
Light Harvesting in Fluctuating Environments: Evolution and Function of Antenna Proteins across Photosynthetic Lineage


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
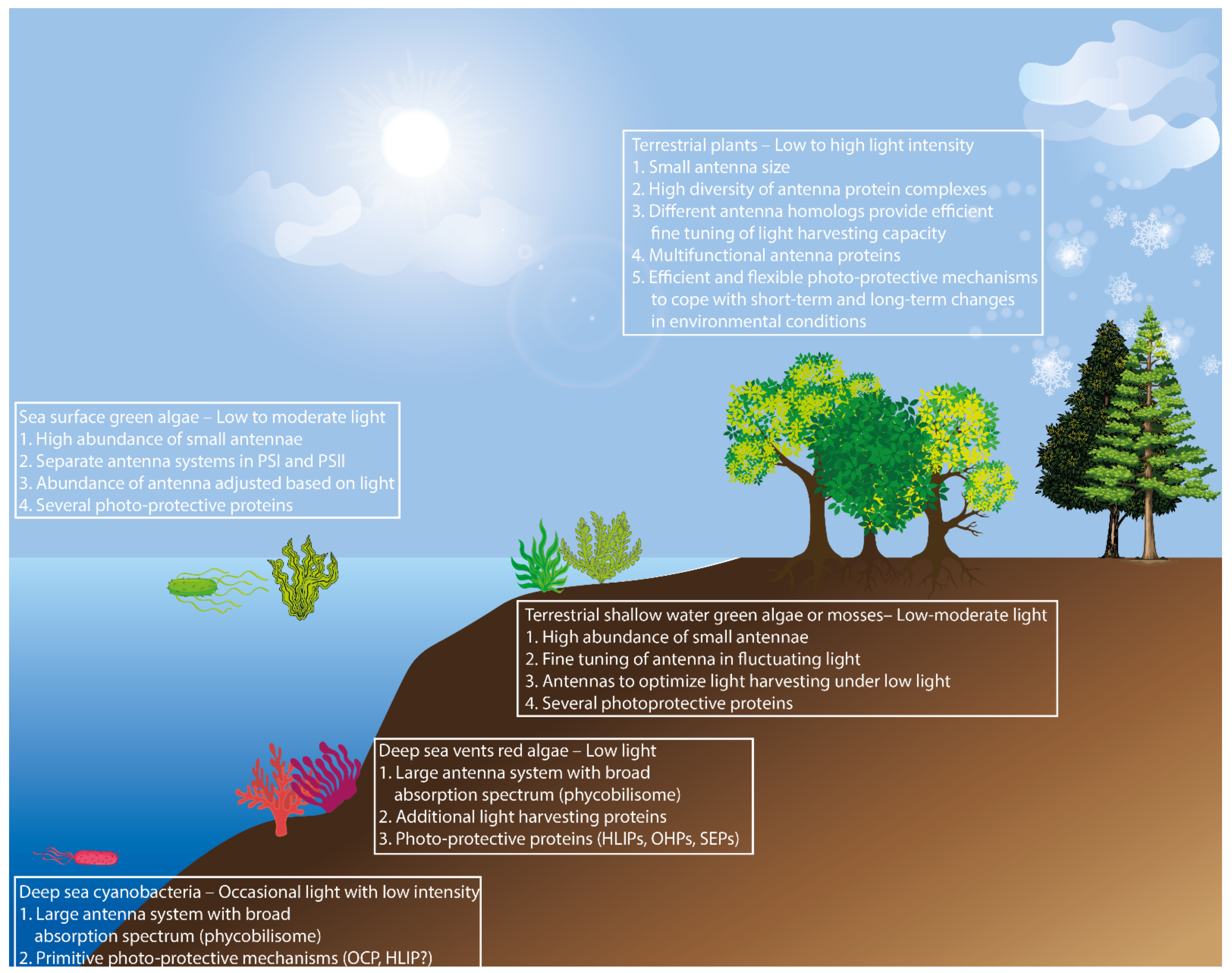
:1. Introduction
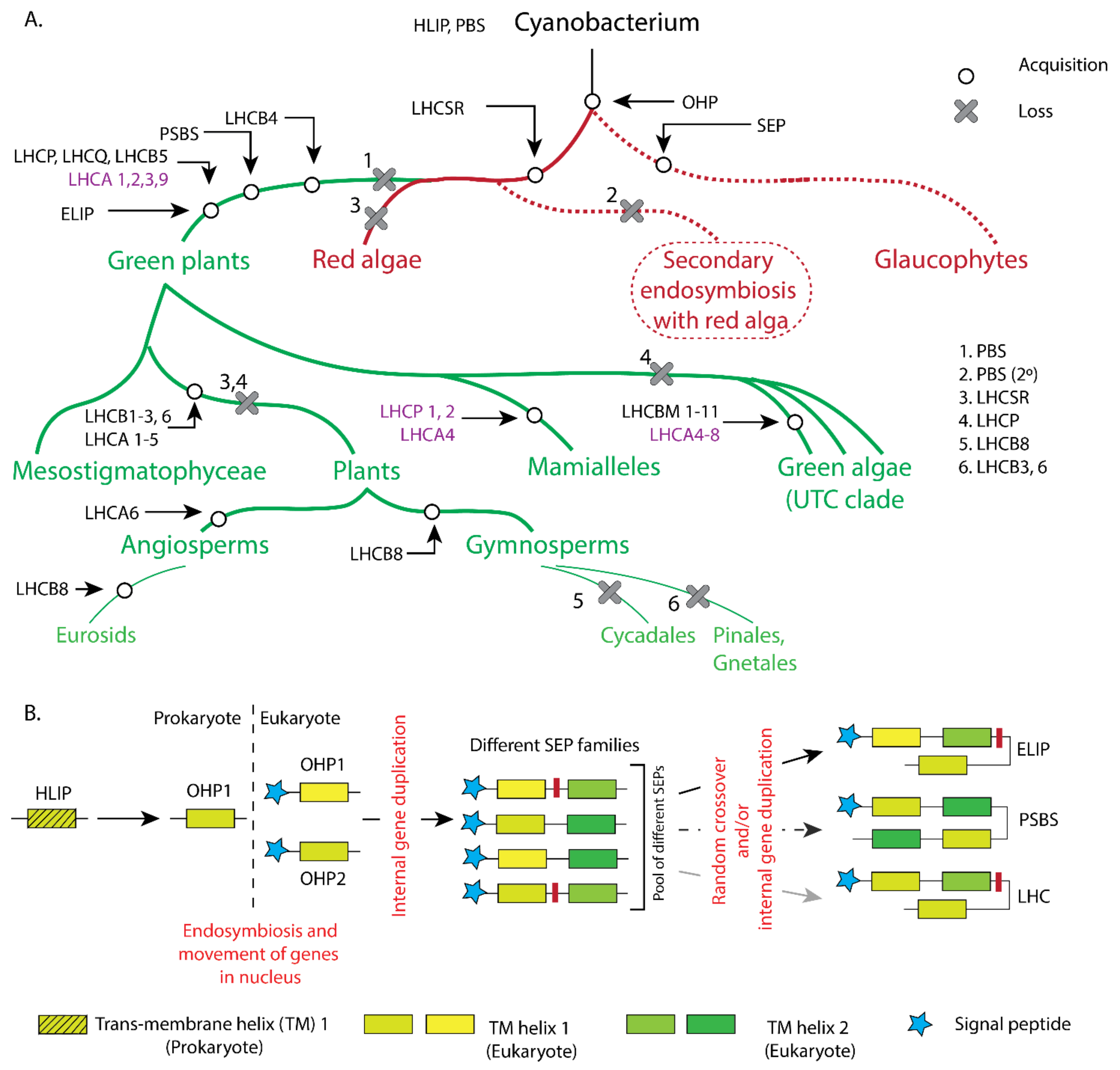
2. Evolutionary Aspects of Light-Harvesting Antennae
2.1. Light-Harvesting Antennae in Cyanobacteria
2.2. Light-Harvesting Antenna in Red Lineages
2.3. Light-Harvesting Antennae in Green Algae
2.4. Light Harvesting in Terrestrial Green Lineage
3. The Functional Properties of Light-Harvesting Antennae in Relation to Coping Mechanisms under Fluctuating Environmental Conditions
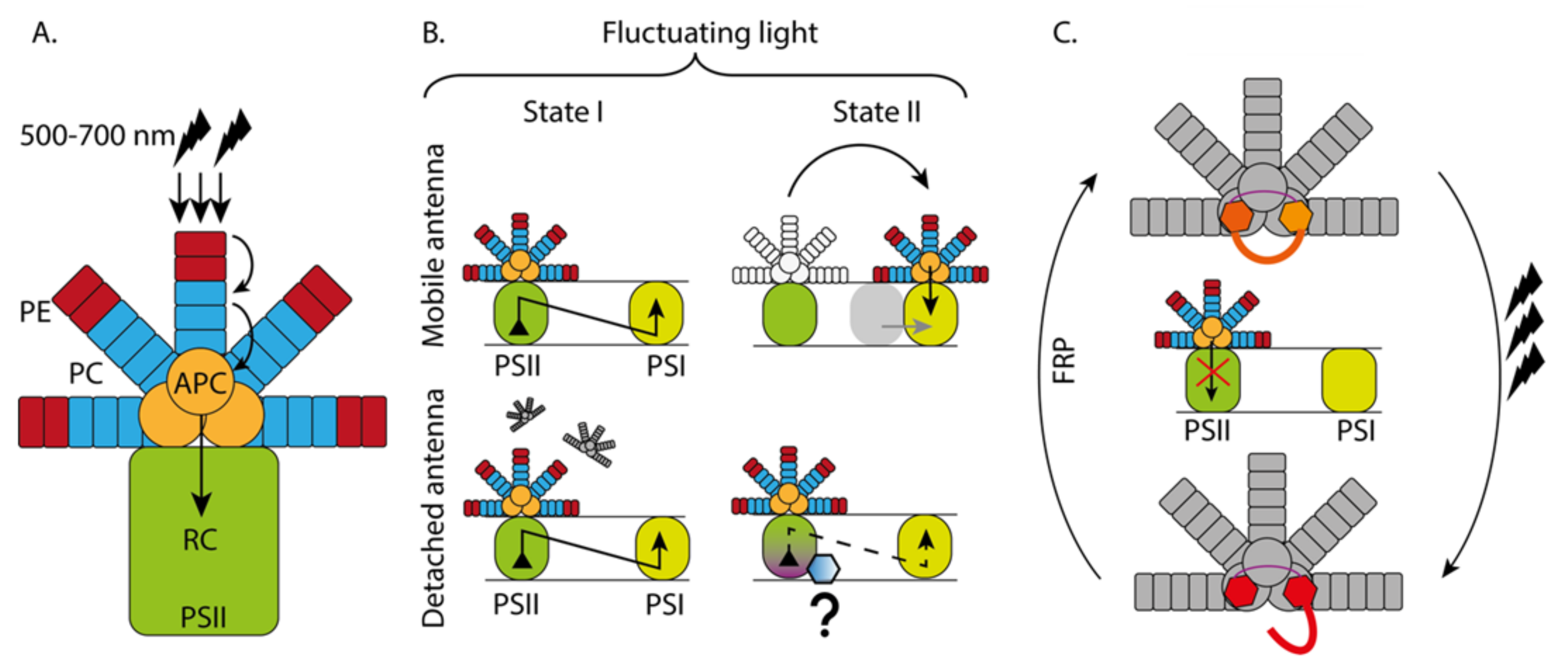
3.1. Phycobilisome Antenna System (PBS)
3.1.1. Light Harvesting Capacity of PBS
3.1.2. PBS Functioning under Different Light Conditions
3.2. Orange Carotenoid Proteins (OCPs) Facilitate Short-Term Acclimation to High Light
3.3. HLIP and Eukaryotic LIL Proteins
3.3.1. The HLIP Family Is Associated with High Light and Oxidative Stress Responses
3.3.2. OHP Family
3.3.3. SEP Family
3.3.4. ELIP Family
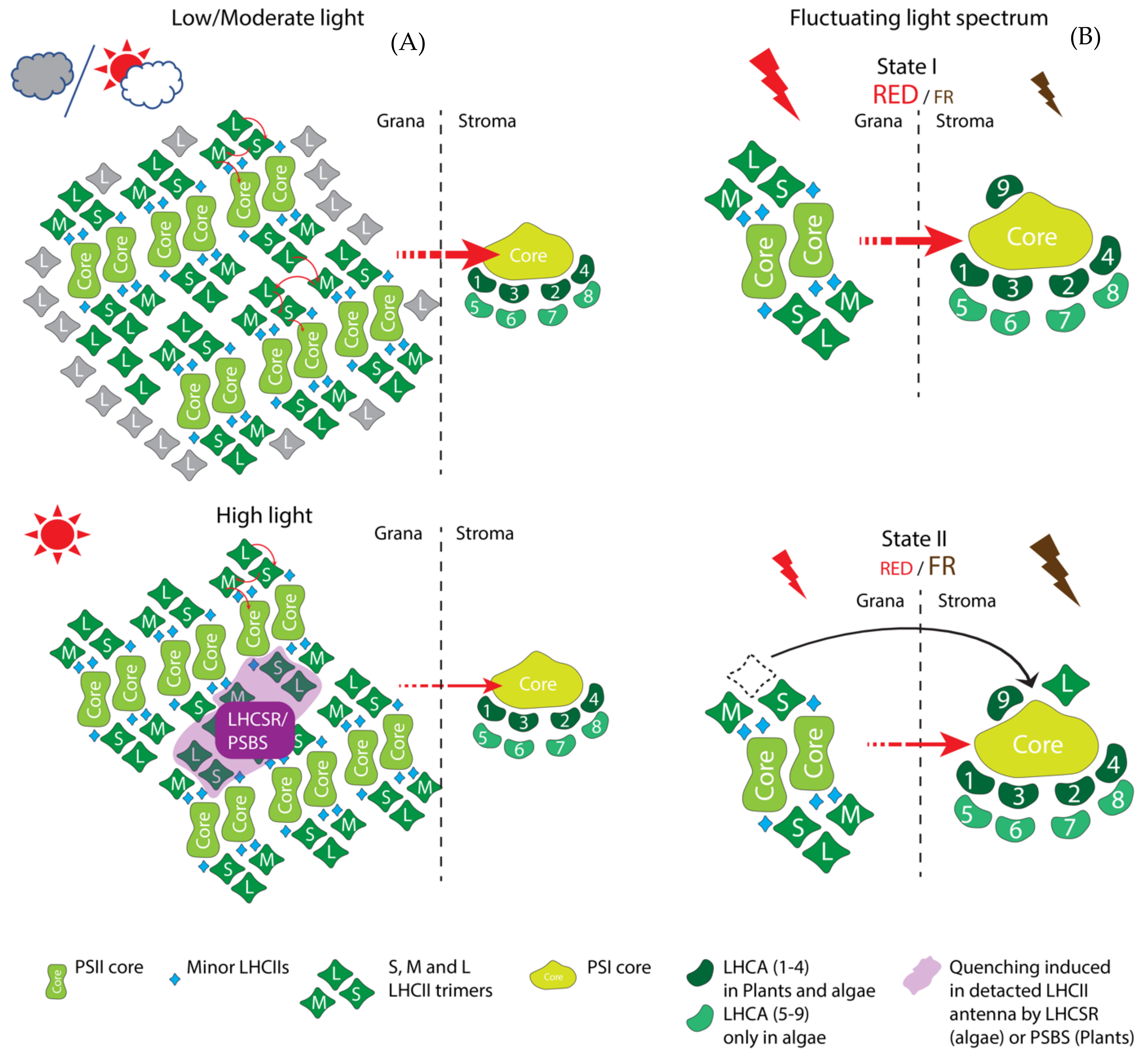
3.4. LhcSR Family Proteins Act in Non-Photochemical Quenching
3.5. Lhc Family the ‘True Lhcs’ Are the Most Complex and Diverse Group of Light-Harvesting Antennae
3.5.1. Minor PSII Antenna—LhcB4 to LhcB9
3.5.2. Major PSII Antenna—LhcP and LhcQ
3.5.3. Major PSII Antennae—LhcBM1 to 11
3.5.4. Major PSII Antennae—LhcB1 to 3
3.5.5. PSI Antenna—LhcAs
3.6. Photosystem II Subunit S (PsbS)
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nevins, D.J. Sugars: Their origin in photosynthesis and subsequent biological interconversions. Am. J. Clin. Nutr. 1995, 61, 915S–921S. [Google Scholar] [CrossRef] [Green Version]
- Jansson, S. A guide to the Lhc genes and their relatives in Arabidopsis. Trends Plant Sci. 1999, 4, 236–240. [Google Scholar] [CrossRef]
- Boekema, E.J.; Hankamer, B.; Bald, D.; Kruip, J.; Nield, J.; Boonstra, A.F.; Barber, J.; Rögner, M. Supramolecular structure of the photosystem II complex from green plants and cyanobacteria. Proc. Natl. Acad. Sci. USA 1995, 92, 175–179. [Google Scholar] [CrossRef] [Green Version]
- Ben-Shem, A.; Frolow, F.; Nelson, N. Crystal structure of plant photosystem I. Nature 2003, 426, 630–635. [Google Scholar] [CrossRef]
- Bedbrook, J.R.; Smith, S.M.; Ellis, R.J. Molecular cloning and sequencing of cDNA encoding the precursor to the small subunit of chloroplast ribulose-1, 5-bisphosphate carboxylase. Nature 1980, 287, 692–697. [Google Scholar] [CrossRef]
- Alboresi, A.; Caffarri, S.; Nogue, F.; Bassi, R.; Morosinotto, T. In silico and biochemical analysis of Physcomitrella patens photosynthetic antenna: Identification of subunits which evolved upon land adaptation. PLoS ONE 2008, 3, e2033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaul, S.; Koo, H.L.; Jenkins, J.; Rizzo, M.; Rooney, T.; Tallon, L.J.; Feldblyum, T.; Nierman, W.; Benito, M.I.; Lin, X.; et al. Analysis of the genome sequence of the flowering plant Arabidopsis thaliana. Nature 2000, 408, 796–815. [Google Scholar]
- Jansson, S.; Pichersky, E.; Bassi, R.; Green, B.R.; Ikeuchi, M.; Melis, A.; Simpson, D.J.; Spangfort, M.; Staehelin, L.A.; Thornber, J.P. A nomenclature for the genes encoding the chlorophylla/b-binding proteins of higher plants. Plant Mol. Biol. Rep. 1992, 10, 242–253. [Google Scholar] [CrossRef]
- Li, L.; Wang, S.; Wang, H.; Sahu, S.K.; Marin, B.; Li, H.; Xu, Y.; Liang, H.; Li, Z.; Cheng, S.; et al. The genome of Prasinoderma coloniale unveils the existence of a third phylum within green plants. Nat. Ecol. Evol. 2020, 4, 1220–1231. [Google Scholar] [CrossRef] [PubMed]
- Bruce, D.; Brimble, S.; Bryant, D.A. State transitions in a phycobilisome-less mutant of the cyanobacterium Synechococcus sp. PCC 7002. Biochim. Biophys. Acta BBA Bioenerg. 1989, 974, 66–73. [Google Scholar] [CrossRef]
- MacColl, R. Cyanobacterial phycobilisomes. J. Struct. Biol. 1998, 124, 311–334. [Google Scholar] [CrossRef] [PubMed]
- Neilson, J.A.D.; Durnford, D.G. Structural and functional diversification of the light-harvesting complexes in photosynthetic eukaryotes. Photosynth. Res. 2010, 106, 57–71. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.-N.; Chen, X.-L.; Zhang, Y.-Z.; Zhou, B.-C. Characterization, structure and function of linker polypeptides in phycobilisomes of cyanobacteria and red algae: An overview. Biochim. Biophys. Acta BBA Bioenerg. 2005, 1708, 133–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adir, N.; Bar-Zvi, S.; Harris, D. The amazing phycobilisome. Biochim. Biophys. Acta BBA Bioenerg. 2020, 1861, 148047. [Google Scholar] [CrossRef]
