- Nagata, N.; Asami, T.; Yoshida, S. Brassinazole, an inhibitor of brassinosteroid biosynthesis, inhibits development of secondary xylem in cress plants (Lepidium sativum). Plant Cell Physiol. 2001, 42, 1006–1011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagata, N.; Min, Y.K.; Nakano, T.; Asami, T.; Yoshida, S. Treatment of dark-grown Arabidopsis thaliana with a brassinosteroid-biosynthesis inhibitor, brassinazole, induces some characteristics of light-grown plants. Planta 2000, 211, 781–790. [Google Scholar] [CrossRef] [PubMed]
- Pereira-Netto, A.B.; Roessner, U.; Fujioka, S.; Bacic, A.; Asami, T.; Yoshida, S.; Clouse, S.D. Shooting control by brassinosteroids: Metabolomic analysis and effect of brassinazole on Malus prunifolia, the Marubakaido apple rootstock. Tree Physiol. 2009, 29, 607–620. [Google Scholar] [CrossRef]
- Pramod, S.; Anju, M.; Rajesh, H.; Thulaseedharan, A.; Rao, K.S. Effect of exogenously applied 24-epibrassinolide and brassinazole on xylogenesis and microdistribution of cell wall polymers in Leucaena leucocephala (Lam) De Wit. J. Plant Growth Regul. 2021, in press. [Google Scholar] [CrossRef]
- Terakado, J.; Fujihara, S.; Goto, S.; Kuratani, R.; Suzuki, Y.; Yoshida, S.; Yoneyama, T. Systemic effect of a brassinosteroid on root nodule formation in soybean as revealed by the application of brassinolide and brassinazole. Soil Sci. Plant Nutr. 2005, 51, 389–395. [Google Scholar] [CrossRef]
- Bajguz, A.; Asami, T. Effects of brassinazole, an inhibitor of brassinosteroid biosynthesis, on light- and dark-grown Chlorella vulgaris. Planta 2004, 218, 869–877. [Google Scholar] [CrossRef] [PubMed]
- Chmur, M.; Bajguz, A.; Piotrowska-Niczyporuk, A. Effect of cadmium on the level of isoprenoid-derived phytohormones in duckweed Wolffia arrhiza. J. Plant Growth Regul. 2020, 39, 1518–1530. [Google Scholar] [CrossRef]
- Kotowska, U.; Karpinska, J.; Kapelewska, J.; Kowejsza, E.M.; Piotrowska-Niczyporuk, A.; Piekutin, J.; Kotowski, A. Removal of phthalates and other contaminants from municipal wastewater during cultivation of Wolffia arrhiza. Process Saf. Environ. Prot. 2018, 120, 268–277. [Google Scholar] [CrossRef]
- Piotrowska, A.; Bajguz, A.; Czerpak, R.; Kot, K. Changes in the growth, chemical composition, and antioxidant activity in the aquatic plant Wolffia arrhiza (L.) Wimm. (Lemnaceae) exposed to jasmonic acid. J. Plant Growth Regul. 2010, 29, 53–62. [Google Scholar] [CrossRef]
- Forni, C.; Tommasi, F. Duckweed: A tool for ecotoxicology and a candidate for phytoremediation. Curr. Biotechnol. 2016, 5, 2–10. [Google Scholar] [CrossRef]
- Bajguz, A.; Asami, T. Suppression of Wolffia arrhiza growth by brassinazole, an inhibitor of brassinosteroid biosynthesis and its restoration by endogenous 24-epibrassinolide. Phytochemistry 2005, 66, 1787–1796. [Google Scholar] [CrossRef]
- Li, Y.X.; Deng, T.; Duan, C.F.; Ni, L.X.; Wang, N.; Guan, Y.F. Dispersive matrix solid-phase extraction method coupled with high performance liquid chromatography-tandem mass spectrometry for ultrasensitive quantification of endogenous brassinosteroids in minute plants and its application for geographical distribution study. J. Agric. Food Chem. 2019, 67, 3037–3045. [Google Scholar] [CrossRef] [PubMed]
