
Raw and Fermented Alfalfa Brown Juice Induces Changes in the Germination and Development of French Marigold (Tagetes patula L.) Plants

 ,
,  and
and
Abstract
:1. Introduction
2. Results
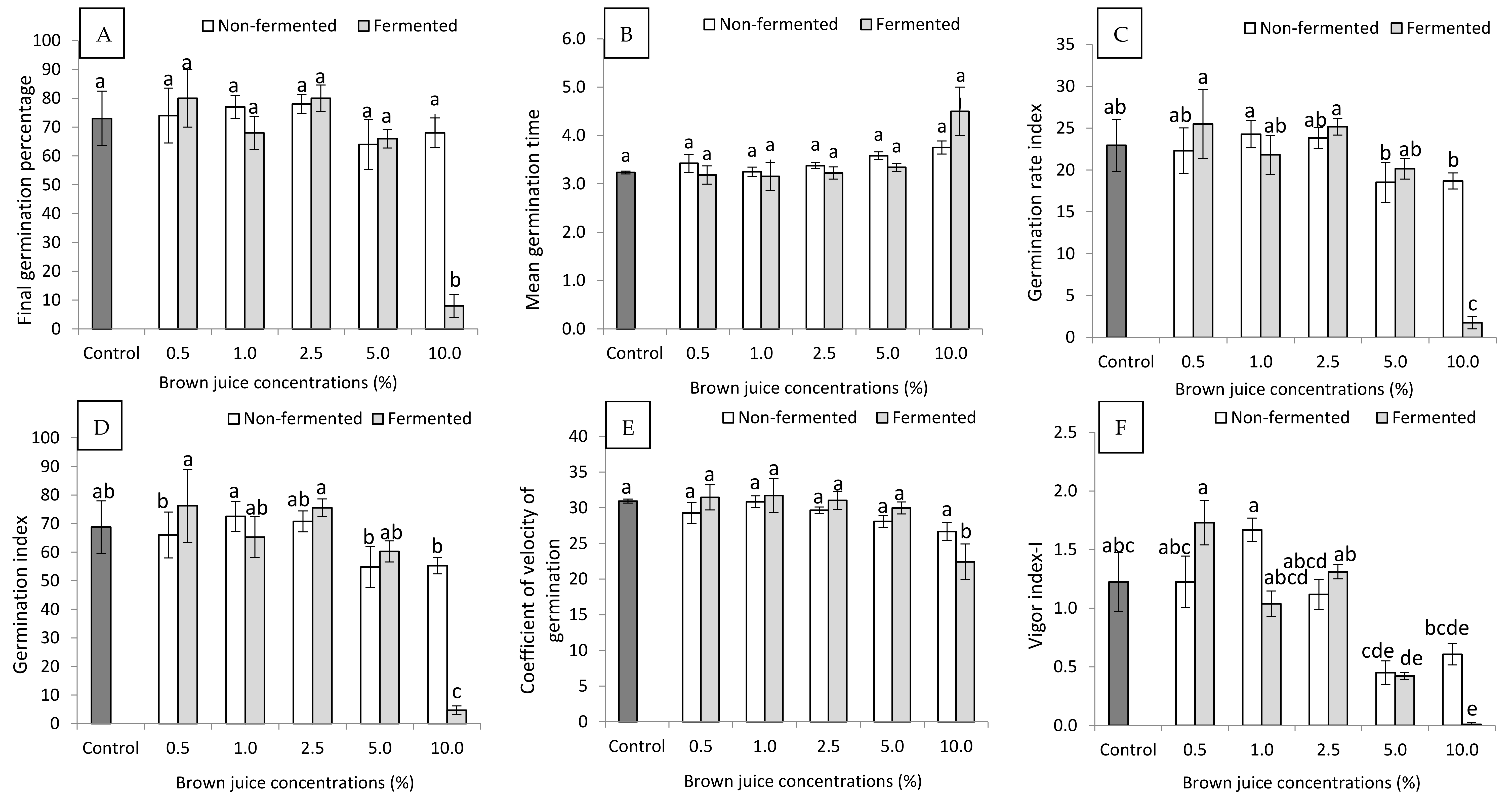
2.1. Effect of Non-Fermented and Fermented BJ on Seed Germination
2.2. Development of French Marigold Plants Treated with BJ
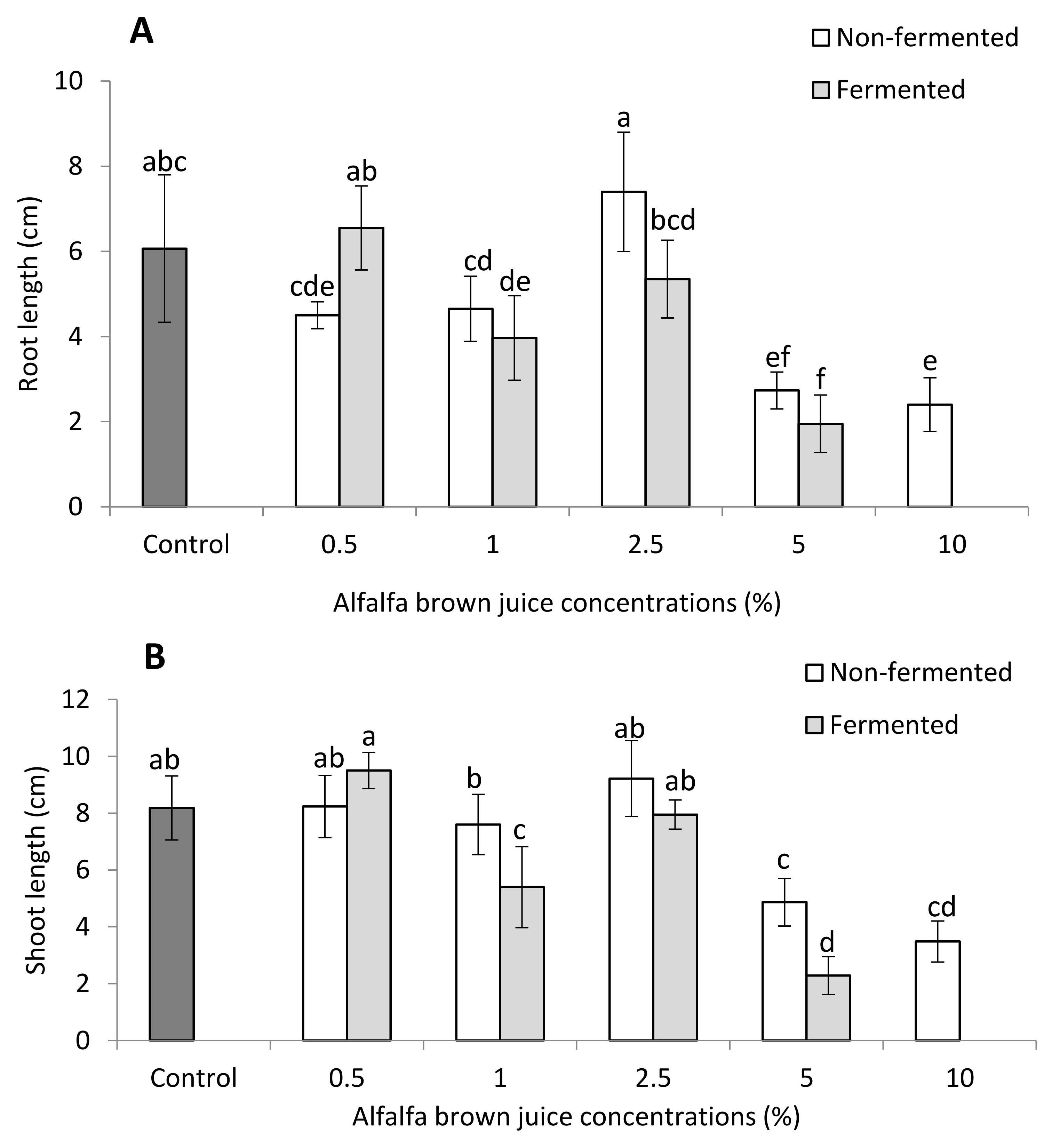
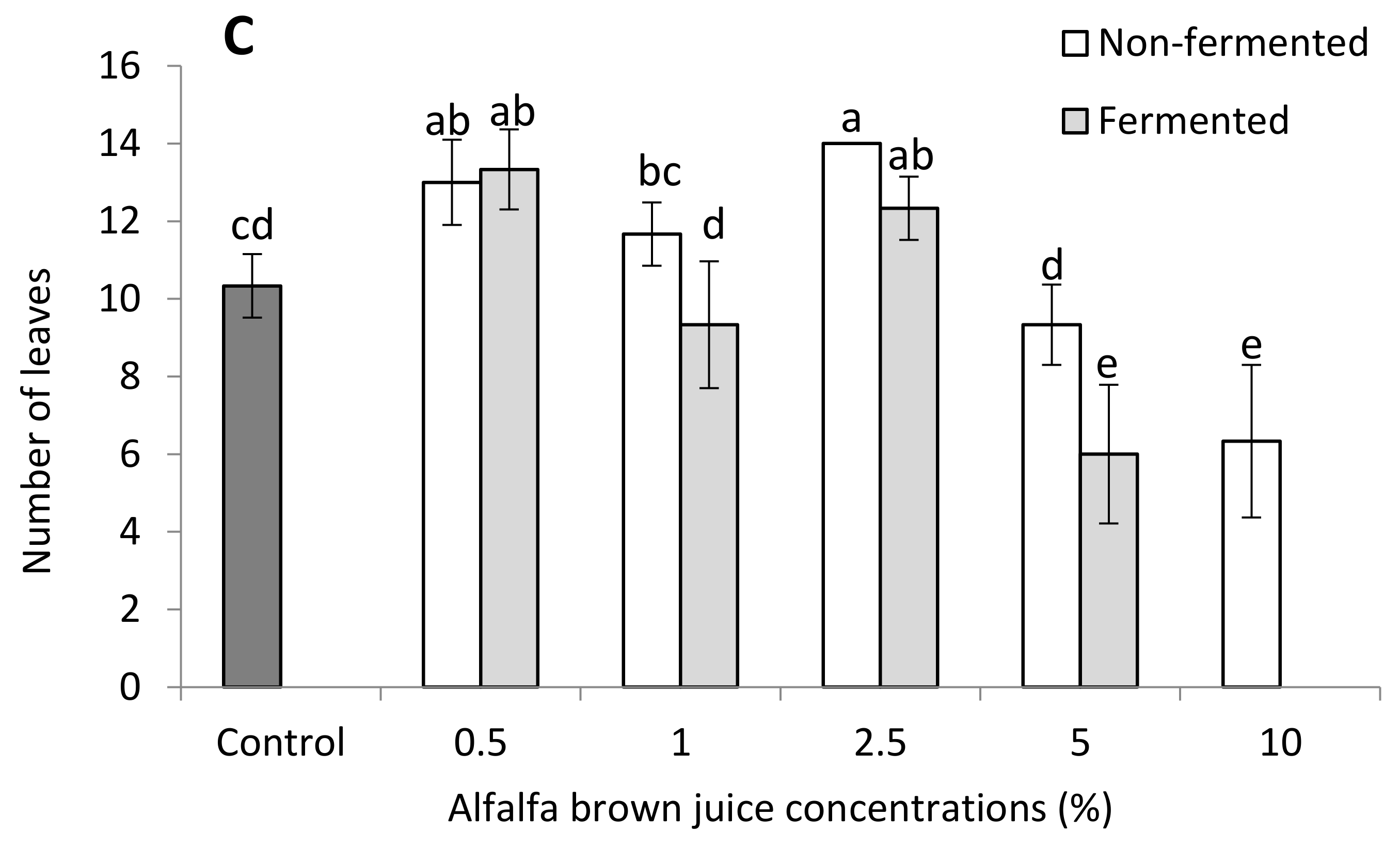
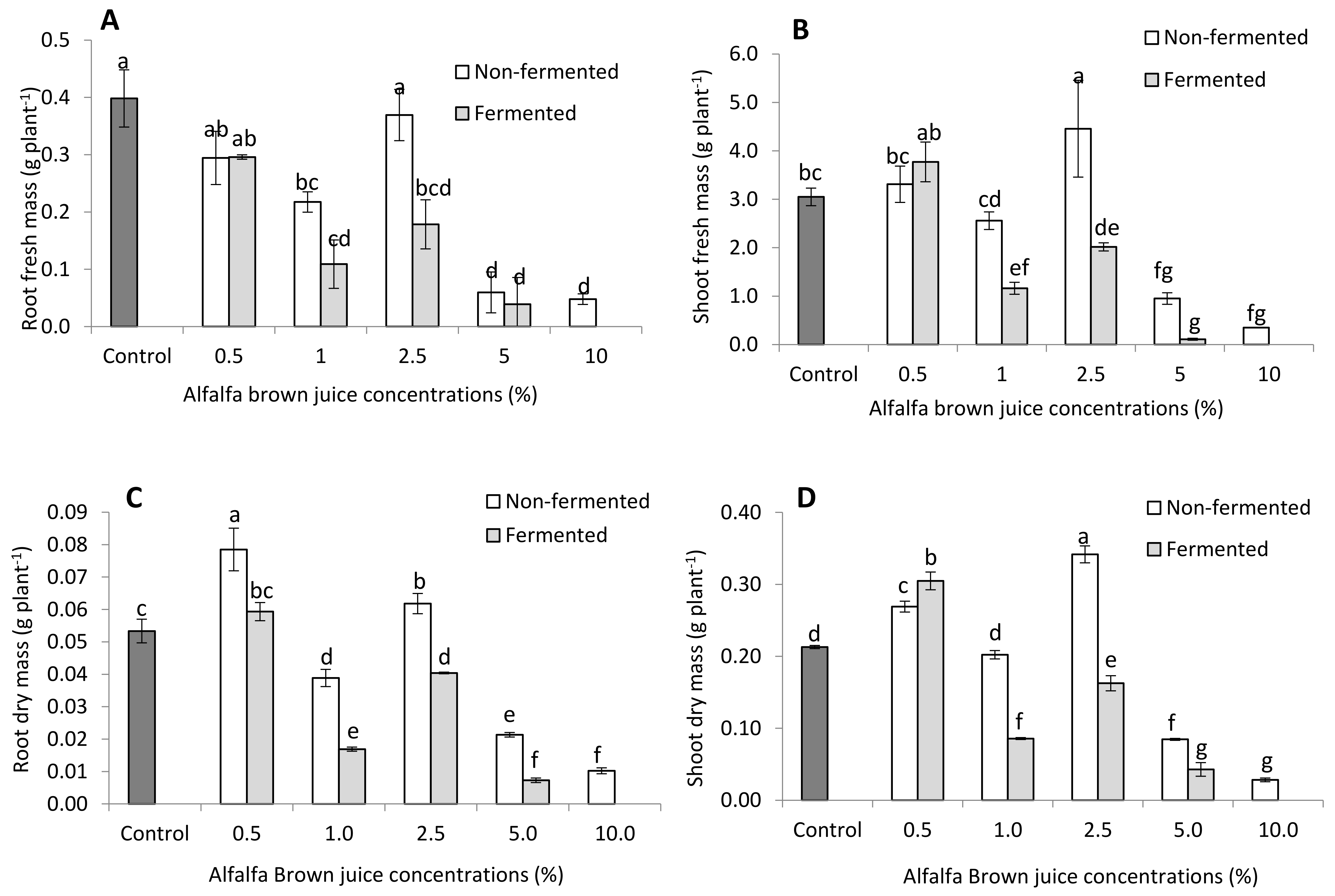
2.2.1. Plant Biometric Parameters
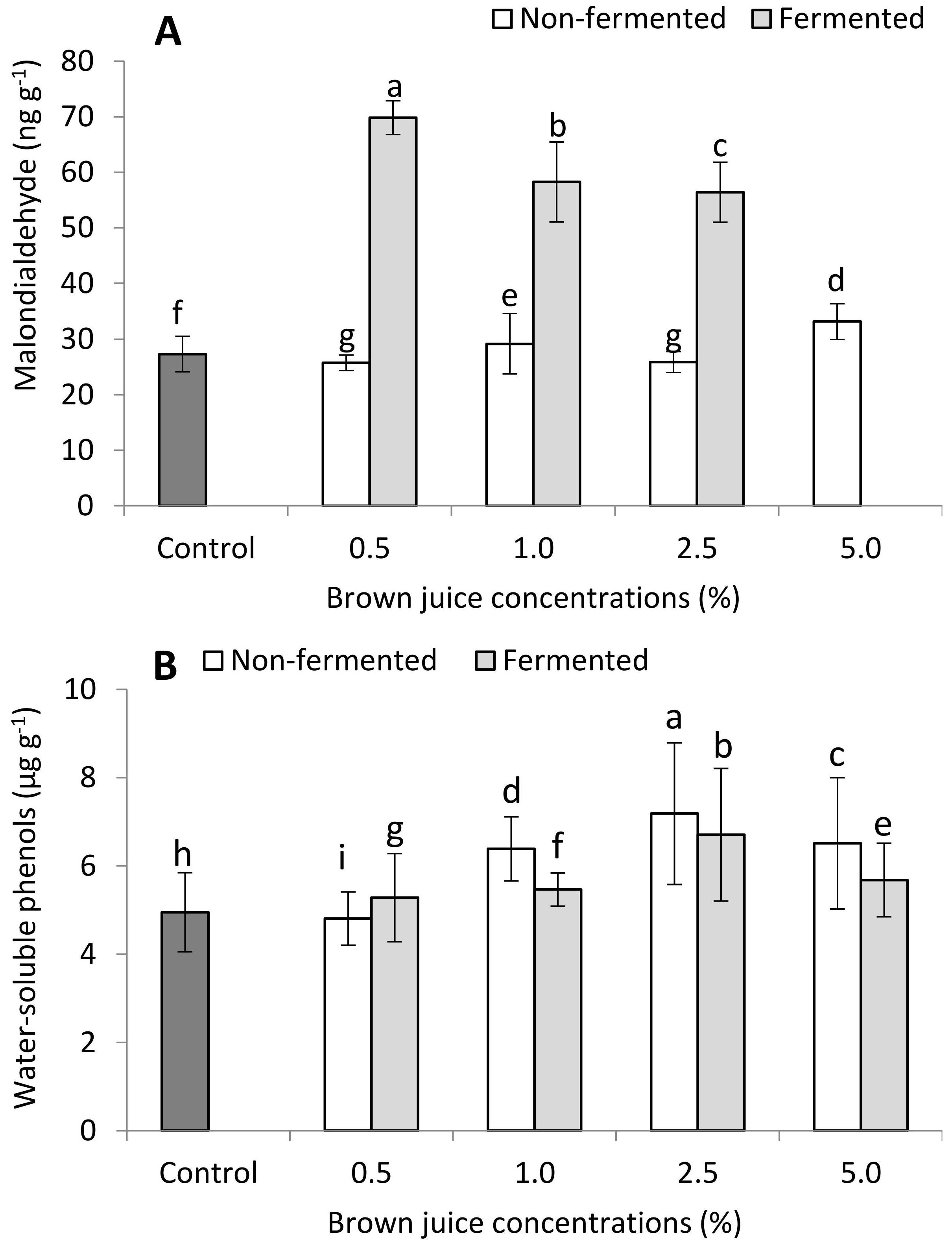
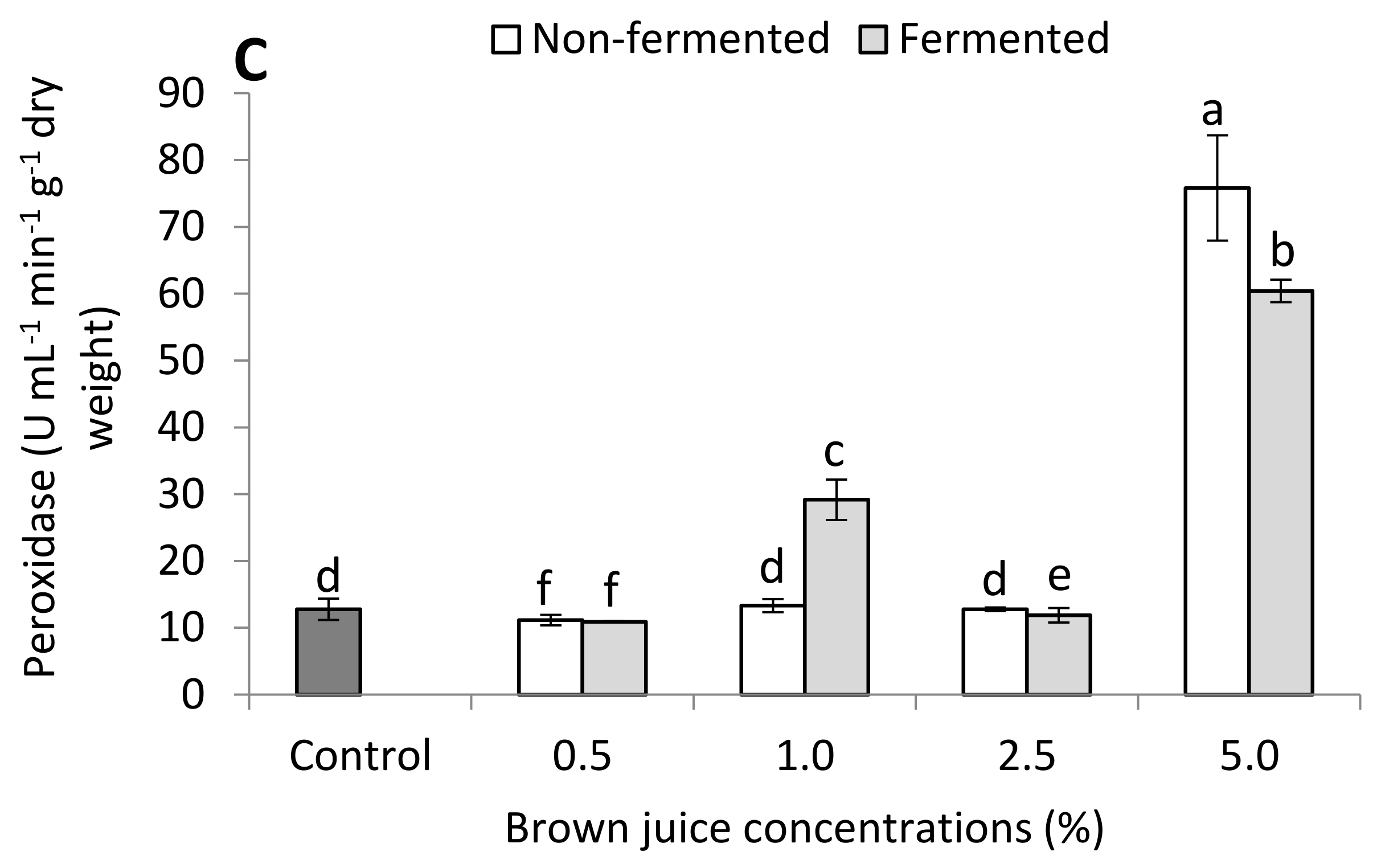
2.2.2. Antioxidant Capacity of French Marigold Plants in the Presence of BJ
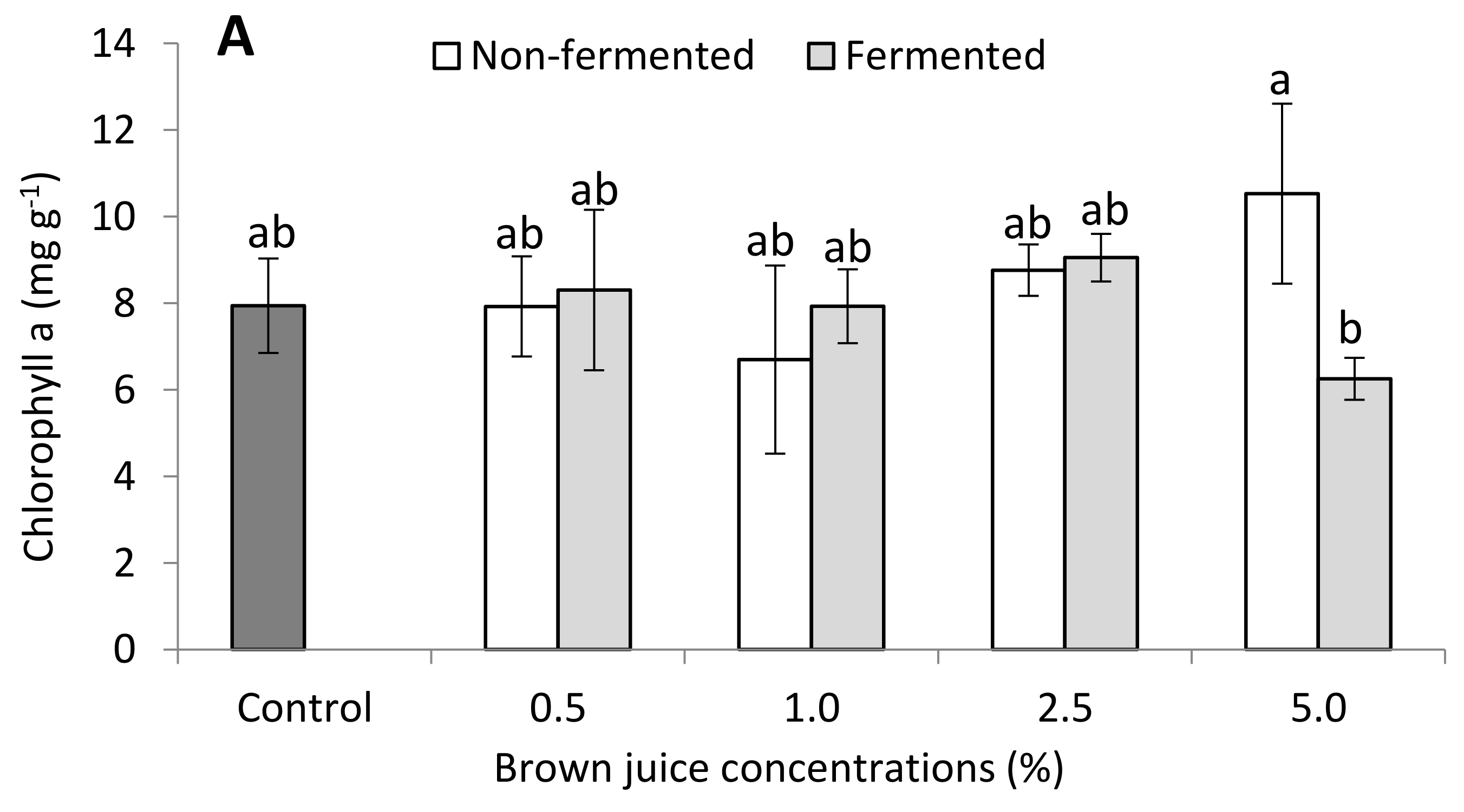
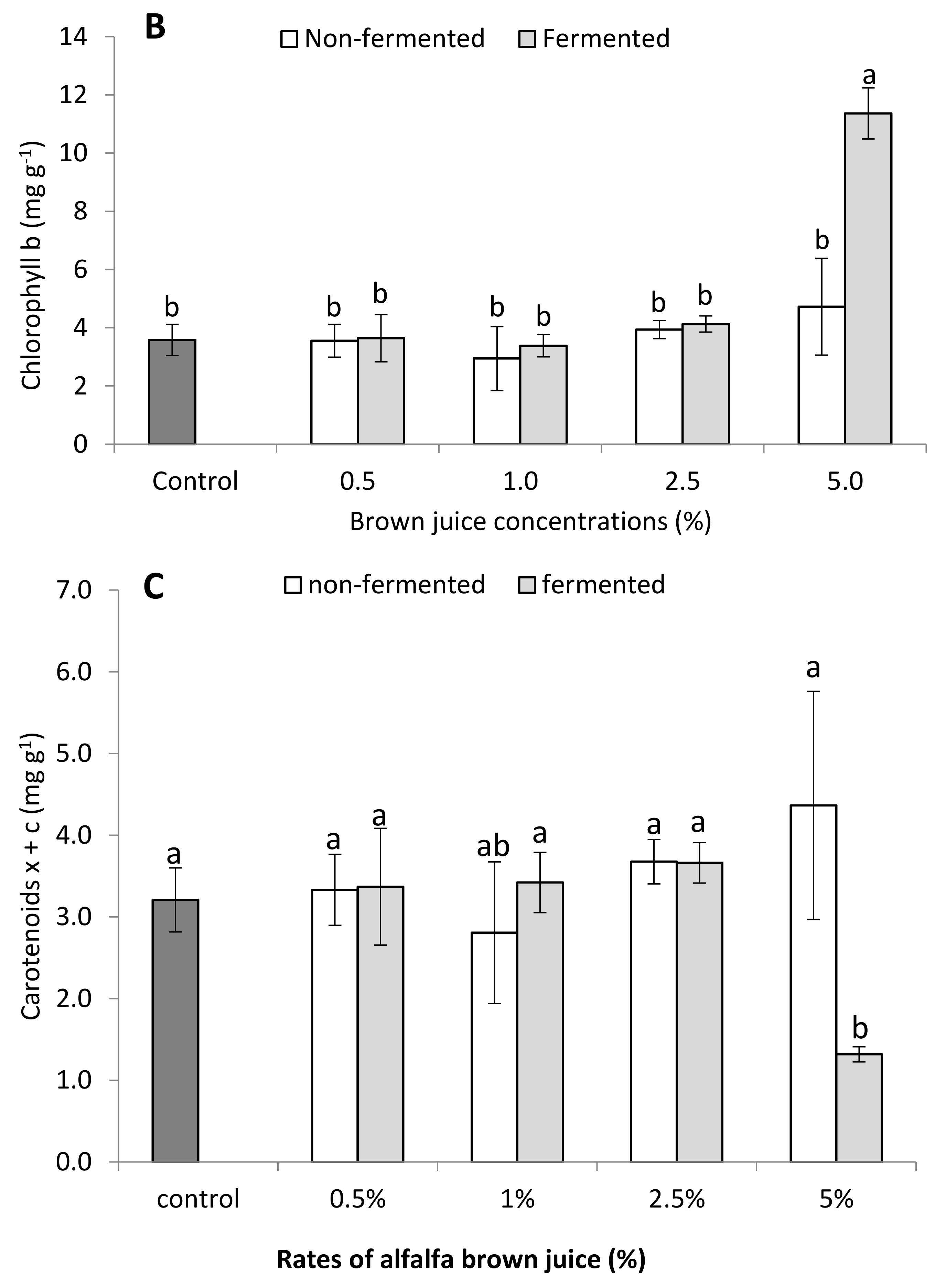
2.2.3. Photosynthetic Pigments of French Marigold Plants in the Presence of BJ
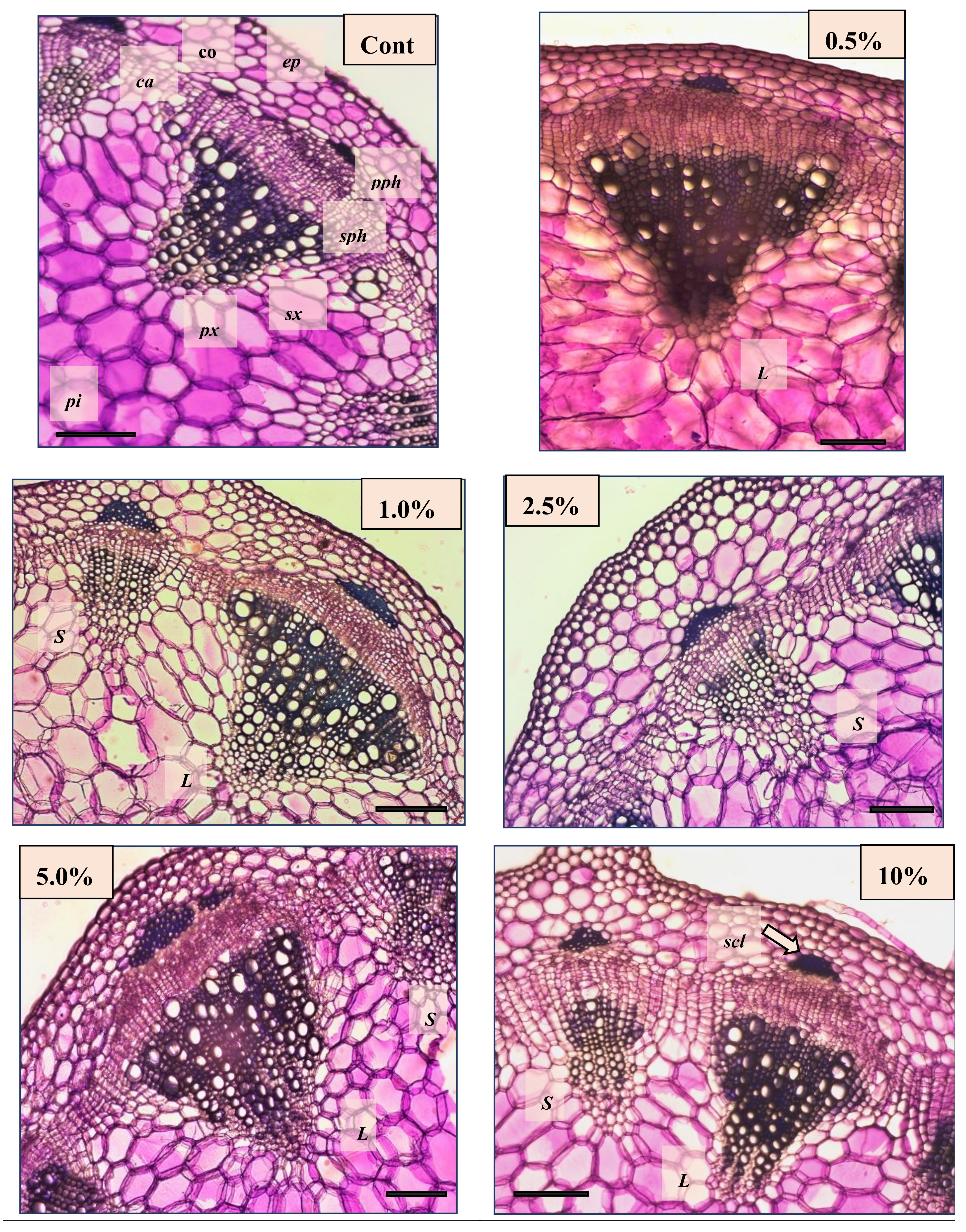
2.3. Effect of Non-Fermented and Fermented BJ on the Anatomical Features of French Marigold
3. Discussion
4. Materials and Methods
4.1. Source of Brown Juice (BJ) and Plant Materials
4.2. Seed Germination Experiment
4.3. Greenhouse Experiment Setup
4.3.1. Determination of Peroxidase
4.3.2. Malondialdehyde and Water-Soluble Phenol Measurement
4.3.3. Photosynthetic Pigment
4.4. Plant Anatomy
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Vita, G.D.; Allegra, V.; Zarbà, A.S. Building Scenarios: A Qualitative Approach to Forecasting Market Developments for Ornamental Plants. Int. J. Bus. Glob. 2015, 15, 130–151. [Google Scholar] [CrossRef]
- Fári, M.G.; Domokos-Szabolcsy, É. Method for Producing Plant Protein Coagulum. Hun. Patent WO2019150144, 8 August 2019. [Google Scholar]
