
Chemical Variability of Peel and Leaf Essential Oils in the Citrus Subgenus Papeda (Swingle) and Few Relatives

 , , and
, , and
Abstract
:1. Introduction
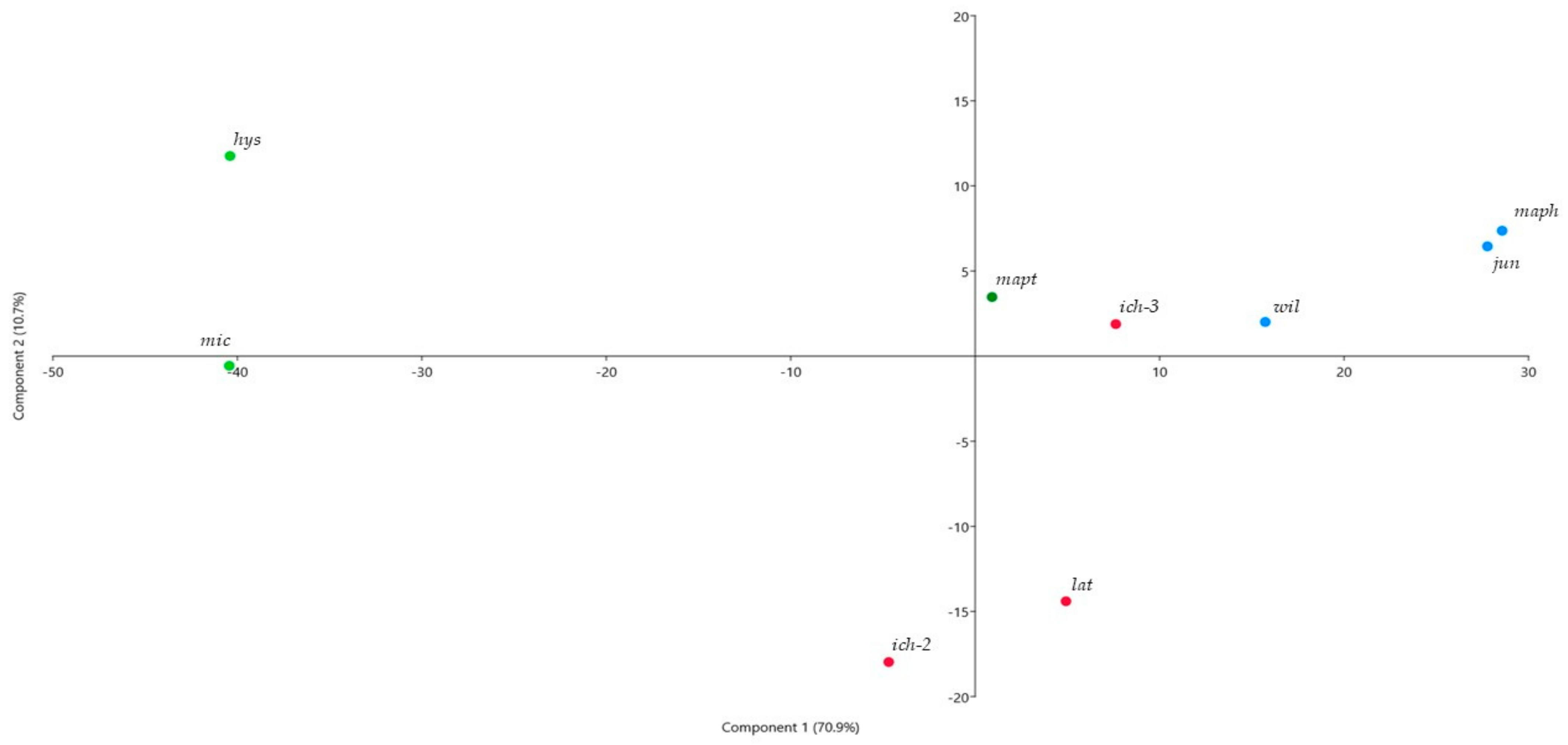
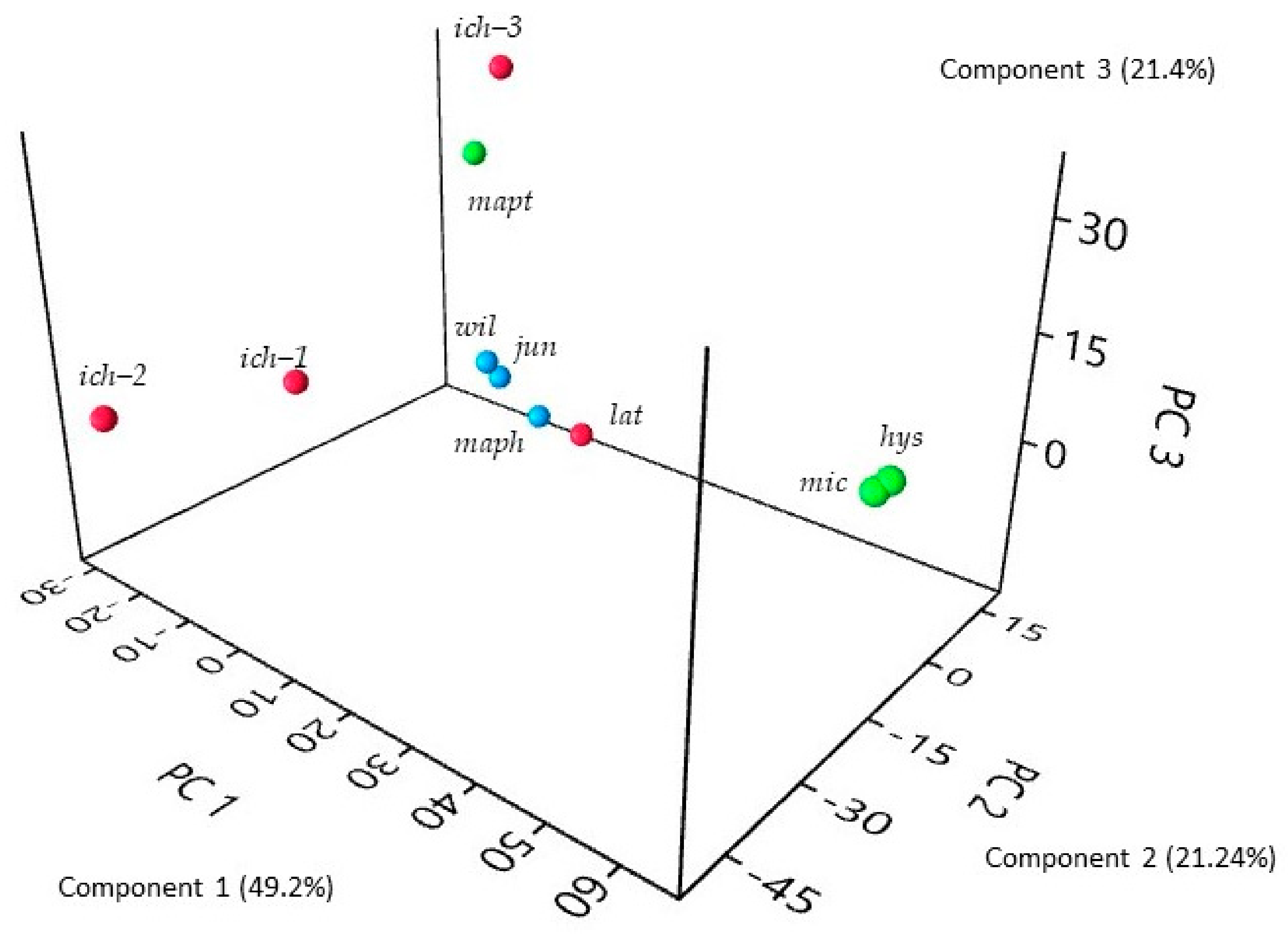
2. Results and Discussion
2.1. Peel Oils
2.1.1. Section Papeda
2.1.2. Section Papedocitrus
2.1.3. Related Species
2.2. Leaf Oils
2.2.1. Section Papeda
Combava (C. hystrix DC.) and Biasong (C. micrantha Wester)
Melanesian Papeda (C. macroptera Montr.)