- Wiltbank, L.B.; Kehoe, D.M. Diverse light responses of cyanobacteria mediated by phytochrome superfamily photoreceptors. Nat. Rev. Microbiol. 2019, 17, 37–50. [Google Scholar] [CrossRef] [PubMed]
- Veith, T.; Büchel, C. The monomeric photosystem I-complex of the diatom Phaeodactylum tricornutum binds specific fucoxanthin chlorophyll proteins (FCPs) as light-harvesting complexes. Biochim. Biophys. Acta BBA Bioenerg. 2007, 1767, 1428–1435. [Google Scholar] [CrossRef] [Green Version]
- Joshi-Deo, J.; Schmidt, M.; Gruber, A.; Weisheit, W.; Mittag, M.; Kroth, P.G.; Büchel, C. Characterization of a trimeric light-harvesting complex in the diatom Phaeodactylum tricornutum built of FcpA and FcpE proteins. J. Exp. Bot. 2010, 61, 3079–3087. [Google Scholar] [CrossRef]
- Kansy, M.; Volke, D.; Sturm, L.; Wilhelm, C.; Hoffmann, R.; Goss, R. Pre-purification of diatom pigment protein complexes provides insight into the heterogeneity of FCP complexes. BMC Plant Biol. 2020, 20, 1–16. [Google Scholar] [CrossRef]
- Nagao, R.; Yokono, M.; Ueno, Y.; Suzuki, T.; Kumazawa, M.; Kato, K.H.; Tsuboshita, N.; Dohmae, N.; Ifuku, K.; Shen, J.R.; et al. Enhancement of excitation-energy quenching in fucoxanthin chlorophyll a/c-binding proteins isolated from a diatom Phaeodactylum tricornutum upon excess-light illumination. Biochim. Biophys. Acta BBA Bioenerg. 2021, 1862, 148350. [Google Scholar] [CrossRef]
- Elnour, H.M.A.M.; Dietzel, L.; Ramanan, C.; Büchel, C.; van Grondelle, R.; Krüger, T.P.J. Energy dissipation mechanisms in the FCPb light-harvesting complex of the diatom Cyclotella meneghiniana. Biochim. Biophys. Acta BBA Bioenerg. 2018, 1859, 1151–1160. [Google Scholar] [CrossRef] [PubMed]
- Agostini, A.; Büchel, C.; di Valentin, M.; Carbonera, D. A distinctive pathway for triplet-triplet energy transfer photoprotection in fucoxanthin chlorophyll-binding proteins from Cyclotella meneghiniana. Biochim. Biophys. Acta BBA Bioenerg. 2021, 1862, 148310. [Google Scholar] [CrossRef]
- Schmid, V.H.R. Light-harvesting complexes of vascular plants. Cell. Mol. Life Sci. 2008, 65, 3619–3639. [Google Scholar] [CrossRef]
- Ballottari, M.; Girardon, J.; DallOsto, L.; Bassi, R. Evolution and functional properties of photosystem II light harvesting complexes in eukaryotes. Biochim. Biophys. Acta BBA Bioenerg. 2012, 1817, 143–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kunugi, M.; Satoh, S.; Ihara, K.; Shibata, K.; Yamagishi, Y.; Kogame, K.; Obokata, J.; Takabayashi, A.; Tanaka, A. Evolution of green plants accompanied changes in light-harvesting systems. Plant Cell Physiol. 2016, 57, 1231–1243. [Google Scholar] [CrossRef]
- Taddei, L.; Stella, G.R.; Rogato, A.; Bailleul, B.; Fortunato, A.E.; Annunziata, R.; Sanges, R.; Thaler, M.; Lepetit, B.; Lavaud, J.; et al. Multisignal control of expression of the LhcX protein family in the marine diatom Phaeodactylum tricornutum. J. Exp. Bot. 2016, 67, 3939–3951. [Google Scholar] [CrossRef] [Green Version]
- Grebe, S.; Trotta, A.; Bajwa, A.A.; Mancini, I.; Bag, P.; Jansson, S.; Tikkanen, M.; Aro, E.M. Specific thylakoid protein phosphorylations are prerequisites for overwintering of Norway spruce (Picea abies) photosynthesis. Proc. Natl. Acad. Sci. USA 2020, 117, 17499–17509. [Google Scholar] [CrossRef] [PubMed]
- Bag, P.; Chukhutsina, V.; Zhang, Z.; Paul, S.; Ivanov, A.G.; Shutova, T.; Croce, R.; Holzwarth, A.R.; Jansson, S. Direct energy transfer from photosystem II to photosystem I confers winter sustainability in Scots Pine. Nat. Commun. 2020, 11, 6388. [Google Scholar] [CrossRef] [PubMed]
- Darwin, C. On the Origin of Species; Harvard University Press: Cambridge, MA, USA, 1859; Volume 24. [Google Scholar]
- Darwin, C. The Movements and Habits of Climbing Plants; SoC. London: London, UK, 1875. [Google Scholar]
- Ruban, V. Evolution under the sun: Optimizing light harvesting in photosynthesis. J. Exp. Bot. 2015, 66, 7–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grossman, R.; Bhaya, D.; Apt, K.E.; Kehoe, D.M. Light-harvesting complexes in oxygenic photosynthesis: Diversity, control, and evolution. Annu. Rev. Genet. 1995, 29, 231–288. [Google Scholar] [CrossRef]
- Gupta, R.S. Origin and spread of photosynthesis based upon conserved sequence features in key bacteriochlorophyll biosynthesis proteins. Mol. Biol. Evol. 2012, 29, 3397–3412. [Google Scholar] [CrossRef]
- Dalrymple, G.B. The age of the Earth in the twentieth century: A problem (mostly) solved. Geol. Soc. Lond. Spec. Publ. 2001, 190, 205–221. [Google Scholar] [CrossRef]
- Xiong, J.; Bauer, C.E. Complex evolution of photosynthesis. Annu. Rev. Plant Biol. 2002, 53, 503–521. [Google Scholar] [CrossRef]
- Blankenship, R.E. Early evolution of photosynthesis. Plant Physiol. 2010, 154, 434–438. [Google Scholar] [CrossRef] [Green Version]