- Bákonyi, N.; Kisvarga, S.; Barna, D.; Tóth, I.O.; El-Ramady, H.R.; El-Sayed, A.G.; Kovács, S.; Fehér, C.; Rozbach, M.; Fári, M.G.; et al. Chemical Traits of Fermented Alfalfa Brown Juice: Its Implications on Physiological, Biochemical, Anatomical, and Growth Parameters of Celosia. Agronomy 2020, 10, 247. [Google Scholar] [CrossRef] [Green Version]
- Manwatkar, W.G.; Gogle, D.P. The Effect of Deproteinised Juice (DPJ) on Seed Germination and Seedling Growth of Different Plants (by Paper Towel Method). Int. J. Life Sci. 2014, 2, 65–68. [Google Scholar]
- Cheynier, V.; Comte, G.; Davies, K.M.; Lattanzio, V.; Martens, S. Plant Phenolics: Recent Advances on Their Biosynthesis, Genetics, and Ecophysiology. Plant Physiol. Biochem. 2013, 72, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Lamont, J.R.; Wilkins, O.; Bywater-Ekegärd, M.; Smith, D.L. From Yogurt to Yield: Potential Applications of Lactic Acid Bacteria in Plant Production. Soil Biol. Biochem. 2017, 111, 1–9. [Google Scholar] [CrossRef]
- Smith, D.L.; Subramanian, S.; Lamont, J.R.; Bywater-Ekegärd, M. Signaling in the Phytomicrobiome: Breadth and Potential. Front. Plant Sci. 2015, 6, 709. [Google Scholar] [CrossRef] [Green Version]
- Kisvarga, S.; Barna, D.; Kovács, S.; Csatári, G.O.; Tóth, I.; Fári, M.G.; Makleit, P.; Veres, S.; Alshaal, T.; Bákonyi, N. Fermented Alfalfa Brown Juice Significantly Stimulates the Growth and Development of Sweet Basil (Ocimum Basilicum L.) Plants. Agronomy 2020, 10, 657. [Google Scholar] [CrossRef]
- Neher, R.T. The Ethnobotany of Tagetes. Econ. Bot. 1968, 22, 317–325. [Google Scholar] [CrossRef]
- Vasudevan, P.; Kashyap, S.; Sharma, S. Tagetes: A Multipurpose Plant. Bioresour. Technol. 1997, 62, 29–35. [Google Scholar] [CrossRef]
