
Effects of Interspecific Chromosome Substitution in Upland Cotton on Cottonseed Macronutrients


 , , , and
, , , and
Abstract
:1. Introduction
2. Results
2.1. CS Lines with High Level of Nutrients in Both Locations
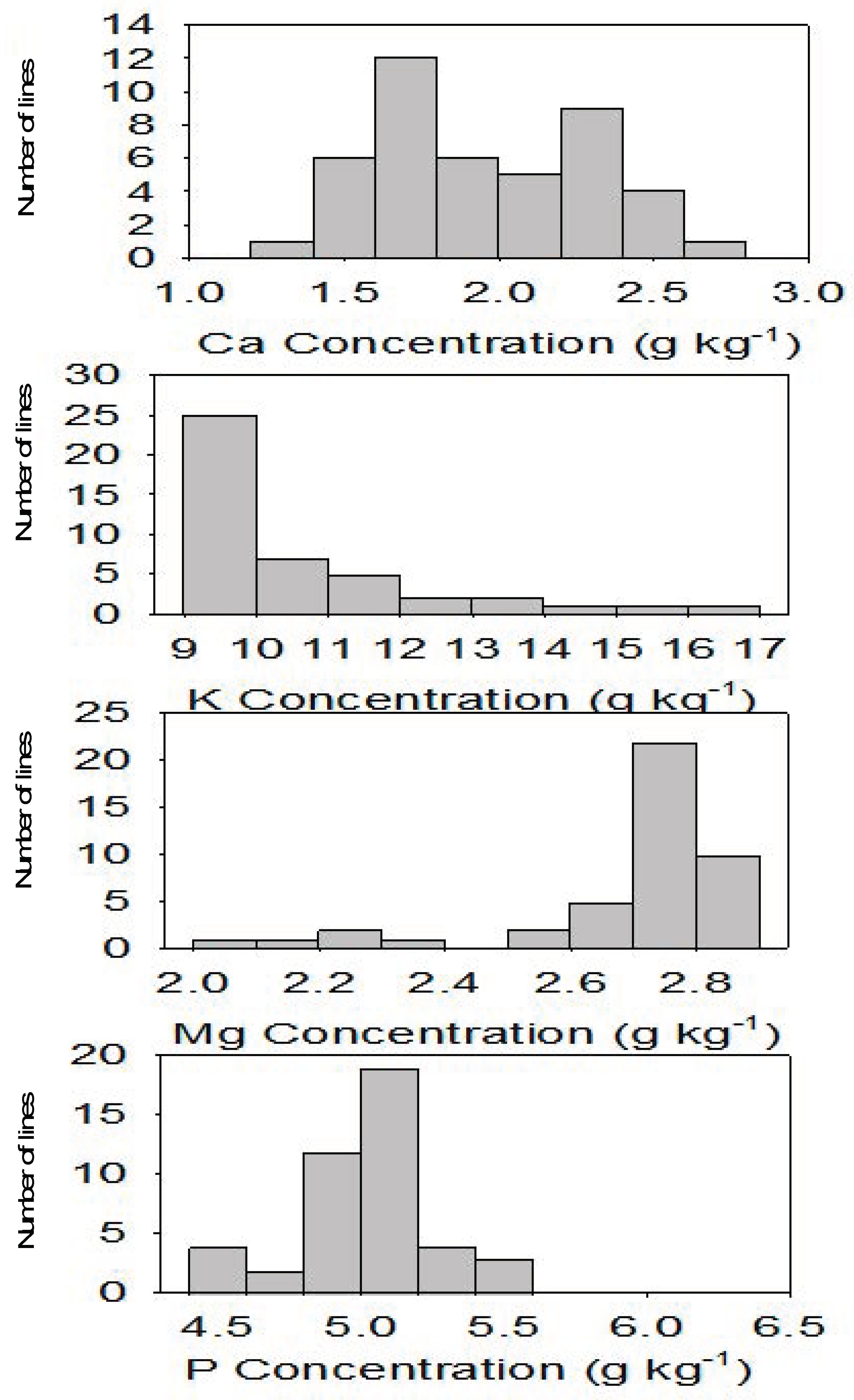
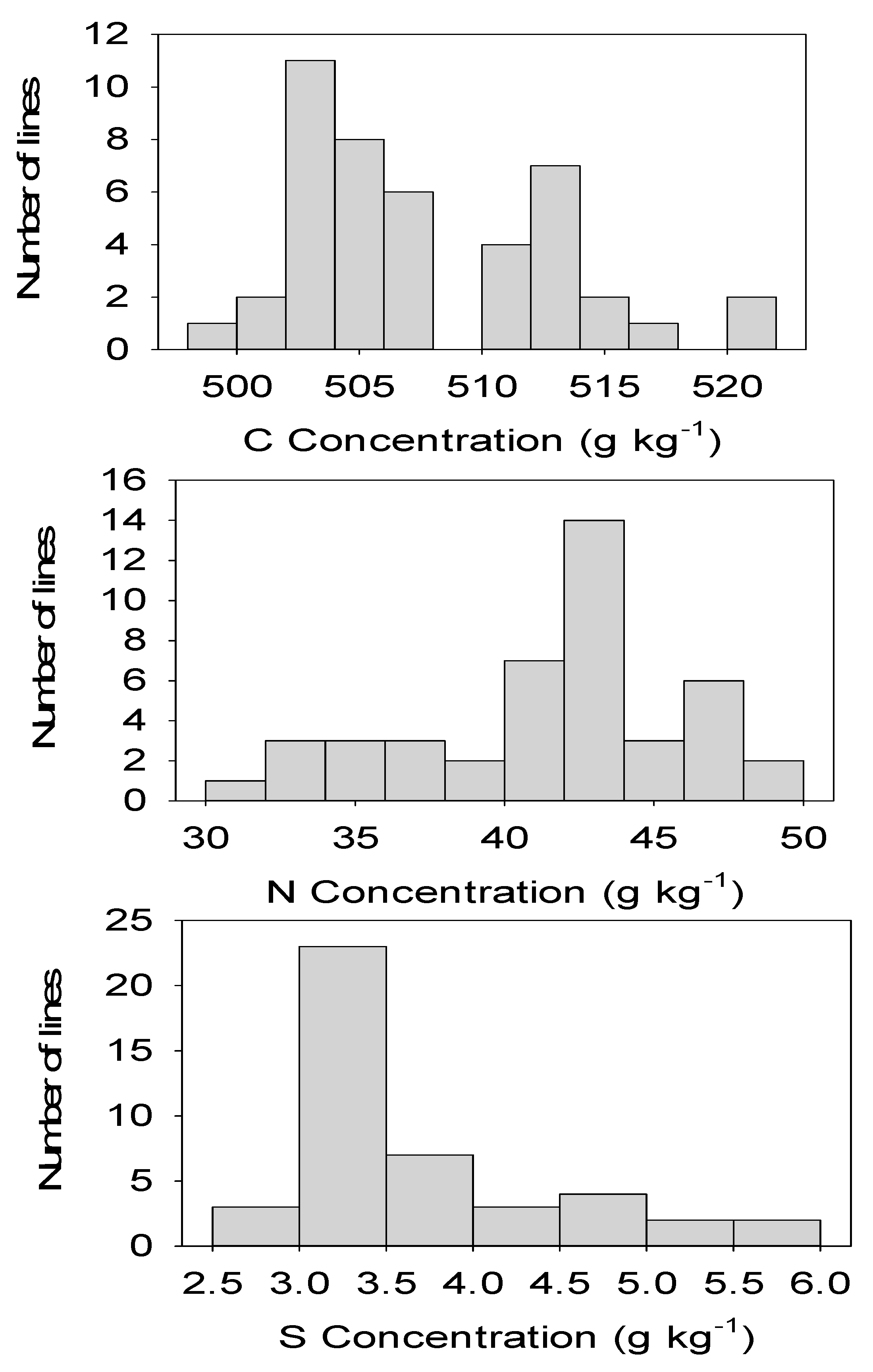
2.2. Correlation and Distribution
3. Discussion
4. Materials and Methods
4.1. Chromosome Substitution Cotton Lines (CS)
4.2. Seed Mineral Nutrient Analyses
4.3. Determination of Seed P
4.4. Experimental Design and Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sawan, Z.M.; Hafez, S.A.; Basyony, A.E.; Alkassas, A.E.E. Cottonseed, protein, oil yields and oil properties as influenced by potassium fertilization and foliar application of zinc and phosphorus. World J. Agric. Sci. 2006, 2, 66–74. [Google Scholar] [CrossRef] [Green Version]