- Symons, G.M.; Reid, J.B. Brassinosteroids do not undergo long-distance transport in pea. Implications for the regulation of endogenous brassinosteroid levels. Plant Physiol. 2004, 135, 2196–2206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roh, J.; Yeom, H.S.; Jang, H.; Kim, S.; Youn, J.H.; Kim, S.K. Identification and biosynthesis of C-24 ethylidene brassinosteroids in Arabidopsis thaliana. J. Plant Biol. 2017, 60, 533–538. [Google Scholar] [CrossRef]
- Ackerman-Lavert, M.; Savaldi-Goldstein, S. Growth models from a brassinosteroid perspective. Curr. Opin. Plant Biol. 2020, 53, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Kalinich, J.F.; Mandava, N.B.; Todhunter, J.A. Relationship of nucleic acid metabolism to brassinolide-induced responses in beans. J. Plant Physiol. 1986, 125, 345–353. [Google Scholar] [CrossRef]
- Mandava, N.B.; Thompson, M.J.; Yopp, J.H. Effects of selected inhibitors of RNA and protein synthesis on brassinosteroid-induced responses in mung bean epicotyls. J. Plant Physiol. 1987, 128, 53–65. [Google Scholar] [CrossRef]
- Bajguz, A. Effect of brassinosteroids on nucleic acids and protein content in cultured cells of Chlorella vulgaris. Plant Physiol. Biochem. 2000, 38, 209–215. [Google Scholar] [CrossRef]
- Mussig, C. Brassinosteroid-promoted growth. Plant Biol. 2005, 7, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Jiroutova, P.; Mikulik, J.; Novak, O.; Strnad, M.; Oklestkova, J. Brassinosteroids induce strong, dose-dependent inhibition of etiolated pea seedling growth correlated with ethylene production. Biomolecules 2019, 9, 849. [Google Scholar] [CrossRef] [Green Version]
- Tong, H.N.; Xiao, Y.H.; Liu, D.P.; Gao, S.P.; Liu, L.C.; Yin, Y.H.; Jin, Y.; Qian, Q.; Chu, C.C. Brassinosteroid regulates cell elongation by modulating gibberellin metabolism in rice. Plant Cell 2014, 26, 4376–4393. [Google Scholar] [CrossRef] [Green Version]
- Talarek-Karwel, M.; Bajguz, A.; Piotrowska-Niczyporuk, A.; Rajewska, I. The effect of 24-epibrassinolide on the green alga Acutodesmus obliquus (Chlorophyceae). Plant Physiol. Biochem. 2018, 124, 175–183. [Google Scholar] [CrossRef]
- Bajguz, A. An enhancing effect of exogenous brassinolide on the growth and antioxidant activity in Chlorella vulgaris cultures under heavy metals stress. Environ. Exp. Bot. 2010, 68, 175–179. [Google Scholar] [CrossRef]
- Latha, P.; Vardhini, V. Effect of brassinolide on the growth of mustard crops grown in semi-arid tropics of nizamabad. Int. J. Plant Soil Sci. 2016, 9, 1–5. [Google Scholar] [CrossRef]
- Talarek-Karwel, M.; Bajguz, A.; Piotrowska-Niczyporuk, A. Hormonal response of Acutodesmus obliquus exposed to combined treatment with 24-epibrassinolide and lead. J. Appl. Phycol. 2020, 32, 2903–2914. [Google Scholar] [CrossRef]
- Stirk, W.A.; Balint, P.; Tarkowska, D.; Novak, O.; Strnad, M.; Ordog, V.; Van Staden, J. Hormone profiles in microalgae: Gibberellins and brassinosteroids. Plant Physiol. Biochem. 2013, 70, 348–353. [Google Scholar] [CrossRef] [PubMed]
- Bajguz, A. Isolation and characterization of brassinosteroids from algal cultures of Chlorella vulgaris Beijerinck (Trebouxiophyceae). J. Plant Physiol. 2009, 166, 1946–1949. [Google Scholar] [CrossRef] [PubMed]