- Apt, K.E.; Collier, J.L.; Grossman, A.R. Evolution of the phycobiliproteins. J. Mol. Biol. 1995, 248, 79–96. [Google Scholar] [CrossRef] [PubMed]
- Schirmer, T.; Bode, W.; Huber, R.; Sidler, W.; Zuber, H. X-ray crystallographic structure of the light-harvesting biliprotein C-phycocyanin from the thermophilic cyanobacterium Mastigocladus laminosus and its resemblance to globin structures. J. Mol. Biol. 1985, 184, 257–277. [Google Scholar] [CrossRef]
- Dolganov, N.A.; Bhaya, D.; Grossman, A.R. Cyanobacterial protein with similarity to the chlorophyll a/b binding proteins of higher plants: Evolution and regulation. Proc. Natl. Acad. Sci. USA 1995, 92, 636–640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komenda, J.; Sobotka, R. Cyanobacterial high-light-inducible proteins—protectors of chlorophyll–protein synthesis and assembly. Biochim. Biophys. Acta BBA Bioenerg. 2016, 1857, 288–295. [Google Scholar] [CrossRef] [PubMed]
- Heddad, M.; Adamska, I. The evolution of light stress proteins in photosynthetic organisms. Comp. Funct. Genom. 2002, 3, 504–510. [Google Scholar] [CrossRef] [Green Version]
- Kerfeld, A.; Melnicki, M.R.; Sutter, M.; Dominguez-Martin, M.A. Structure, function and evolution of the cyanobacterial orange carotenoid protein and its homologs. New Phytol. 2017, 215, 937–951. [Google Scholar] [CrossRef] [Green Version]
- Savard, F.; Richard, C.; Guertin, M. The Chlamydomonas reinhardtii LI818 gene represents a distant relative of the cabI/II genes that is regulated during the cell cycle and in response to illumination. Plant Mol. Biol. 1996, 32, 461–473. [Google Scholar] [CrossRef]
- Koziol, G.; Borza, T.; Ishida, K.-I.; Keeling, P.; Lee, R.W.; Durnford, D.G. Tracing the evolution of the light-harvesting antennae in chlorophyll a/b-containing organisms. Plant Physiol. 2007, 143, 1802–1816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engelken, J.; Brinkmann, H.; Adamska, I. Taxonomic distribution and origins of the extended Lhc (light-harvesting complex) antenna protein superfamily. BMC Evol. Biol. 2010, 10, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Richard, C.; Ouellet, H.; Guertin, M. Characterization of the LI818 polypeptide from the green unicellular alga Chlamydomonas reinhardtii. Plant Mol. Biol. 2000, 42, 303–316. [Google Scholar] [CrossRef] [PubMed]
- Rockwell, N.C.; Martin, S.S.; Li, F.; Mathews, S.; Lagarias, J.C. The phycocyanobilin chromophore of streptophyte algal phytochromes is synthesized by HY2. New Phytol. 2017, 214, 1145–1157. [Google Scholar] [CrossRef] [Green Version]
- Green, B.R. What Happened to the Phycobilisome? Biomolecules 2019, 9, 748. [Google Scholar] [CrossRef] [Green Version]
- Rogers, M.B.; Gilson, P.R.; Su, V.; McFadden, G.I.; Keeling, P.J. The complete chloroplast genome of the chlorarachniophyte Bigelowiella natans: Evidence for independent origins of chlorarachniophyte and euglenid secondary endosymbionts. Mol. Biol. Evol. 2007, 24, 54–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Six, C.; Worden, A.Z.; Rodríguez, F.; Moreau, H.; Partensky, F. New insights into the nature and phylogeny of prasinophyte antenna proteins: Ostreococcus tauri, a case study. Mol. Biol. Evol. 2005, 22, 2217–2230. [Google Scholar] [CrossRef] [Green Version]
- Li, X.-P.; BjoÈrkman, O.; Shih, C.; Grossman, A.R.; Rosenquist, M.; Jansson, S.; Niyogi, K.K. A pigment-binding protein essential for regulation of photosynthetic light harvesting. Nature 2000, 403, 391–395. [Google Scholar] [CrossRef]
- Lee, R.E. (Ed.) Chlorophyta. In Phycology, 5th ed.; Cambridge University Press: Cambridge, MA, USA, 2018; pp. 133–230. [Google Scholar]
- Sánchez-Baracaldo, P.; Raven, J.A.; Pisani, D.; Knoll, A.H. Early photosynthetic eukaryotes inhabited low-salinity habitats. Proc. Natl. Acad. Sci. USA 2017, 114, E7737–E7745. [Google Scholar] [CrossRef] [Green Version]
- Shively, J.M. Complex Intracellular Structures in Prokaryotes; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2006. [Google Scholar]
- Guidi, L.; Tattini, M.; Landi, M. How does chloroplast protect chlorophyll against excessive light. In Chlorophyll; IntechOpen: Rijeka, Croatia, 2017; p. 21. [Google Scholar]
- Nguyen, Y.; Bricker, W.P.; Zhang, H.; Weisz, D.A.; Gross, M.L.; Pakrasi, H.B. The proteolysis adaptor, NblA, binds to the N-terminus of β-phycocyanin: Implications for the mechanism of phycobilisome degradation. Photosynth. Res. 2017, 132, 95–106. [Google Scholar] [CrossRef] [Green Version]
- Saer, R.G.; Blankenship, R.E. Light harvesting in phototrophic bacteria: Structure and function. Biochem. J. 2017, 474, 2107–2131. [Google Scholar] [CrossRef] [PubMed]