- Mengel, K.; Kirkby, E.A. Principles of Plant Nutrition, 3rd ed.; International Potash Institute: Worblaufen-Bern, Switzerland, 1982. [Google Scholar]
- Rodriguez, D.; Zubillaga, M.M.; Ploschuck, E.; Keltjens, W.; Goudriaan, J.; Lavado, R. Leaf area expansion and assimilate prediction in sunflower growing under low phosphorus conditions. Plant Soil 1998, 202, 133–147. [Google Scholar] [CrossRef]
- Taiz, L.; Zeiger, E. Plant Physiology: Mineral Nutrition; The Benjamin Cummings Publishing: Redwood City, CA, USA, 1991. [Google Scholar]
- Rajeswari, R.V. Foliar application of growth regulators and nutrients on boll development and yield in cotton. J. Ind. Soc Cotton. Improv. 1996, 21, 71. [Google Scholar]
- Rathinavel, K.; Dharmalingam, C.; Paneersewam, S. Effect of micronutrient on the productivity and quality of cotton seed Cv. TCB 209 (Gossypium barbadense L.). Madras Agric. J. 1999, 86, 313–316. [Google Scholar]
- Ahlawat, I.P.S. A note on response of cotton (Gossypium hirsutum) to soil and foliar application of micronutrients. Ind. J. Agron. 1974, 19, 71–74. [Google Scholar]
- Khuzhanazarov, S.H.; Dzhumaev, N.; Mamanov, S.H. Effect of copper and zinc in the composition of basal fertilizer on yield of fine fibred cotton. Field Crop Abstr. 1983, 37, 780. [Google Scholar] [CrossRef] [Green Version]
- WHO. Guideline: Potassium Intake for Adults and Children; World Health Organization: Geneva, Switzerland, 2009; Available online: https://www.who.int/elena/titles/guidance_summaries/potassium_intake/en/ (accessed on 6 November 2020).
- WHO. Calcium Supplementation during Pregnancy to Reduce the Risk of Pre-Eclampsia. 2019. Available online: https://www.who.int/elena/titles/calcium_pregnancy/en/ (accessed on 6 November 2020).