- Bajguz, A.; Orczyk, W.; Golebiewska, A.; Chmur, M.; Piotrowska-Niczyporuk, A. Occurrence of brassinosteroids and influence of 24-epibrassinolide with brassinazole on their content in the leaves and roots of Hordeum vulgare L. cv. Golden Promise. Planta 2019, 249, 123–137. [Google Scholar] [CrossRef] [Green Version]
- Janeczko, A.; Swaczynova, J. Endogenous brassinosteroids in wheat treated with 24-epibrassinolide. Biol. Plant. 2010, 54, 477–482. [Google Scholar] [CrossRef]
- Patias, L.D.; Fernandes, A.S.; Petry, F.C.; Mercadante, A.Z.; Jacob-Lopes, E.; Zepka, L.Q. Carotenoid profile of three microalgae/cyanobacteria species with peroxyl radical scavenger capacity. Food Res. Int. 2017, 100, 260–266. [Google Scholar] [CrossRef] [PubMed]
- Poojary, M.M.; Barba, F.J.; Aliakbarian, B.; Donsì, F.; Pataro, G.; Dias, D.A.; Juliano, P. Innovative alternative technologies to extract carotenoids from microalgae and seaweeds. Mar. Drugs 2016, 14, 214. [Google Scholar] [CrossRef] [PubMed]
- Czerpak, R.; Dobrzyń, P.; Krotke, A.; Kicińska, E. The effect of auxins and salicylic acid on chlorophyll and carotenoid contents in Wolffia arrhiza (L.) Wimm. (Lemnaceae) growing on media of various trophicities. Pol. J. Environ. Stud. 2002, 11, 231–235. [Google Scholar]
- Piotrowska-Niczyporuk, A.; Bajguz, A.; Kotowska, U.; Bralska, M.; Talarek-Karwel, M. Growth, metabolite profile, oxidative status, and phytohormone levels in the green alga Acutodesmus obliquus exposed to exogenous auxins and cytokinins. J. Plant Growth Regul. 2018, 37, 1159–1174. [Google Scholar] [CrossRef] [Green Version]
- Ortiz, D.; Ponrajan, A.; Bonnet, J.P.; Rocheford, T.; Ferruzzi, M.G. Carotenoid stability during dry milling, storage, and extrusion processing of biofortified maize genotypes. J. Agric. Food Chem. 2018, 66, 4683–4691. [Google Scholar] [CrossRef] [PubMed]
- Paznocht, L.; Burešová, B.; Kotíková, Z.; Martinek, P. Carotenoid content of extruded and puffed products made of colored-grain wheats. Food Chem. 2021, 340, 127951. [Google Scholar] [CrossRef] [PubMed]
- Alamu, E.O.; Olatunde, G.O.; Adegunwa, M.O.; Adebanjo, L.A.; Awoyinfa, O.C.; Soyoye, J.B. Carotenoid profile and functional properties of flour blends from biofortified maize and improved soybean varieties for product developments. Cogent Food Agric. 2021, 7, 1868665. [Google Scholar] [CrossRef]
- Bajguz, A.; Czerpak, R. Physiological and biochemical role of brassinosteroids and their structure-activity relationship in the green alga Chlorella vulgaris Beijerinck (Chlorophyceae). J. Plant Growth Regul. 1998, 17, 131–139. [Google Scholar] [CrossRef]
- Dong, Y.J.; Wang, W.W.; Hu, G.Q.; Chen, W.F.; Zhuge, Y.P.; Wang, Z.L.; He, M.R. Role of exogenous 24-epibrassinolide in enhancing the salt tolerance of wheat seedlings. J. Soil Sci. Plant Nutr. 2017, 17, 554–569. [Google Scholar] [CrossRef] [Green Version]
- Maity, U.; Bera, A. Effect of exogenous application of brassinolide and salicylic acid on certain physiological and biochemical aspects of green gram (Vigna radiata L. Wilczek). Indian J. Agric. Res. 2009, 43, 194–199. [Google Scholar]