2.2.2. Section Papedocitrus
Ichang Papeda (C. ichangensis Swingle)
Khasi papeda (C. latipes (Swingle) Tanaka)
2.2.3. Related Species
Ichang Lemon (C. wilsonii Tanaka)
Yuzu (C. junos Sieb. Ex Tan)
Alemow (C. macrophylla Wester)
3. Materials and Methods
3.1. Plant Material
3.2. Gas Chromatography (GC) Analysis
3.3. Mass Spectrometry
3.4. NMR Analysis
3.5. Identification of Individual Components
3.6. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wu, G.A.; Terol, J.; Ibanez, V.; López-García, A.; Pérez-Román, E.; Borredá, C.; Domingo, C.; Tadeo, F.R.; Carbonell-Caballero, J.; Alonso, R. Genomics of the origin and evolution of citrus. Nature 2018, 554, 311–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ollitrault, P.; Jacquemond, C.; Dubois, C.; Luro, F. Citrus. In Genetic Diversity of Cultivated Tropical Plants; Hamon, P., Seguin, M., Perrier, X., Glaszmann, J.-C., Eds.; CIRAD: Montpellier, France, 2003; pp. 193–217. [Google Scholar]
- Nicolosi, E. Origin and taxonomy. In Citrus Genetics, Breeding and Biotechnology; CAB International Publishers: Wallingford, UK, 2007; pp. 19–43. [Google Scholar]
- Garcia-Lor, A.; Curk, F.; Snoussi-Trifa, H.; Morillon, R.; Ancillo, G.; Luro, F.; Navarro, L.; Ollitrault, P. A nuclear phylogenetic analysis: SNPs, indels and SSRs deliver new insights into the relationships in the ‘True Citrus Fruit Trees’ group (Citrinae, Rutaceae) and the origin of cultivated species. Ann. Bot. 2013, 111, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, G.A.; Prochnik, S.; Jenkins, J.; Salse, J.; Hellsten, U.; Murat, F.; Perrier, X.; Ruiz, M.; Scalabrin, S.; Terol, J. Sequencing of diverse mandarin, pummelo and orange genomes reveals complex history of admixture during citrus domestication. Nat. Biotechnol. 2014, 32, 656–662. [Google Scholar] [CrossRef]
- Swingle, W.T.; Reece, P.C. The Botany of Citrus and Its Wild Relatives. In The Citrus Industry; Reuther, W., Webber, H.J., Batchelor, L.D., Eds.; University of California: Berkeley, LA, USA, 1967; Volume 1, pp. 190–430. [Google Scholar]
- Demarcq, B.; Cavailles, M.; Lambert, L.; Schippa, C.; Ollitrault, P.; Luro, F. Characterization of odor-active compounds of ichang lemon (Citrus wilsonii Tan.) and identification of its genetic interspecific origin by DNA genotyping. J. Agric. Food Chem. 2021, 69, 3175–3188. [Google Scholar] [CrossRef] [PubMed]
- Dugo, G.; Mondello, L. Citrus Oils: Composition, Advanced Analytical Techniques, Contaminants, and Biological Activity; Dugo, G., Mondello, L., Eds.; CRC Press: Boca Raton, FL, USA, 2010; 561p. [Google Scholar]
- Swingle, W.T. The Botany of Citrus and Its Wild Relatives in the Orange Subfamily. In The Citrus Industry; University of California: Berkeley, LA, USA, 1943; Volume 1, pp. 128–474. [Google Scholar]
- Talon, M.; Caruso, M.; Gmitter, F.G., Jr. The Genus Citrus; Woodhead Publishing: Duxford, UK, 2020; 515p. [Google Scholar]
- Ollitrault, P.; Terol, J.; Garcia-Lor, A.; Bérard, A.; Chauveau, A.; Froelicher, Y.; Belzile, C.; Morillon, R.; Navarro, L.; Brunel, D. SNP mining in C. clementina BAC end sequences; transferability in the citrus genus (Rutaceae), phylogenetic inferences and perspectives for genetic mapping. BMC Genom. 2012, 13, 13. [Google Scholar] [CrossRef] [Green Version]
- Curk, F.; Ollitrault, F.; Garcia-Lor, A.; Luro, F.; Navarro, L.; Ollitrault, P. Phylogenetic origin of limes and lemons revealed by cytoplasmic and nuclear markers. Ann. Bot. 2016, 117, 565–583. [Google Scholar] [CrossRef] [Green Version]
- Ollitrault, P.; Curk, F.; Krueger, R. Citrus taxonomy. In The Genus Citrus; Elsevier: Amsterdam, The Netherlands, 2020; pp. 57–81. [Google Scholar]
- Tanaka, T. Contribution to the knowledge of citrus classification. Rep. Citrol. 1961, 3, 107–114. [Google Scholar]
- Zhang, D.X.; Mabberley, D.J. Citrus Linnaeus, Sp. Pl. 2: 782. 1753. In Flora of China 11 (Oxalidaceae through Aceraceae); Wu, Z.Y., Raven, P.H., Hong, D.Y., Eds.; Science Press: Beijing, China; Missouri Botanical Garden Press: St Louis, MI, USA, 2008; pp. 90–96. [Google Scholar]