- Nganou, C.; David, L.; Adir, N.; Mkandawire, M. Linker proteins enable ultrafast excitation energy transfer in the phycobilisome antenna system of Thermosynechococcus vulcanus. Photochem. Photobiol. Sci. 2016, 15, 31–44. [Google Scholar] [CrossRef] [PubMed]
- Calzadilla, P.I.; Kirilovsky, D. Revisiting cyanobacterial state transitions. Photochem. Photobiol. Sci. 2020, 19, 585–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mullineaux, C.W. Excitation energy transfer from phycobilisomes to photosystem I in a cyanobacterial mutant lacking photosystem II. Biochim. Biophys. Acta (BBA) Bioenerg. 1994, 1184, 71–77. [Google Scholar] [CrossRef]
- Mullineaux, C.W.; Allen, J.F. The state 2 transition in the cyanobacterium Synechococcus 6301 can be driven by respiratory electron flow into the plastoquinone pool. FEBS Lett. 1986, 205, 155–160. [Google Scholar] [CrossRef] [Green Version]
- Bruce, D.; Biggins, J.; Steiner, T.; Thewalt, M. Mechanism of the light state transition in photosynthesis. IV. Picosecond fluorescence spectroscopy of Anacystis nidulans and Porphyridium cruentum in state 1 and state 2 at 77 K. Biochim. Biophys. Acta BBA Bioenerg. 1985, 806, 237–246. [Google Scholar] [CrossRef]
- Steinbeck, J.; Ross, I.L.; Rothnagel, R.; Gäbelein, P.; Schulze, S.; Giles, N.; Ali, R.; Drysdale, R.; Sierecki, E.; Gambin, Y.; et al. Structure of a PSI-LhcI-cyt b6f supercomplex in Chlamydomonas reinhardtii promoting cyclic electron flow under anaerobic conditions. Proc. Natl. Acad. Sci. USA 2018, 115, 10517–10522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lemeille, S.; Willig, A.; Depège-Fargeix, N.; Delessert, C.; Bassi, R.; Rochaix, J.-D. Analysis of the chloroplast protein kinase Stt7 during state transitions. PLoS Biol. 2009, 7, e1000045. [Google Scholar] [CrossRef]
- Takahashi, H.; Iwai, M.; Takahashi, Y.; Minagawa, J. Identification of the mobile light-harvesting complex II polypeptides for state transitions in Chlamydomonas reinhardtii. Proc. Natl. Acad. Sci. USA 2006, 103, 477–482. [Google Scholar] [CrossRef] [Green Version]
- Leoni, C.; Pietrzykowska, M.; Kiss, A.Z.; Suorsa, M.; Ceci, L.R.; Aro, E.M.; Jansson, S. Very rapid phosphorylation kinetics suggest a unique role for L hcb2 during state transitions in Arabidopsis. Plant J. 2013, 76, 236–246. [Google Scholar] [CrossRef]
- Bergner, S.V.; Scholz, M.; Trompelt, K.; Barth, J.; Gäbelein, P.; Steinbeck, J.; Xue, H.; Clowez, S.; Fucile, G.; Goldschmidt-Clermont, M.; et al. STATE TRANSITION7-dependent phosphorylation is modulated by changing environmental conditions, and its absence triggers remodeling of photosynthetic protein complexes. Plant Physiol. 2015, 168, 615–634. [Google Scholar] [CrossRef] [Green Version]
- Kopečná, J.; Komenda, J.; Bučinská, L.; Sobotka, R. Long-term acclimation of the cyanobacterium Synechocystis sp. PCC 6803 to high light is accompanied by an enhanced production of chlorophyll that is preferentially channeled to trimeric photosystem I. Plant Physiol. 2012, 160, 2239–2250. [Google Scholar] [CrossRef] [Green Version]
- Gwizdala, M.; Wilson, A.; Kirilovsky, D. In vitro reconstitution of the cyanobacterial photoprotective mechanism mediated by the Orange Carotenoid Protein in Synechocystis PCC 6803. Plant Cell 2011, 23, 2631–2643. [Google Scholar] [CrossRef] [Green Version]
- Kerfeld, C.A.; Sawaya, M.R.; Brahmandam, V.; Cascio, D.; Ho, K.K.; Trevithick-Sutton, C.C.; Krogmann, D.W.; Yeates, T.O. The crystal structure of a cyanobacterial water-soluble carotenoid binding protein. Structure 2003, 11, 55–65. [Google Scholar] [CrossRef] [Green Version]
- Wilson, A.; Kinney, J.N.; Zwart, P.H.; Punginelli, C.; d’Haene, S.; Perreau, F.; Klein, M.G.; Kirilovsky, D.; Kerfeld, C.A. Structural determinants underlying photoprotection in the photoactive orange carotenoid protein of cyanobacteria. J. Biol. Chem. 2010, 285, 18364–18375. [Google Scholar] [CrossRef] [Green Version]
- Kirilovsky, D.; Kerfeld, C.A. The Orange Carotenoid Protein: A blue-green light photoactive protein. Photochem. Photobiol. Sci. 2013, 12, 1135–1143. [Google Scholar] [CrossRef]
- Kirilovsky, D.; Kerfeld, C.A. Cyanobacterial photoprotection by the orange carotenoid protein. Nat. Plants 2016, 2, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mori, Y. Computational study on the color change of 3′-hydroxyechinenone in the orange carotenoid protein. Chem. Phys. Lett. 2016, 652, 184–189. [Google Scholar] [CrossRef]
- Boulay, C.; Wilson, A.; Haene, S.D.; Kirilovsky, D. Identification of a protein required for recovery of full antenna capacity in OCP-related photoprotective mechanism in cyanobacteria. Proc. Natl. Acad. Sci. USA 2010, 107, 11620–11625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Igual, R.; Wilson, A.; Leverenz, R.L.; Melnicki, M.R.; de Carbon, C.B.; Sutter, M.; Turmo, A.; Perreau, F.; Kerfeld, C.A.; Kirilovsky, D. Different functions of the paralogs to the N-terminal domain of the orange carotenoid protein in the cyanobacterium Anabaena sp. PCC 7120. Plant Physiol. 2016, 171, 1852–1866. [Google Scholar] [CrossRef] [Green Version]