- Grusak, M.A.; DellaPenna, D. Improving the nutrient composition of plants to enhance human nutrition and health. Ann. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 133–161. [Google Scholar] [CrossRef] [PubMed]
- White, P.J.; Broadley, M.R. Biofortification of Crops with Seven Mineral Elements Often Lacking in Human Diets—Iron, Zinc, Copper, Calcium, Magnesium, Selenium and Iodine. New Phytol. 2009, 182, 49–84. [Google Scholar] [CrossRef] [PubMed]
- Bellaloui, N.; Turley, R.B.; Stetina, S.R. Cottonseed protein, oil, and minerals in cotton (Gossypium hirsutum L.) lines differing in curly leaf morphology. Plants 2020, 10, 525. [Google Scholar] [CrossRef]
- Marschner, P. Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Academic Press: San Diego, CA, USA, 2012. [Google Scholar]
- Brown, P.H.; Bellaloui, N.; Wimmer, M.A.; Bassil, E.S.; Ruiz, J.; Hu, H. Boron in plant biology. Plant Biol. 2002, 4, 205–223. [Google Scholar] [CrossRef]
- Garcia-Oliveira, A.L.; Tan, L.; Fu, Y.; Sun, C. Genetic identification of quantitative trait loci for contents of mineral nutrients in rice grain. J. Integr. Plant Biol. 2009, 51, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Peleg, Z.; Cakmak, I.; Ozturk, L. Quantitative trait loci conferring grain mineral nutrient concentrations in durum wheat×wild emmer wheat RIL population. Theor. Appl. Genet. 2009, 119, 353–369. [Google Scholar] [CrossRef] [Green Version]
- Sankaran, R.; Huguet, T.; Grusak, M. Identification of QTL affecting seed mineral concentrations and content in the model legume Medicago truncatula. Theor. Appl. Genet. 2009, 119, 241–253. [Google Scholar] [CrossRef] [PubMed]
- Bellaloui, N.; Khandaker, L.; Akond, M.; Kantartzi, S.K.; Meksem, K.; Mengistu, A.; Lightfoot, D.A.; Kassem, M.A. Genetic mapping of QTL associated with seed macronutrients accumulation in ‘MD 96-5722′ by ‘Spencer’ recombinant inbred lines of soybean. Atlas J. Biol. 2015, 3, 224–235. [Google Scholar] [CrossRef]
- Lu, K.; Li, L.; Zheng, X.; Zhang, Z.; Mou, T.; Hu, Z. Quantitative trait loci controlling Cu, Ca, Zn, Mn and Fe content in rice grains. J. Genet. 2008, 87, 305–310. [Google Scholar] [CrossRef] [PubMed]
- Paran, I.; Zamir, D. Quantitative traits in plants: Beyond the QTL. Trends Genet. 2003, 19, 303–306. [Google Scholar] [CrossRef]
- Bellaloui, N.; Khandaker, L.; Akond, M.; Kantartzi, S.K.; Meksem, K.; Mengistu, A.; Lightfoot, D.A.; Kassem, M.A. Identification of QTL underlying seed micronutrients accumulation in ‘MD 96-5722′ by ‘Spencer’ recombinant inbred lines of soybean. Atlas J. Plant Biol. 2015, 1, 39–49. [Google Scholar] [CrossRef]
- Zeng, C.; Han, Y.; Shi, L.; Peng, L.; Wang, Y.; Xu, F.; Meng, J. Genetic Analysis of the Physiological Responses to Low Boron Stress in Arabidopsis thaliana. Plant Cell Environ. 2008, 31, 112–122. [Google Scholar] [CrossRef]
- Le Gouis, J. Genetic improvement of nutrient use efficiency in wheat. In Molecular and Physiological Basis of Nutrient Use Efficiency in Crops; Hawkesford, M.J., Barraclough, P., Eds.; Wiley Blackwell: Chichester, UK, 2011; Chapter 7; pp. 123–138. [Google Scholar]