- Eckstein, A.; Zięba, P.; Gabryś, H. Sugar and light effects on the condition of the photosynthetic apparatus of Arabidopsis thaliana cultured in vitro. J. Plant Growth Regul. 2012, 31, 90–101. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.Q.; Huang, L.F.; Hu, W.H.; Zhou, Y.H.; Mao, W.H.; Ye, S.F.; Nogués, S. A role for brassinosteroids in the regulation of photosynthesis in Cucumis sativus. J. Exp. Bot. 2004, 55, 1135–1143. [Google Scholar] [CrossRef]
- Ren, C.M.; Han, C.Y.; Peng, W.; Huang, Y.; Peng, Z.H.; Xiong, X.Y.; Zhu, Q.; Gao, B.D.; Xie, D.X. A leaky mutation in DWARF4 reveals an antagonistic role of brassinosteroid in the inhibition of root growth by jasmonate in Arabidopsis. Plant Physiol. 2009, 151, 1412–1420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.J.; Liu, X.Y.; Wang, R.; Li, W.C.; Rodermel, S.; Yu, F. Overexpression of a putative Arabidopsis BAHD acyltransferase causes dwarfism that can be rescued by brassinosteroid. J. Exp. Bot. 2012, 63, 5787–5801. [Google Scholar] [CrossRef] [Green Version]
- Shahnejat-Bushehri, S.; Tarkowska, D.; Sakuraba, Y.; Balazadeh, S. Arabidopsis NAC transcription factor JUB1 regulates GA/BR metabolism and signalling. Nat. Plants 2016, 2, 16013. [Google Scholar] [CrossRef] [PubMed]
- Mazorra, L.M.; Nunez, M.; Napoles, M.C.; Yoshida, S.; Robaina, C.; Coll, F.; Asami, T. Effects of structural analogs of brassinosteroids on the recovery of growth inhibition by a specific brassinosteroid biosynthesis inhibitor. Plant Growth Regul. 2004, 44, 183–185. [Google Scholar] [CrossRef]
- Hutner, S.H. Comparative physiology of heterotrophic growth in plants. In Growth and Differentiation in Plants; Loomis, W.E., Ed.; Iowa State College Press: Ames, IA, USA, 1953; pp. 417–446. [Google Scholar]
- Xin, P.Y.; Yan, J.J.; Fan, J.S.; Chu, J.F.; Yan, C.Y. An improved simplified high-sensitivity quantification method for determining brassinosteroids in different tissues of rice and Arabidopsis. Plant Physiol. 2013, 162, 2056–2066. [Google Scholar] [CrossRef] [Green Version]
- Zapata, M.; Rodríguez, F.; Garrido, J.L. Separation of chlorophylls and carotenoids from marine phytoplankton: A new HPLC method using a reversed phase C8 column and pyridine-containing mobile phases. Mar. Ecol. Prog. Ser. 2000, 195, 29–45. [Google Scholar] [CrossRef] [Green Version]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Somogyi, M. Notes on sugar determination. J. Biol. Chem. 1952, 195, 19–23. [Google Scholar] [CrossRef]
- Nelson, N. A photometric adaptation of the Somogyi method for the determination of glucose. J. Biol. Chem. 1944, 153, 375–380. [Google Scholar] [CrossRef]
- Blunden, C.A.; Wilson, M.F. A specific method for the determination of soluble sugars in plant extracts using enzymatic analysis and its application to the sugar content of developing pear fruit buds. Anal. Biochem. 1985, 151, 403–408. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing (R Version 4.0.2, Taking Off Again); R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 22 June 2020).
- Hojsgaard, S.; Halekoh, U. doBy: Groupwise Statistics, LSmeans, Linear Contrasts, Utilities (R Package Version 4.6.8). 2020. Available online: https://CRAN.R-project.org/package=doBy (accessed on 15 February 2020).