- Staleva, H.; Komenda, J.; Shukla, M.K.; Šlouf, V.; Kaňa, R.; Polívka, T.; Sobotka, R. Mechanism of photoprotection in the cyanobacterial ancestor of plant antenna proteins. Nat. Chem. Biol. 2015, 11, 287–291. [Google Scholar] [CrossRef]
- Sinha, R.K.; Komenda, J.; Knoppova, J.; Sedlářová, M.; Pospíšil, P. Small CAB-like proteins prevent formation of singlet oxygen in the damaged photosystem II complex of the cyanobacterium Synechocystis sp. PCC 6803. Plant. Cell Environ. 2012, 35, 806–818. [Google Scholar] [CrossRef]
- Chidgey, J.W.; Linhartová, M.; Komenda, J.; Jackson, P.J.; Dickman, M.J.; Canniffe, D.P.; Koník, P.; Pilný, J.; Hunter, C.N.; Sobotka, R. A cyanobacterial chlorophyll synthase-HliD complex associates with the Ycf39 protein and the YidC/Alb3 insertase. Plant Cell 2014, 26, 1267–1279. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Jantaro, S.; Lu, B.; Majeed, W.; Bailey, M.; He, Q. The high light-inducible polypeptides stabilize trimeric photosystem I complex under high light conditions in Synechocystis PCC 6803. Plant Physiol. 2008, 147, 1239–1250. [Google Scholar] [CrossRef] [Green Version]
- Engelken, J.; Funk, C.; Adamska, I. The Extended Light-Harvesting Complex (Lhc) Protein Superfamily: Classification and Evolutionary Dynamics BT. In Functional Genomics and Evolution of Photosynthetic Systems; Burnap, R., Vermaas, W., Eds.; Springer: Dordrecht, Netherlands, 2012; pp. 265–284. [Google Scholar]
- Beck, J.; Lohscheider, J.N.; Albert, S.; Andersson, U.; Mendgen, K.W.; Rojas-Stütz, M.C.; Adamska, I.; Funck, D. Small one-helix proteins are essential for photosynthesis in Arabidopsis. Front. Plant Sci. 2017, 8, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Psencik, J.; Hey, D.; Grimm, B.; Lokstein, H. Photoprotection of photosynthetic pigments in Plant One-Helix Protein 1/2 heterodimers. J. Phys. Chem. Lett. 2020, 11, 9387–9392. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Takabayashi, A.; Tanaka, A.; Tanaka, R. Functional analysis of light-harvesting-like protein 3 (LIL3) and its light-harvesting chlorophyll-binding motif in Arabidopsis. J. Biol. Chem. 2014, 289, 987–999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rensing, S.A.; Lang, D.; Zimmer, A.D.; Terry, A.; Salamov, A.; Shapiro, H.; Nishiyama, T.; Perroud, P.F.; Lindquist, E.A.; Kamisugi, Y.; et al. The Physcomitrella genome reveals evolutionary insights into the conquest of land by plants. Science 2008, 319, 64–69. [Google Scholar] [CrossRef] [Green Version]
- Meyer, G.; Kloppstech, K. A rapidly light-induced chloroplast protein with a high turnover coded for by pea nuclear DNA. Eur. J. Biochem. 1984, 138, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Adamska, I.; Kloppstech, K.; Ohad, I. UV light stress induces the synthesis of the early light-inducible protein and prevents its degradation. J. Biol. Chem. 1992, 267, 24732–24737. [Google Scholar] [CrossRef]
- Pötter, E.; Kloppstech, K. Effects of light stress on the expression of early light-inducible proteins in barley. Eur. J. Biochem. 1993, 214, 779–786. [Google Scholar] [CrossRef] [PubMed]
- Heddad, M.; Norén, H.; Reiser, V.; Dunaeva, M.; Andersson, B.; Adamska, I. Differential expression and localization of early light-induced proteins in Arabidopsis. Plant Physiol. 2006, 142, 75–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adamska, I.; Ohad, I.; Kloppstech, K. Synthesis of the early light-inducible protein is controlled by blue light and related to light stress. Proc. Natl. Acad. Sci. USA 1992, 89, 2610–2613. [Google Scholar] [CrossRef] [Green Version]
- Kleine, T.; Kindgren, P.; Benedict, C.; Hendrickson, L.; Strand, Å. Genome-wide gene expression analysis reveals a critical role for CRYPTOCHROME1 in the response of Arabidopsis to high irradiance. Plant Physiol. 2007, 144, 1391–1406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sävenstrand, H.; Olofsson, M.; Samuelsson, M.; Strid, Å. Induction of early light-inducible protein gene expression in Pisum sativum after exposure to low levels of UV-B irradiation and other environmental stresses. Plant Cell Rep. 2004, 22, 532–536. [Google Scholar] [CrossRef] [PubMed]
- Adamska, I.; Roobol-Bóza, M.; Lindahl, M.; Andersson, B. Isolation of pigment-binding early light-inducible proteins from pea. Eur. J. Biochem. 1999, 260, 453–460. [Google Scholar] [CrossRef] [PubMed]