- Negi, J.S.; Bisht, V.K.; Bhandari, A.K.; Sundriyal, R.C. Essential Oil Contents and Antioxidant Activity of Tagetes Patula L. J. Essent. Oil Bear. Plants 2013, 16, 364–367. [Google Scholar] [CrossRef]
- Romagnoli, C.; Bruni, R.; Andreotti, E.; Rai, M.K.; Vicentini, C.B.; Mares, D. Chemical Characterization and Antifungal Activity of Essential Oil of Capitula from Wild Indian Tagetes Patula L. Protoplasma 2005, 225, 57–65. [Google Scholar] [CrossRef]
- Comba, L.; Corbet, S.A.; Barron, A.; Bird, A.; Collinge, S.; Miyazaki, N.; Powell, M. Garden Flowers: Insect Visits and the Floral Reward of Horticulturally-Modified Variants. Ann. Bot. 1999, 83, 73–86. [Google Scholar] [CrossRef] [Green Version]
- Rajasekaran, T.; Ravishankar, G.A.; Reddy, B.O. In Vitro Growth of Tagetes Patula L. Hairy Roots, Production of Thiophenes and Its Mosquito Larvicidal Activity. Indian J. Biotechnol. 2004, 3, 92–96. [Google Scholar]
- Dharmagadda, V.S.S.; Naik, S.N.; Mittal, P.K.; Vasudevan, P. Larvicidal Activity of Tagetes Patula Essential Oil against Three Mosquito Species. Bioresour. Technol. 2005, 96, 1235–1240. [Google Scholar] [CrossRef]
- Liu, Y.-T.; Chen, Z.-S.; Hong, C.-Y. Cadmium-Induced Physiological Response and Antioxidant Enzyme Changes in the Novel Cadmium Accumulator, Tagetes Patula. J. Hazard. Mater. 2011, 189, 724–731. [Google Scholar] [CrossRef]
- Bhattacharyya, S.; Datta, S.; Mallick, B.; Dhar, P.; Ghosh, S. Lutein Content and in Vitro Antioxidant Activity of Different Cultivars of Indian Marigold Flower (Tagetes Patula L.) Extracts. J. Agric. Food Chem. 2010, 58, 8259–8264. [Google Scholar] [CrossRef]
- Zhang, P.; Zeng, L.; Su, Y.-X.; Gong, X.-W.; Wang, X.-S. Karyotype Studies on Tagetes Erecta L. and Tagetes Patula L. Afr. J. Biotechnol. 2011, 10, 16138–16144. [Google Scholar]