- Da Silva, L.J. Laercio-Package: Duncan Test, Tukey Test and Scott-Knott Test. 2010. Available online: https://CRAN.R-project.org/package=laercio (accessed on 20 February 2015).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Treatment | Brz | BL | EBL | HBL | norBL |
|---|---|---|---|---|---|
| Control | 0 | 1.79 ± 0.31 d | 0.17 ± 0.04 d | 0.63 ± 0.15 c, d | 0.12 ± 0.04 c, d |
| [1.52–2.06] | [0.13–0.21] | [0.5–0.76] | [0.09–0.15] | ||
| 0.1 µM Brz | 911.04 ± 81.21 c | 1.19 ± 0.38 d | 0.12 ± 0.03 d | 0.49 ± 0.08 d, e | 0.09 ± 0.02 c, d |
| [839.86–982.22] | [0.85–1.52] | [0.1–0.14] | [0.42–0.56] | [0.07–0.11] | |
| 1 µM Brz | 6987.57 ± 263.03 b | 0.44 ± 0.09 d | 0.06 ± 0.02 d | 0.13 ± 0.04 e | 0.05 ± 0.01 c, d |
| [6757.01−7218.13] | [0.36–0.51] | [0.05–0.08] | [0.1–0.17] | [0.04–0.06] | |
| 10 µM Brz | 9381.43 ± 1007.15 a | 0.13 ± 0.04 d | 0.02 ± 0.02 d | 0.06 ± 0.02 e | 0.02 ± 0.01 d |
| [8498.62–10264.23] | [0.1–0.17] | [0–0.04] | [0.05–0.07] | [0.01–0.02] | |
| 0.001 µM BL | 0 | 2 ± 0.23 d | 0.21 ± 0.06 d | 0.68 ± 0.14 c, d | 0.16 ± 0.04 c |
| [1.8–2.2] | [0.16–0.26] | [0.56–0.81] | [0.12–0.2] | ||
| 0.01 µM BL | 0 | 94.27 ± 19.58 d | 1.23 ± 0.17 d | 1.17 ± 0.38 b | 0.29 ± 0.06 b |
| [77.11–111.43] | [1.08–1.38] | [0.84–1.5] | [0.24–0.35] | ||
| 0.1 µM BL | 0 | 447.18 ± 40.86 b | 16.08 ± 3.2 a | 1.72 ± 0.3 a | 0.49 ± 0.11 a |
| [411.37–482.99] | [13.27–18.89] | [1.45–1.98] | [0.4–0.59] | ||
| 1 µM BL | 0 | 682.61 ± 82.16 a | 14.36 ± 1.73 a, b | 1.03 ± 0.18 b, c | 0.37 ± 0.08 b |
| [610.6–754.63] | [12.84–15.88] | [0.88–1.19] | [0.3–0.45] | ||
| 0.1 µM BL + 0.1 µM Brz | 719.6 ± 104.76 c | 433.03 ± 52.71 b | 13.37 ± 2.68 a, b | 1.21 ± 0.36 b | 0.34 ± 0.06 b |
| [627.78–811.43] | [386.83–479.24] | [11.02–15.72] | [0.9–1.53] | [0.28–0.39] | |
| 0.1 µM BL + 1 µM Brz | 6303.29 ± 1051.26 b | 362.98 ± 100.02 b | 11.68 ± 1.93 b | 0.37 ± 0.13 d, e | 0.08 ± 0.01 c, d |
| [5381.82–7224.76] | [275.31–450.65] | [9.98–13.37] | [0.26–0.48] | [0.07–0.08] | |
| 0.1 µM BL + 10 µM Brz | 10677.36 ± 2183.66 a | 259.68 ± 42 c | 6.51 ± 1.56 c | 0.1 ± 0.02 e | 0.03 ± 0.01 d |
| [8763.29–12591.42] | [222.87–296.5] | [5.14–7.87] | [0.08–0.12] | [0.02–0.03] | |
| CT | CS | ECS | TY | 6dTY | |
| Control | 1.96 ± 0.61 b, c, d | 2.77 ± 0.29 c, d | 0.03 ± 0.01 d, e | 0.18 ± 0.05 d, e, f | 0.03 ± 0.01 b, c, d |