- Truong, T.B. Investigating the Role (s) of LhcSRs in Chlamydomonas Reinhardtii. Ph.D. Thesis, University California Berkeley, Berkeley, CA, USA, 2011. [Google Scholar]
- Perozeni, F.; Beghini, G.; Cazzaniga, S.; Ballottari, M. Chlamydomonas reinhardtii LhcSR1 and LhcSR3 proteins involved in photoprotective non-photochemical quenching have different quenching efficiency and different carotenoid affinity. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef]
- Girolomoni, L.; Cazzaniga, S.; Pinnola, A.; Perozeni, F.; Ballottari, M.; Bassi, R. LhcSR3 is a nonphotochemical quencher of both photosystems in Chlamydomonas reinhardtii. Proc. Natl. Acad. Sci. USA 2019, 116, 4212–4217. [Google Scholar] [CrossRef] [Green Version]
- Roach, T.; Na, C.S. LhcSR3 affects de-coupling and re-coupling of LhcII to PSII during state transitions in Chlamydomonas reinhardtii. Sci. Rep. 2017, 7, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Maruyama, S.; Tokutsu, R.; Minagawa, J. Transcriptional regulation of the stress-responsive light harvesting complex genes in Chlamydomonas reinhardtii. Plant Cell Physiol. 2014, 55, 1304–1310. [Google Scholar] [CrossRef]
- Tokutsu, R.; Fujimura-Kamada, K.; Yamasaki, T.; Okajima, K.; Minagawa, J. UV-A/B radiation rapidly activates photoprotective mechanisms in Chlamydomonas reinhardtii. Plant Physiol. 1996, 47, 655–684. [Google Scholar]
- Allorent, G.; Tokutsu, R.; Roach, T.; Peers, G.; Cardol, P.; Girard-Bascou, J.; Seigneurin-Berny, D.; Petroutsos, D.; Kuntz, M.; Breyton, C.; et al. A dual strategy to cope with high light in Chlamydomonas reinhardtii. Plant Cell 2013, 25, 545–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allorent, G.; Lefebvre-Legendre, L.; Chappuis, R.; Kuntz, M.; Truong, T.B.; Niyogi, K.K.; Ulm, R.; Goldschmidt-Clermont, M. UV-B photoreceptor-mediated protection of the photosynthetic machinery in Chlamydomonas reinhardtii. Proc. Natl. Acad. Sci. USA 2016, 113, 14864–14869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, H.; Bergner, S.V.; Scholz, M.; Hippler, M. Novel insights into the function of LhcSR3 in Chlamydomonas reinhardtii. Plant Signal. Behav. 2015, 10, e1058462. [Google Scholar] [CrossRef]
- Girolomoni, L.; Ferrante, P.; Berteotti, S.; Giuliano, G.; Bassi, R.; Ballottari, M. The function of LhcBM4/6/8 antenna proteins in Chlamydomonas reinhardtii. J. Exp. Bot. 2017, 68, 627–641. [Google Scholar] [CrossRef]
- Semchonok, D.A.; Yadav, K.N.S.; Xu, P.; Drop, B.; Croce, R.; Boekema, E.J. Interaction between the photoprotective protein LhcSR3 and C2S2 Photosystem II supercomplex in Chlamydomonas reinhardtii. Biochim. Biophys. Acta BBA Bioenerg. 2017, 1858, 379–385. [Google Scholar] [CrossRef]
- Horton, P.; Ruban, A.V.; Walters, R.G. Regulation of light harvesting in green plants. Annu. Rev. Plant Biol. 1996, 47, 655–684. [Google Scholar] [CrossRef] [PubMed]
- DallOsto, L.; Cazzaniga, S.; Bressan, M.; Paleček, D.; Židek, K.; Niyogi, K.K.; Fleming, G.R.; Zigmantas, D.; Bassi, R. Two mechanisms for dissipation of excess light in monomeric and trimeric light-harvesting complexes. Nat. Plants 2017, 3, 1–9. [Google Scholar] [CrossRef]
- Townsend, J.; Saccon, F.; Giovagnetti, V.; Wilson, S.; Ungerer, P.; Ruban, A.V. The causes of altered chlorophyll fluorescence quenching induction in the Arabidopsis mutant lacking all minor antenna complexes. Biochim. Biophys. Acta BBA Bioenerg. 2018, 1859, 666–675. [Google Scholar] [CrossRef]
- Barera, S.; Pagliano, C.; Pape, T.; Saracco, G.; Barber, J. Characterization of PSII–LhcII supercomplexes isolated from pea thylakoid membrane by one-step treatment with α-and β-dodecyl-d-maltoside. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 3389–3399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albanese, P.; Manfredi, M.; Marengo, E.; Saracco, G.; Pagliano, C. Structural and functional differentiation of the light-harvesting protein Lhcb4 during land plant diversification. Physiol. Plant. 2019, 166, 336–350. [Google Scholar] [CrossRef]
- Grinzato, A.; Albanese, P.; Marotta, R.; Swuec, P.; Saracco, G.; Bolognesi, M.; Zanotti, G.; Pagliano, C. High-Light versus Low-Light: Effects on Paired Photosystem II Supercomplex Structural Rearrangement in Pea Plants. Int. J. Mol. Sci. 2020, 21, 8643. [Google Scholar] [CrossRef]
- De Bianchi, S.; DallOsto, L.; Tognon, G.; Morosinotto, T.; Bassi, R. Minor antenna proteins CP24 and CP26 affect the interactions between photosystem II subunits and the electron transport rate in grana membranes of Arabidopsis. Plant Cell 2008, 20, 1012–1028. [Google Scholar] [CrossRef] [Green Version]