- Karuppaiah, P.; Krishna, G. Response of Spacings and Nitrogen Levels on Growth, Flowering and Yield Characters of French Marigold (Tagetes Patula Linn). J. Ornam. Hortic. 2005, 8, 96–99. [Google Scholar]
- Chander, S.; Beniwal, B.S.; Dalal, R.P.S.; Sheoran, S. Effect of Organic Manures and Their Levels on Yield, Nutrient Content and Seed Characteristics of French Marigold (Tagetes Patula L.). Ann. Agri-Bio Res. 2015, 20, 207–211. [Google Scholar]
- Draghia, L.; Chelariu, E.-L.; Delinschi, V.; Grădinariu, G.; Bireescu, L. Influence of Bio-Fertilizer Foliar Application on Growth and Anatomical Changes of Tagetes Patula. In Proceedings of the 43rd Croatian and 3rd International Symposium on Agriculture, Opatija, Croatia, 18–21 February 2008; pp. 511–515. [Google Scholar]
- Naik, M.R. Influence of Bio-Fertilizers on Growth, Flowering and Yield of African Marigold Cv. Orange Bunch. Indian Hortic. J. 2015, 5, 78–81. [Google Scholar]
- Tang, S.; Shi, W.; Dong, Y. Organic Amendment Made from Bio-Pharmaceutical Wastes. J. Agro-Environ. Sci. 2006, 25 (Suppl. 1), 234–236. [Google Scholar] [CrossRef]
- Zeljković, S.; Parađiković, N.; Vinković, T.; Tkalec, M. Biostimulant application in the production of seedlings of seasonal flowers. Agrozn. Agro-Knowl. J. 2011, 12, 175–181. [Google Scholar]
- Zeljković, S.; Parađiković, N.; Vinković, T.; Tkalec, M.; Maksimović, I.; Haramija, J. Nutrient status, growth and proline concentration of French marigold (Tagetes patula L.) as affected by biostimulant treatment. J. Food Agric. Environ. 2013, 11, 2324–2327. [Google Scholar]
- Zeljković, S.; Parađiković, N.; Vinković, T.; Oljača, R.; Tkalec, M. Contents of mineral elements in nursery stock of marigold (Tagetes patula L.) under bio stimulant treatment. Agrozn. Agro-Knowl. J. 2010, 11, 127–134. [Google Scholar]