| [1.42–2.49] | [2.52–3.03] | [0.02–0.03] | [0.14–0.22] | [0.02–0.04] | |
| 0.1 µM Brz | 1.6 ± 0.15 c, d, e | 2.2 ± 0.27 d | 0.02 ± 0 e, f | 0.14 ± 0.03 e, f, g | 0.02 ± 0.01 d |
| [1.46–1.73] | [1.96–2.43] | [0.01–0.02] | [0.12–0.16] | [0.01–0.03] | |
| 1 µM Brz | 0.94 ± 0.16 d, e, f | 1.12 ± 0.11 e, f | 0.01 ± 0 f | 0.06 ± 0.01 f, g | 0 |
| [0.8–1.08] | [1.02–1.22] | [0.01–0.01] | [0.05–0.07] | ||
| 10 µM Brz | 0.52 ± 0.13 f | 0.73 ± 0.16 f | 0.01 ± 0.01 f | 0.02 ± 0 g | 0 |
| [0.4–0.63] | [0.59–0.87] | [0–0.01] | [0.02–0.03] | ||
| 0.001 µM BL | 2.08 ± 0.15 b, c | 2.95 ± 0.17 b, c | 0.03 ± 0.01 c, d | 0.31 ± 0.05 c, d | 0.04 ± 0.01 b |
| [1.94–2.21] | [2.8–3.1] | [0.03–0.04] | [0.27–0.35] | [0.04–0.04] | |
| 0.01 µM BL | 2.76 ± 0.52 a, b | 3.53 ± 0.31 a, b | 0.04 ± 0.01 b, c | 0.49 ± 0.1 a, b | 0.05 ± 0.01 b |
| [2.31–3.22] | [3.26–3.8] | [0.04–0.05] | [0.4–0.58] | [0.04–0.05] | |
| 0.1 µM BL | 3.47 ± 0.56 a | 4.02 ± 0.26 a | 0.05 ± 0.01 a | 0.57 ± 0.12 a | 0.06 ± 0.01 a |
| [2.98–3.96] | [3.79–4.24] | [0.05–0.06] | [0.47–0.67] | [0.05–0.07] | |
| 1 µM BL | 2.03 ± 0.8 b, c | 2.75 ± 0.13 c, d | 0.03 ± 0 d, e | 0.42 ± 0.08 b, c | 0.04 ± 0.01 b |
| [1.33–2.74] | [2.63–2.87] | [0.03–0.03] | [0.35–0.48] | [0.03–0.05] | |
| 0.1 µM BL + 0.1 µM Brz | 3.01 ± 1.02 a, b | 3.54 ± 0.44 a | 0.05 ± 0 a, b | 0.25 ± 0.14 d, e | 0.04 ± 0.01 b, c |
| [2.12–3.9] | [3.15–3.92] | [0.04–0.05] | [0.14–0.37] | [0.03–0.04] | |
| 0.1 µM BL + 1 µM Brz | 1.25 ± 0.13 c, d, e, f | 1.6 ± 0.44 e | 0.01 ± 0 f | 0.03 ± 0.01 g | 0.02 ± 0.01 c, d |
| [1.14–1.37] | [1.21–1.99] | [0.01–0.02] | [0.02–0.04] | [0.02–0.03] | |
| 0.1 µM BL + 10 µM Brz | 0.7 ± 0.11 e, f | 0.87 ± 0.13 f | 0.01 ± 0 f | 0 | 0 |
| [0.61–0.79] | [0.76–0.98] | [0.01–0.02] |
| Treatment | Chlorophyll a | Chlorophyll b | α-Carotene | β-Carotene | Neoxanthin |
|---|---|---|---|---|---|
| Control | 162.16 ± 2.69 e, f | 42.19 ± 2.2 d, e | 1.29 ± 0.01 e | 1.78 ± 0.07 c | 0.92 ± 0.04 e |
| [159.8–164.51] | [40.26–44.13] | [1.28–1.3] | [1.72–1.84] | [0.88–0.95] | |
| 0.1 µM Brz | 157.7 ± 1.53 f | 40.63 ± 1.68 e | 1.17 ± 0.01 f | 1.52 ± 0.07 d | 0.85 ± 0.01 e |
| [156.36–159.04] | [39.15–42.1] | [1.15–1.18] | [1.47–1.58] | [0.84–0.86] | |
| 1 µM Brz | 120.71 ± 1.52 h | 29.77 ± 1.01 g | 0.91 ± 0.05 g | 1.2 ± 0.05 e | 0.52 ± 0.04 g |
| [119.38–122.04] | [28.88–30.65] | [0.87–0.95] | [1.16–1.25] | [0.48–0.55] | |