- Wu, J.; Jenkins, J.N.; McCarty, J.C.; Thaxton, P. Seed trait evaluation of Gossypium barbadense L. chromosomes/arms in a G. hirsutum L. background. Euphytica 2009, 167, 371–380. [Google Scholar] [CrossRef]
- Saha, S.; Wu, J.; Jenkins, J.N.; McCarty, J.C.; Gutierrez, O.; Stelly, D.M.; Percy, R.G.; Raska, D.A. Effect of chromosome substitutions from Gossypium barbadense L. 3–79 into G. hirsutum L. TM-1 on agronomic and fiber traits. J. Cotton Sci. 2004, 8, 162–169. [Google Scholar]
- Saha, S.; Jenkins, J.N.; Wu, J.; McCarty, J.C.; Gutierrez, O.; Percy, R.C.; Cantrell, R.G.; Stelly, D.M. Effects of chromosome specific introgression in upland cotton on fiber and agronomic traits. Genetics 2006, 172, 1927–1938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stelly, D.M.; Saha, S.; Raska, D.A.; Jenkins, J.N.; McCarty, J.C.; Gutierrez, O. Registration of 17 germplasm lines of upland cotton (Gossypium hirsutum), each with a different pair of G. barbadense chromosome or chromosome arms substituted for the respective G. hirsutum chromosome or chromosome arms. Crop Sci. 2005, 45, 2663–2665. [Google Scholar] [CrossRef]
- Jenkins, J.N.; Wu, J.; McCarty, J.C.; Saha, S.; Gutierrez, O.; Hayes, R.; Stelly, D.M. Genetic evaluation for thirteen chromosome substitution lines crossed with five commercial cultivars: I. Yield traits. Crop Sci. 2006, 46, 1169–1178. [Google Scholar] [CrossRef] [Green Version]
- Jenkins, J.N.; McCarty, J.C.; Wu, J.; Saha, S.; Gutierrez, O.; Hayes, R.; Stelly, D.M. Genetic evaluation of thirteen chromosome substitution lines crossed with five commercial cultivars: II. Fiber quality traits. Crop Sci. 2007, 47, 561–572. [Google Scholar] [CrossRef] [Green Version]
- McCarty, J.C.; Wu, J.; Saha, S.; Jenkins, J.N.; Hayes, R. Effects of chromosome 5sh from Gossypium barbadense L. on flower production in G. hirsutum L. Euphytica 2006, 152, 99–107. [Google Scholar] [CrossRef]
- Wu, J.; McCarty, J.C.; Jenkins, J.N. Cotton chromosome substitution lines crossed with cultivars: Genetic model evaluation and seed trait analyses. Theor. Appl. Genet. 2010, 120, 1473–1483. [Google Scholar] [CrossRef]
- Wu, Y.; Machado, A.C.; White, R.G.; Llewellyn, D.J.; Dennis, E.S. Expression profiling identifies genes expressed early during lint fibre initiation in cotton. Plant Cell Physiol. 2006, 47, 107–127. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Wu, B.; Singh, R.P.; Velu, G. QTL mapping for micronutrients concentration and yield component traits in a hexaploid wheat mapping population. J. Cereal Sci. 2019, 88, 57–64. [Google Scholar] [CrossRef]
- Wu, J.; Zhu, J.; Ji, D.; Xu, F. Genetic analysis of direct and maternal effects of seed traits in upland cotton. Acta Agron. Sin. 1995, 21, 659–664. (In Chinese) [Google Scholar]