- Parađiković, N.; Teklić, T.; Zeljković, S.; Lisjak, M.; Špoljarević, M. Biostimulants Research in Some Horticultural Plant Species—A Review. Food Energy Secur. 2019, 8, e00162. [Google Scholar] [CrossRef]
- Majkowska-Gadomska, J.; Francke, A.; Dobrowolski, A.; Mikulewicz, E. The Effect of Selected Biostimulants on Seed Germination of Four Plant Species. Acta Agrophys. 2017, 24, 591–599. [Google Scholar]
- Turner, G.D.; Lau, R.R.; Young, D.R. Effect of Acidity on Germination and Seedling Growth of Paulownia Tomentosa. J. Appl. Ecol. 1988, 25, 561–567. [Google Scholar] [CrossRef]
- Fatima, U.; Senthil-Kumar, M. Plant and Pathogen Nutrient Acquisition Strategies. Front. Plant Sci. 2015, 6, 750. [Google Scholar] [CrossRef] [PubMed]
- Moghaddam, M.; Farhadi, N.; Panjtandoust, M.; Ghanati, F. Seed Germination, Antioxidant Enzymes Activity and Proline Content in Medicinal Plant Tagetes Minuta under Salinity Stress. Plant Biosyst. 2019, 154, 835–842. [Google Scholar] [CrossRef]
- Reddy, G.U.; Deshmukh, V.R.; Joshi, R.N.; Kayama, R. Utilization of Alfalfa (Mediacago sativa L.) Whey as a Fertilizer in Irrigation. Jpn. J. Grassl. Sci. 1987, 33, 32–37. [Google Scholar] [CrossRef]
- Ream, H.W.; Smith, D.; Walgenbach, R.P. Effects of Deproteinized Alfalfa Juice Applied to Alfalfa Bromegrass, Bromegrass, and Corn. Agronom. J. 1977, 69, 685–689. [Google Scholar] [CrossRef]
- Pirie, N.W. Leaf Protein and Other Aspects of Fodder Fractionation; Cambridge University Press: Cambridge, UK, 1987; p. 209. [Google Scholar]
- Demmig-Adams, B.; Adams, W.W. Antioxidants in Photosynthesis and Human Nutrition. Science 2002, 298, 2149–2153. [Google Scholar] [CrossRef] [PubMed]