| 10 µM Brz | 87.77 ± 2.41 i | 20.35 ± 0.89 i | 0.61 ± 0.02 h | 0.78 ± 0.1 g | 0.35 ± 0.02 h |
| [85.66–89.89] | [19.57–21.13] | [0.59–0.63] | [0.69–0.86] | [0.33–0.36] | |
| 0.001 µM BL | 163.83 ± 2.73 e | 44.64 ± 3.02 d | 1.49 ± 0.03 d | 1.83 ± 0.03 c | 1.14 ± 0.08 d |
| [161.44–166.22] | [41.99–47.28] | [1.46–1.52] | [1.8–1.86] | [1.07–1.2] | |
| 0.01 µM BL | 192.25 ± 2.52 c | 50.91 ± 2.27 c | 1.73 ± 0.06 c | 2.24 ± 0.08 b | 1.31 ± 0.02 b |
| [190.04–194.45] | [48.92–52.9] | [1.68–1.78] | [2.17–2.31] | [1.29–1.33] | |
| 0.1 µM BL | 222.8 ± 1.73 a | 61.64 ± 1.23 a | 2.4 ± 0.05 a | 2.79 ± 0.12 a | 1.59 ± 0.04 a |
| [221.28–224.32] | [60.57–62.72] | [2.36–2.44] | [2.69–2.9] | [1.56–1.62] | |
| 1 µM BL | 172.23 ± 2.69 d | 49.07 ± 1.33 c | 1.74 ± 0.07 c | 2.29 ± 0.06 b | 1.22 ± 0.04 c |
| [169.87–174.59] | [47.9–50.23] | [1.68–1.8] | [2.24–2.34] | [1.18–1.25] | |
| 0.1 µM BL + 0.1 µM Brz | 202.95 ± 2.26 b | 55.45 ± 1.14 b | 2.12 ± 0.04 b | 2.68 ± 0.07 a | 1.27 ± 0.03 b, c |
| [200.97–204.93] | [54.46–56.45] | [2.09–2.15] | [2.62–2.74] | [1.25–1.3] | |
| 0.1 µM BL + 1 µM Brz | 152.32 ± 1.44 g | 35.01 ± 1.7 f | 1.38 ± 0.04 e | 1.7 ± 0.01 c | 0.73 ± 0.01 f |
| [151.06–153.58] | [33.52–36.5] | [1.34–1.42] | [1.69–1.71] | [0.72–0.74] | |
| 0.1 µM BL + 10 µM Brz | 122.16 ± 4.24 h | 25.23 ± 1.05 h | 0.81 ± 0.09 g | 1.04 ± 0.04 f | 0.45 ± 0.01 g |
| [118.44–125.88] | [24.31–26.15] | [0.72–0.89] | [1.01–1.08] | [0.44–0.46] | |
| Violaxanthin | Astaxanthin | Zeaxanthin | Cryptoxanthin | Lutein | |
| Control | 0.7 ± 0.01 f | 0.35 ± 0.02 d, e | 3.21 ± 0.06 d | 4.24 ± 0.08 e | 0.47 ± 0.01 d |
| [0.69–0.71] | [0.33–0.37] | [3.16–3.26] | [4.17–4.31] | [0.47–0.48] | |
| 0.1 µM Brz | 0.6 ± 0.01 h | 0.32 ± 0.01 e, f | 2.73 ± 0.1 e | 3.96 ± 0.15 f | 0.42 ± 0.02 e |
| [0.6–0.61] | [0.31–0.33] | [2.64–2.82] | [3.83–4.09] | [0.4–0.44] | |
| 1 µM Brz | 0.45 ± 0.01 i | 0.18 ± 0.01 g | 2.09 ± 0.14 f | 2.58 ± 0.1 g | 0.33 ± 0.02 f |
| [0.45–0.46] | [0.17–0.19] | [1.98–2.21] | [2.49–2.66] | [0.31–0.35] | |
| 10 µM Brz | 0.25 ± 0.01 k | 0.08 ± 0.01 h | 0.94 ± 0.07 h | 2.15 ± 0.07 h | 0.25 ± 0.02 h |
| [0.25–0.26] | [0.07–0.09] | [0.88–0.99] | [2.09–2.21] | [0.22–0.27] | |
| 0.001 µM BL | 0.73 ± 0.01 e | 0.38 ± 0.01 d | 3.27 ± 0.06 d | 4.42 ± 0.13 e | 0.51 ± 0.01 c |
| [0.72–0.74] | [0.38–0.39] | [3.22–3.32] | [4.31–4.54] | [0.5–0.53] | |