- De Bianchi, S.; Betterle, N.; Kouril, R.; Cazzaniga, S.; Boekema, E.; Bassi, R.; Dall’Osto, L. Arabidopsis mutants deleted in the light-harvesting protein Lhcb4 have a disrupted photosystem II macrostructure and are defective in photoprotection. Plant Cell 2011, 23, 2659–2679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinnola, A.; Alboresi, A.; Nosek, L.; Semchonok, D.; Rameez, A.; Trotta, A.; Barozzi, F.; Kouřil, R.; Dall’Osto, L.; Aro, E.M.; et al. A LhcB9-dependent photosystem I megacomplex induced under low light in Physcomitrella patens. Nat. Plants 2018, 4, 910–919. [Google Scholar] [CrossRef]
- Croce, R.; van Amerongen, H. Light harvesting in oxygenic photosynthesis: Structural biology meets spectroscopy. Science 2020, 369, eaay2058. [Google Scholar] [CrossRef] [PubMed]
- Elrad, D.; Grossman, A.R. A genomes-eye view of the light-harvesting polypeptides of Chlamydomonas reinhardtii. Curr. Genet. 2004, 45, 61–75. [Google Scholar] [CrossRef]
- Peers, G.; Truong, T.B.; Ostendorf, E.; Busch, A.; Elrad, D.; Grossman, A.R.; Hippler, M.; Niyogi, K.K. An ancient light-harvesting protein is critical for the regulation of algal photosynthesis. Nature 2009, 462, 518–521. [Google Scholar] [CrossRef] [PubMed]
- Bonente, G.; Ballottari, M.; Truong, T.B.; Morosinotto, T.; Ahn, T.K.; Fleming, G.R.; Niyogi, K.K.; Bassi, R. Analysis of LhcSR3, a protein essential for feedback de-excitation in the green alga Chlamydomonas reinhardtii. PLoS Biol. 2011, 9, e1000577. [Google Scholar] [CrossRef] [Green Version]
- Ferrante, P.; Ballottari, M.; Bonente, G.; Giuliano, G.; Bassi, R. LhcBM1 and LhcBM2/7 polypeptides, components of major LhcII complex, have distinct functional roles in photosynthetic antenna system of Chlamydomonas reinhardtii. J. Biol. Chem. 2012, 287, 16276–16288. [Google Scholar] [CrossRef] [Green Version]
- Pan, X.; Tokutsu, R.; Li, A.; Takizawa, K.; Song, C.; Murata, K.; Yamasaki, T.; Liu, Z.; Minagawa, J.; Li, M. Structural basis of LhcbM5-mediated state transitions in green algae. bioRxiv 1101, 2021.03.02.433643. [Google Scholar] [CrossRef]
- Nguyen, V.; Thomas-Hall, S.R.; Malnoë, A.; Timmins, M.; Mussgnug, J.H.; Rupprecht, J.; Kruse, O.; Hankamer, B.; Schenk, P.M. Transcriptome for photobiological hydrogen production induced by sulfur deprivation in the green alga Chlamydomonas reinhardtii. Eukaryot. Cell 2008, 7, 1965–1979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grewe, S.; Ballottari, M.; Alcocer, M.; D’Andrea, C.; Blifernez-Klassen, O.; Hankamer, B.; Mussgnug, J.H.; Bassi, R.; Kruse, O. Light-harvesting complex protein LhcBM9 is critical for photosystem II activity and hydrogen production in Chlamydomonas reinhardtii. Plant Cell 2014, 26, 1598–1611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pietrzykowska, M.M.; Suorsa, M.; Semchonok, D.A.; Tikkanen, M.; Boekema, E.J.; Aro, E.M.; Jansson, S. The light-harvesting chlorophyll a/b binding proteins Lhcb1 and Lhcb2 play complementary roles during state transitions in Arabidopsis. Plant Cell 2014, 26, 3646–3660. [Google Scholar] [CrossRef] [Green Version]
- Boekema, E.J.; van Roon, H.; van Breemen, J.F.L.; Dekker, J.P. Supramolecular organization of photosystem II and its light-harvesting antenna in partially solubilized photosystem II membranes. Eur. J. Biochem. 1999, 266, 444–452. [Google Scholar] [CrossRef] [Green Version]
- Damkjær, T.; Kereïche, S.; Johnson, M.P.; Kovacs, L.; Kiss, A.Z.; Boekema, E.J.; Ruban, A.V.; Horton, P.; Jansson, S. The photosystem II light-harvesting protein Lhcb3 affects the macrostructure of photosystem II and the rate of state transitions in Arabidopsis. Plant Cell 2009, 21, 3245–3256. [Google Scholar] [CrossRef] [Green Version]
- Kouřil, R.; Nosek, L.; Bartoš, J.; Boekema, E.J.; Ilík, P. Evolutionary loss of light-harvesting proteins Lhcb6 and Lhcb3 in major land plant groups–break-up of current dogma. New Phytol. 2016, 210, 808–814. [Google Scholar] [CrossRef] [Green Version]
- Mozzo, M.; Mantelli, M.; Passarini, F.; Caffarri, S.; Croce, R.; Bassi, R. Functional analysis of Photosystem I light-harvesting complexes (Lhca) gene products of Chlamydomonas reinhardtii. Biochim. Biophys. ActaBioenerg. 2010, 1797, 212–221. [Google Scholar] [CrossRef] [Green Version]
- Wientjes, E.; Croce, R. The light-harvesting complexes of higher-plant Photosystem I: Lhca1/4 and Lhca2/3 form two red-emitting heterodimers. Biochem. J. 2011, 433, 477–485. [Google Scholar] [CrossRef] [Green Version]
- Castelletti, S.; Morosinotto, T.; Robert, B.; Caffarri, S.; Bassi, R.; Croce, R. Recombinant Lhca2 and Lhca3 subunits of the photosystem I antenna system. Biochemistry 2003, 42, 4226–4234. [Google Scholar] [CrossRef]
- Wientjes, E.; Oostergetel, G.T.; Jansson, S.; Boekema, E.J.; Croce, R. The role of Lhca complexes in the supramolecular organization of higher plant photosystem I. J. Biol. Chem. 2009, 284, 7803–7810. [Google Scholar] [CrossRef] [Green Version]