- Yasheshwar, U.S.; Sharma, M.P.; Khan, W.; Ahmad, S. Variation in Ornamental Traits, Physiological Responses of Tagetes Erecta L. and T. Patula L. in Relation to Antioxidant and Metabolic Profile under Deficit Irrigation Strategies. Sci. Hortic. 2017, 214, 200–208. [Google Scholar] [CrossRef]
- Sathiyaraj, G.; Lee, O.R.; Parvin, S.; Khorolragchaa, A.; Kim, Y.-J.; Yang, D.C. Transcript Profiling of Antioxidant Genes during Biotic and Abiotic Stresses in Panax Ginseng CA Meyer. Mol. Biol. Rep. 2011, 38, 2761–2769. [Google Scholar] [CrossRef]
- Anugoolprasert, O.; Kinoshita, S.; Naito, H.; Shimizu, M.; Ehara, H. Effect of Low PH on the Growth, Physiological Characteristics and Nutrient Absorption of Sago Palm in a Hydroponic System. Plant Prod. Sci. 2012, 15, 125–131. [Google Scholar] [CrossRef]
- Long, A.; Zhang, J.; Yang, L.T.; Ye, X.; Lai, N.W.; Tan, L.L.; Lin, D.; Chen, L.S. Effects of Low PH on Photosynthesis, Related Physiological Parameters, and Nutrient Profiles of Citrus. Front. Plant Sci. 2017, 8, 185. [Google Scholar] [CrossRef] [Green Version]
- Alsaeedi, A.; El-Ramady, H.; Alshaal, T.; El-Garawani, M.; Elhawat, N.; Al-Otaibi, A. Exogenous Nanosilica Improves Germination and Growth of Cucumber by Maintaining K+/Na+ Ratio under Elevated Na+ Stress. Plant Physiol. Biochem. 2018, 125, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Abdul-Baki, A.A.; Anderson, J.D. Vigor Determination in Soybean Seed by Multiple Criteria. Crop Sci. 1973, 13, 630–633. [Google Scholar] [CrossRef]
- Roxas, V.P.; Smith, R.K.; Allen, E.R.; Allen, R.D. Overexpression of Glutathione S-Transferase/Glutathione Peroxidase Enhances the Growth of Transgenic Tobacco Seedlings during Stress. Nat. Biotechnol. 1997, 15, 988–991. [Google Scholar] [CrossRef]