| 0.01 µM BL | 0.84 ± 0.01 c | 0.51 ± 0.01 b | 3.77 ± 0.08 c | 5.21 ± 0.13 c | 0.61 ± 0.01 b |
| [0.83–0.85] | [0.5–0.52] | [3.69–3.84] | [5.09–5.32] | [0.6–0.62] | |
| 0.1 µM BL | 0.96 ± 0.01 a | 0.58 ± 0.01 a | 4.61 ± 0.09 a | 6.25 ± 0.08 a | 0.72 ± 0.01 a |
| [0.95–0.97] | [0.58–0.59] | [4.53–4.69] | [6.19–6.32] | [0.71–0.73] | |
| 1 µM BL | 0.78 ± 0.01 d | 0.44 ± 0.01 c | 3.65 ± 0.06 c | 4.93 ± 0.16 d | 0.6 ± 0.01 b |
| [0.76–0.79] | [0.43–0.45] | [3.59–3.7] | [4.79–5.07] | [0.59–0.61] | |
| 0.1 µM BL + 0.1 µM Brz | 0.91 ± 0.01 b | 0.55 ± 0.01 a, b | 4.23 ± 0.08 b | 5.92 ± 0.05 b | 0.62 ± 0.01 b |
| [0.91–0.92] | [0.54–0.55] | [4.16–4.29] | [5.87–5.97] | [0.6–0.63] | |
| 0.1 µM BL + 1 µM Brz | 0.65 ± 0.01 g | 0.28 ± 0.03 f | 2.72 ± 0.11 e | 3.73 ± 0.12 f | 0.47 ± 0.01 d |
| [0.64–0.66] | [0.26–0.31] | [2.62–2.82] | [3.62–3.83] | [0.46–0.48] | |
| 0.1 µM BL + 10 µM Brz | 0.33 ± 0.01 j | 0.16 ± 0.04 g | 1.3 ± 0.08 g | 2.68 ± 0.16 g | 0.3 ± 0.01 g |
| [0.32–0.34] | [0.13–0.19] | [1.23–1.37] | [2.54–2.82] | [0.28–0.31] |
| Compound | Precursor m/z | Product m/z | CE | Retention Time (min) |
|---|---|---|---|---|
| Brz | 327.8 | 70.05 | −25 | 3.976 |
| BL | 610.0 | 176.05 | −43 | 11.201 |
| EBL | 610.0 | 190.2 | −42 | 10.720 |
| HBL | 624.0 | 190.1 | −43 | 13.118 |
| norBL | 596.0 | 190.0 | −44 | 9.350 |
| CT | 433.0 | 415.2 | −21 | 1.796 |
| CS | 594.1 | 175.95 | −36 | 13.254 |
| ECS | 594.1 | 175.95 | −38 | 13.552 |
| 6dCS | 580.1 | 562.2 | −40 | 23.778 |
| TY | 578.1 | 560.15 | −37 | 18765 |
| 6dTY | 564.2 | 176.15 | −55 | 18.957 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chmur, M.; Bajguz, A. Brassinolide Enhances the Level of Brassinosteroids, Protein, Pigments, and Monosaccharides in Wolffia arrhiza Treated with Brassinazole. Plants 2021, 10, 1311. https://doi.org/10.3390/plants10071311
Chmur M, Bajguz A. Brassinolide Enhances the Level of Brassinosteroids, Protein, Pigments, and Monosaccharides in Wolffia arrhiza Treated with Brassinazole. Plants. 2021; 10(7):1311. https://doi.org/10.3390/plants10071311
Chicago/Turabian StyleChmur, Magdalena, and Andrzej Bajguz. 2021. "Brassinolide Enhances the Level of Brassinosteroids, Protein, Pigments, and Monosaccharides in Wolffia arrhiza Treated with Brassinazole" Plants 10, no. 7: 1311. https://doi.org/10.3390/plants10071311