- Yadav, N.S.; Semchonok, D.A.; Nosek, L.; Kouřil, R.; Fucile, G.; Boekema, E.J.; Eichacker, L.A. Supercomplexes of plant photosystem I with cytochrome b6f, light-harvesting complex II and NDH. Biochim. Biophys. Acta Bioenerg. 2017, 1858, 12–20. [Google Scholar] [CrossRef]
- Peng, Y.; Fukao, M.; Fujiwara, T.; Shikanai, T. Efficient operation of NAD(P)H dehydrogenase requires supercomplex formation with photosystem I via minor LhcI in arabidopsis. Plant Cell 2009, 21, 3623–3640. [Google Scholar] [CrossRef] [Green Version]
- Xu, P.; Tian, L.; Kloz, M.; Croce, R. Molecular insights into Zeaxanthin-dependent quenching in higher plants. Sci. Rep. 2015, 5, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Steen, C.J.; Morris, J.M.; Short, A.H.; Niyogi, K.K.; Fleming, G.R. Complex Roles of PsbS and Xanthophylls in the Regulation of Nonphotochemical Quenching in Arabidopsis thaliana under Fluctuating Light. J. Phys. Chem. B 2020, 124, 10311–10325. [Google Scholar] [CrossRef] [PubMed]
- Nicol, W.; Nawrocki, J.; Croce, R. Disentangling the sites of non-photochemical quenching in vascular plants. Nat. Plants 2019, 5, 1177–1183. [Google Scholar] [CrossRef]
- Pawlak, P.S.; Liu, C.; Reus, M.; Yang, C.; Holzwarth, A.R. On the PsbS-induced quenching in the plant major light-harvesting complex LhcII studied in proteoliposomes. Photosynth. Res. 2020, 144, 195–208. [Google Scholar] [CrossRef]
- Haniewicz, P.; De Sanctis, D.; Büchel, C.; Schröder, W.P.; Loi, M.C.; Kieselbach, T.; Bochtler, M.; Piano, D. Isolation of monomeric photosystem II that retains the subunit PsbS. Photosynth. Res. 2013, 118, 199–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ware, A.; Giovagnetti, V.; Belgio, E.; Ruban, A.V. PsbS protein modulates non-photochemical chlorophyll fluorescence quenching in membranes depleted of photosystems. J. Photochem. Photobiol. B Biol. 2015, 152, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Roach, T.; Krieger-Liszkay, A. The role of the PsbS protein in the protection of photosystems I and II against high light in Arabidopsis thaliana. Biochim. Biophys. Acta-Bioenerg. 2012, 1817, 2158–2165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tibiletti, T.; Auroy, P.; Peltier, G.; Caffarri, S. Chlamydomonas reinhardtii PsbS protein is functional and accumulates rapidly and transiently under high light. Plant Physiol. 2016, 171, 2717–2730. [Google Scholar] [CrossRef] [Green Version]
- Correa-Galvis, V.; Redekop, P.; Guan, K.; Griess, A.; Truong, T.B.; Wakao, S.; Niyogi, K.K.; Jahns, P. Photosystem II subunit PsbS is involved in the induction of LhcSR protein-dependent energy dissipation in Chlamydomonas reinhardtii. J. Biol. Chem. 2016, 291, 17478–17487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Głowacka, K.; Kromdijk, J.; Kucera, K.; Xie, J.; Cavanagh, A.P.; Leonelli, L.; Leakey, A.D.; Ort, D.R.; Niyogi, K.K.; Long, S.P. Photosystem II Subunit S overexpression increases the efficiency of water use in a field-grown crop. Nat. Commun. 2018, 9, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ulloa, O.; Henríquez-Castillo, C.; Ramírez-Flandes, S.; Plominsky, A.M.; Murillo, A.A.; Morgan-Lang, C.; Hallam, S.J.; Stepanauskas, R. The cyanobacterium Prochlorococcus has divergent light-harvesting antennae and may have evolved in a low-oxygen ocean. Proc. Natl. Acad. Sci. USA 2021, 118, e2025638118. [Google Scholar] [CrossRef] [PubMed]
- Treves, H.; Raanan, H.; Kedem, I.; Murik, O.; Keren, N.; Zer, H.; Berkowicz, S.M.; Giordano, M.; Norici, A.; Shotland, Y.; et al. The mechanisms whereby the green alga Chlorella ohadii, isolated from desert soil crust, exhibits unparalleled photodamage resistance. New Phytol. 2016, 210, 1229–1243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Treves, H.; Siemiatkowska, B.; Luzarowska, U.; Murik, O.; Fernandez-Pozo, N.; Moraes, T.A.; Erban, A.; Armbruster, U.; Brotman, Y.; Kopka, J.; et al. Multi-omics reveals mechanisms of total resistance to extreme illumination of a desert alga. Nat. Plants 2020, 6, 1031–1043. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bag, P. Light Harvesting in Fluctuating Environments: Evolution and Function of Antenna Proteins across Photosynthetic Lineage. Plants 2021, 10, 1184. https://doi.org/10.3390/plants10061184
Bag P. Light Harvesting in Fluctuating Environments: Evolution and Function of Antenna Proteins across Photosynthetic Lineage. Plants. 2021; 10(6):1184. https://doi.org/10.3390/plants10061184
Chicago/Turabian StyleBag, Pushan. 2021. "Light Harvesting in Fluctuating Environments: Evolution and Function of Antenna Proteins across Photosynthetic Lineage" Plants 10, no. 6: 1184. https://doi.org/10.3390/plants10061184