- Zhang, Z.; Huang, R. Analysis of Malondialdehyde, Chlorophyll Proline, Soluble Sugar, and Glutathione Content in Arabidopsis Seedling. Bio-Protocol 2013, 3. [Google Scholar] [CrossRef]
- Box, J.D. Investigation of the Folin-Ciocalteau Phenol Reagent for the Determination of Polyphenolic Substances in Natural Waters. Water Res. 1983, 17, 511–525. [Google Scholar] [CrossRef]
- Moran, R.; Porath, D. Chlorophyll Determination in Intact Tissues Using N,N-Dimethylformamide. Plant Physiol. 1980, 65, 478–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wellburn, A.R. The Spectral Determination of Chlorophylls a and b, as Well as Total Carotenoids, Using Various Solvents with Spectrophotometers of Different Resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Holm, S. A Simple Sequentially Rejective Multiple Test Procedure. Scand. J. Stat. 1979, 6, 65–70. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diameter (Stem) | Large Vascular Bundles | Small Vascular Bundles | |||||
|---|---|---|---|---|---|---|---|
| Diameter (Bundle) | Secondary Xylem | Sclerenchyma | Diameter (Bundle) | Secondary Xylem | SCLERENCHYMA | ||
| Cont | 3172.64 ± 109.84c | 539.11 ± 63.36c | 191.33 ± 44.91d | 26.97 ± 8.64c | 327.56 ± 40.57d | 189.73 ± 38.57d | 34.42 ± 9.02c |
| 0.5% | 3537.60 ± 235.97b | 603.41 ± 84.27b | 274.23 ± 67.53b | 44.08 ± 11.42b | 414.69 ± 66.45ab | 271.86 ± 62.56b | 56.53 ± 17.06b |
| 1% | 3669.29 ± 257.50ab | 621.45 ± 32.82b | 288.66 ± 41.93b | 42.05 ± 12.51b | 347.24 ± 34.27bd | 256.10 ± 42.50b | 54.80 ± 12.54b |
| 2.5% | 3751.34 ± 174.37a | 660.55 ± 59.15a | 317.23 ± 60.16a | 39.86 ± 11.41b | 392.04 ± 44.40b | 277.75 ± 45.62b | 50.81 ± 14.04b |