- Wang, G.; Zhu, J.; Zang, R.; Xu, F.; Ji, D. Analysis of covariance components between seed and agronomy traits in upland cotton. Acta Gossypii Sin. 1996, 8, 295–300. [Google Scholar]
- Zhu, J.; Weir, B.S. Analysis of cytoplasmic and maternal effects I. A genetic model for diploid plant seeds and animals. Theor. Appl. Genet. 1994, 89, 153–159. [Google Scholar] [CrossRef]
- Zhu, J.; Weir, B.S. Analysis of cytoplasmic and maternal effects II. Genetic models for triploid endosperms. Theor. Appl. Genet. 1994, 89, 160–166. [Google Scholar] [CrossRef]
- Descalsota-Empleo, G.I.; Amparadoa, G.I.; Inabangan-Asilo, A.; Tesoro, M.A.; Stangoulisc, F.; Reinkea, J.; Swamya, R. Genetic mapping of QTL for agronomic traits and grain mineral elements in rice. Crop J. 2019, 7, 560–572. [Google Scholar] [CrossRef]
- Saha, S.; Bellaloui, N.; Jenkins, J.N.; Jack, C.; McCarty, J.C.; David, M.; Stelly, D.M. Effect of Chromosome Substitutions from Gossypium barbadense L., G. tomentosum Nutt. Ex Seem and G. mustelinum Watt into G. hirsutum L. on cottonseed protein and oil content. Euphytica 2020, 216, 218. [Google Scholar] [CrossRef]
- Bellaloui, N.; Mengistu, A.; Walker, R.R.; Young, L.D. Soybean Seed Composition Affected by Seeding Rates and Row Spacing in the Midsouth USA. Crop Sci. 2014, 54, 1782–1795. [Google Scholar] [CrossRef]
- Bellaloui, N.; Smith, J.R.; Gillen, A.M.; Ray, J.D. Effects of Maturity, Genotypic Background, and Temperature on Seed Mineral Composition in Near Isogenic Soybean Lines in the Early Soybean Production System. Crop Sci. 2011, 51, 1161–1171. [Google Scholar] [CrossRef]
- Cavell, A.J. The Colorimetric Determination of Phosphorus in Plant Materials. J. Sci. Food Agric. 1955, 6, 479–480. [Google Scholar] [CrossRef]
- SAS Institute. SAS; SAS Institute: Cary, NC, USA, 2002–2012. [Google Scholar]



{kind=link}
{kind=link}
| Effect | DF | Ca | K | Mg | P | C | N | S | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| F | p | F | p | F | p | F | p | F | p | F | p | F | p | ||
| Location | 1 | 3.86 | ns | 2.98 | ns | 139.81 | *** | 494.42 | *** | 1.46 | ns | 22.68 | *** | 22.51 | *** |
| Line | 10 | 40.63 | ns | 6.49 | *** | 5.32 | *** | 18.96 | *** | 1.23 | ns | 36.95 | *** | 31.64 | *** |
| Location × line | 10 | 26.49 | ns | 8.44 | *** | 8.81 | *** | 15.43 | *** | 1.25 | ns | 21.11 | *** | 32.18 | *** |
| Residual | 0.011 | 0.585 | 0.016 | 0.038 | 22.7 | 3.069 | 0.046 |
| Line | Ca | K | Mg | P | C | N | S |
|---|---|---|---|---|---|---|---|
| CS-B02 | 2.04D | 9.69D | 2.81AB | 5.07DE | 507CD | 42.26CD | 4.33B |
| CS-B04 | 1.88E | 9.65D | 2.73D | 5.05E | 508BC | 42.65C | 3.31D |
| CS-B08sh | 1.80F | 10.16C | 2.78DCD | 5.10CD | 505DE | 44.02B | 3.44C |
| CS-M02 | 1.49I | 9.62D | 2.78BCD | 4.82F | 508BC | 35.55E | 3.50C |
| CS-M04 | 2.14G | 10.96B | 2.75BCD | 5.15B | 506CDE | 35.28E | 3.18E |
| CS-M08sh | 1.64H | 14.50A | 2.74CD | 5.08DE | 511A | 34.00F | 3.26DE |
| CS-T02 | 2.63A | 9.67D | 2.80ABC | 5.12BC | 503E | 47.79A | 4.26B |
| CS-T04 | 2.30B | 10.41C | 2.85A | 5.38A | 506CD | 44.09B | 5.39A |