| 5% | 3802.14 ± 141.03a | 676.12 ± 63.15a | 285.16 ± 49.41b | 54.61 ± 14.83a | 431.06 ± 71.77a | 312.08 ± 66.55a | 60.27 ± 13.25a |
| 10% | 3504.17 ± 186.73b | 610.87 ± 75.64b | 240.98 ± 48.65c | 45.13 ± 11.50b | 361.14 ± 50.91c | 238.52 ± 47.36c | 48.85 ± 11.54b |
| BJ Rates (%) | pH | EC (dS m−1) | Absorbed Solution (g 10−1 seeds 2 h−1) | |||
|---|---|---|---|---|---|---|
| Non-Fermented | Fermented | Non-Fermented | Fermented | Non-Fermented | Fermented | |
| Distilled water | 6.91 | 0.0 | 0.032 | |||
| 0.5 | 5.68 | 5.51 | 0.19 | 0.16 | 0.031 | 0.028 |
| 1.0 | 5.61 | 5.39 | 0.35 | 0.28 | 0.029 | 0.030 |
| 2.5 | 5.43 | 5.28 | 0.81 | 0.64 | 0.025 | 0.027 |
| 5.0 | 5.33 | 5.19 | 1.43 | 1.20 | 0.024 | 0.032 |
| 10.0 | 5.24 | 5.09 | 2.54 | 2.11 | 0.026 | 0.032 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barna, D.; Kisvarga, S.; Kovács, S.; Csatári, G.; Tóth, I.O.; Fári, M.G.; Alshaal, T.; Bákonyi, N. Raw and Fermented Alfalfa Brown Juice Induces Changes in the Germination and Development of French Marigold (Tagetes patula L.) Plants. Plants 2021, 10, 1076. https://doi.org/10.3390/plants10061076
Barna D, Kisvarga S, Kovács S, Csatári G, Tóth IO, Fári MG, Alshaal T, Bákonyi N. Raw and Fermented Alfalfa Brown Juice Induces Changes in the Germination and Development of French Marigold (Tagetes patula L.) Plants. Plants. 2021; 10(6):1076. https://doi.org/10.3390/plants10061076
Chicago/Turabian StyleBarna, Döme, Szilvia Kisvarga, Szilvia Kovács, Gábor Csatári, Ibolya O. Tóth, Miklós Gábor Fári, Tarek Alshaal, and Nóra Bákonyi. 2021. "Raw and Fermented Alfalfa Brown Juice Induces Changes in the Germination and Development of French Marigold (Tagetes patula L.) Plants" Plants 10, no. 6: 1076. https://doi.org/10.3390/plants10061076