| CS-T08sh | 2.18C | 11.21B | 2.57E | 5.11CD | 511AB | 43.75B | 3.31D |
| TM-1 | 1.81F | 11.25B | 2.21F | 4.48G | 504DE | 41.92CD | 3.17E |
| AM UA48 | 1.72G | 9.74D | 2.55E | 5.10CD | 512A | 41.67D | 2.89F |
| Line | Ca | K | Mg | P | C | N | S |
|---|---|---|---|---|---|---|---|
| CS-B02 | 2.13C | 9.95 DE | 3.08 BC | 6.11 C | 503 G | 41.19 D | 3.38 CD |
| CS-B04 | 2.00D | 10.12 CDE | 2.94 D | 5.51 F | 505 DE | 35.59 E | 3.46 BC |
| CS-B08sh | 1.70F | 10.48 BC | 2.98 D | 5.43 F | 507 BCD | 45.75 A | 4.65 A |
| CS-M02 | 1.95D | 10.95 AB | 2.83 E | 5.95 D | 507 BCD | 34.00 G | 3.21 EF |
| CS-M04 | 1.88E | 10.33 CDE | 3.04 C | 6.25 B | 503 FG | 33.25 H | 3.46 BC |
| CS-M08sh | 2.55A | 10.16 CDE | 2.96 D | 5.73 E | 507 BCD | 45.00 B | 3.50 B |
| CS-T02 | 2.19B | 10.25 CDE | 2.80 E | 6.25 B | 506 CDE | 45.91 A | 3.46 BC |
| CS-T04 | 2.24B | 10.08 DE | 3.21 A | 6.68 A | 507 BCD | 34.75 F | 3.30 DE |
| CS-T08sh | 1.98D | 9.86 E | 3.27 A | 6.63 A | 505 EF | 34.50 FG | 2.84 G |
| TM-1 | 2.09C | 10.29 CDE | 3.13 B | 6.10 C | 511 A | 42.47 C | 3.14 F |
| AM UA48 | 1.51G | 10.86 AB | 2.85 E | 4.95 G | 508 B | 41.02 D | 3.25 E |
| Ca | K | Mg | P | C | N | |
|---|---|---|---|---|---|---|
| K | R = −0.191 | |||||
| p = ns | ||||||
| Mg | R = 0.190 | −0.188 | ||||
| p = ns | ns | |||||
| P | R = 0.441 | −0.071 | 0.668 | |||
| p = *** | ns | *** | ||||
| C | R = −0.237 | 0.152 | −0.208 | 0.045 | ||
| p= ns | ns | ns | ns | |||
| N | R = 0.560 | −0.430 | −0.074 | 0.119 | −0.043 | |
| p = *** | *** | ns | ns | ns | ||
| S | R = 0.546 | −0.213 | 0.410 | 0.436 | −0.190 | 0.401 |
| p = *** | ns | *** | *** | ns | *** |
| Ca | Ca | K | Mg | P | C | N |
|---|---|---|---|---|---|---|
| K | R = −0.262 | |||||
| p = ns | ||||||
| Mg | R = 0.191 | −0.125 | ||||
| p = ns | ns | |||||
| P | R = 0.414 | −0.196 | 0.569 | |||
| p = *** | ns | *** | ||||
| C | R = −0.052 | 0.053 | −0.109 | −0.070 | ||
| p = ns | ns | ns | ns | |||
| N | R = 0.146 | 0.057 | −0.294 | −0.340 | 0.050 | |
| p = ns | ns | * | * | ns | ||
| S | R = −0.169 | −0.011 | −0.136 | −0.356 | −0.138 | 0.450 |
| p = ns | ns | ns | * | ns | *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bellaloui, N.; Saha, S.; Tonos, J.L.; Scheffler, J.A.; Jenkins, J.N.; McCarty, J.C.; Stelly, D.M. Effects of Interspecific Chromosome Substitution in Upland Cotton on Cottonseed Macronutrients. Plants 2021, 10, 1158. https://doi.org/10.3390/plants10061158
Bellaloui N, Saha S, Tonos JL, Scheffler JA, Jenkins JN, McCarty JC, Stelly DM. Effects of Interspecific Chromosome Substitution in Upland Cotton on Cottonseed Macronutrients. Plants. 2021; 10(6):1158. https://doi.org/10.3390/plants10061158
Chicago/Turabian StyleBellaloui, Nacer, Sukumar Saha, Jennifer L. Tonos, Jodi A. Scheffler, Johnie N. Jenkins, Jack C. McCarty, and David M. Stelly. 2021. "Effects of Interspecific Chromosome Substitution in Upland Cotton on Cottonseed Macronutrients" Plants 10, no. 6: 1158. https://doi.org/10.3390/plants10061158