- Malik, S.K.; Uchoi, A.; Kumar, S.; Choudhary, R.; Pal, D.; Kole, P.R.; Chaudhury, R.; Bhat, K.V. Molecular characterization of Citrus macroptera Montr. (Satkara): An endangered wild species from Northeast India. Plant Biosyst. 2013, 147, 857–863. [Google Scholar] [CrossRef]
- Yang, X.; Li, H.; Yu, H.; Chai, L.; Xu, Q.; Deng, X. Molecular phylogeography and population evolution analysis of Citrus ichangensis (Rutaceae). Tree Genet. Genomes 2017, 13, 29. [Google Scholar] [CrossRef]
- Uehara, A.; Baldovini, N. Volatile constituents of Yuzu (Citrus junos Sieb. Ex Tanaka) peel oil: A review. Flavour Fragr. J. 2021, 36, 292–318. [Google Scholar] [CrossRef]
- Huang, Y.; Pu, Z.; Chen, Q. The chemical composition of the leaf essential oils from 110 citrus species, cultivars, hybrids and varieties of chinese origin. Perfum. Flavorist 2000, 25, 53–66. [Google Scholar]
- Liu, C.; Jiang, D.; Cheng, Y.; Deng, X.; Chen, F.; Fang, L.; Ma, Z.; Xu, J. Chemotaxonomic study of citrus, poncirus and fortunella genotypes based on peel oil volatile compounds-deciphering the genetic origin of mangshanyegan (Citrus nobilis Lauriro). PLoS ONE 2013, 8, e58411. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Xie, Y.; Liu, C.; Chen, S.; Hu, S.; Xie, Z.; Deng, X.; Xu, J. Comprehensive comparative analysis of volatile compounds in citrus fruits of different species. Food Chem. 2017, 230, 316–326. [Google Scholar] [CrossRef]
- Zhang, H.; Chen, M.; Wen, H.; Wang, Z.; Chen, J.; Fang, L.; Zhang, H.; Xie, Z.; Jiang, D.; Cheng, Y. Transcriptomic and metabolomic analyses provide insight into the volatile compounds of citrus leaves and flowers. BMC Plant Biol. 2020, 20, 1–14. [Google Scholar] [CrossRef]
- Dugo, G.; Di Giacomo, A. Citrus: The Genus Citrus; CRC Press: Boca Raton, FL, USA, 2002; 642p. [Google Scholar]
- Waikedre, J.; Dugay, A.; Barrachina, I.; Herrenknecht, C.; Cabalion, P.; Fournet, A. Chemical composition and antimicrobial activity of the essential oils from new caledonian Citrus macroptera and Citrus hystrix. Chem. Biodivers. 2010, 7, 871–877. [Google Scholar] [CrossRef] [PubMed]
- Hijaz, F.; Nehela, Y.; Killiny, N. Possible role of plant volatiles in tolerance against huanglongbing in citrus. Plant Signal. Behav. 2016, 11, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Ollitrault, P.; Luro, F. Citrus. In Tropical Plant Breeding; Charrier, A., Jacquot, M., Hamon, S., Nicolas, D., Eds.; CIRAD: Enfield, NH, USA; Plymouth, UK, 2001; pp. 55–77. ISBN 9782876144262. [Google Scholar]
- Xi, W.; Li, L.; Jiang, D.; Jiao, B.; Zhou, Z. Variation patterns of the volatile compounds in flowers of chinese native citrus species and their taxonomic implications. J. Food Nutr. Res. 2015, 3, 235–245. [Google Scholar] [CrossRef]
- Jing, L.; Lei, Z.; Zhang, G.; Pilon, A.C.; Huhman, D.V.; Xie, R.; Xi, W.; Zhou, Z.; Sumner, L.W. Metabolite profiles of essential oils in citrus peels and their taxonomic implications. Metabolomics 2015, 11, 952–963. [Google Scholar] [CrossRef]
- Zhang, H.; Wen, H.; Chen, J.; Peng, Z.; Shi, M.; Chen, M.; Yuan, Z.; Liu, Y.; Zhang, H.; Xu, J. Volatile compounds in fruit peels as novel biomarkers for the identification of four citrus species. Molecules 2019, 24, 4550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luro, F.; Venturini, N.; Costantino, G.; Paolini, J.; Ollitrault, P.; Costa, J. Genetic and Chemical diversity of citron (Citrus medica L.) based on nuclear and cytoplasmic markers and leaf essential oil composition. Phytochemistry 2012, 77, 186–196. [Google Scholar] [CrossRef]
- Luro, F.; Bloquel, E.; Tomu, B.; Costantino, G.; Tur, I.; Riolacci, S.; Varamo, F.; Ollitrault, P.; Froelicher, Y.; Curk, F.; et al. The INRA-CIRAD citrus germplasm collection of San Giuliano, Corsica. In AGRUMED: Archaeology and History of Citrus Fruit in the Mediterranean: Acclimatization, Diversifications, Uses; Fiorentino, G., Zech-Matterne, V., Eds.; Collection du Centre Jean Bérard, Publications du Centre Jean Bérard: Naples, Italy, 2017; pp. 243–261. ISBN 9782918887775. [Google Scholar]
- Lawrence, B.M. The oil composition of less common Citrus species. In Citrus: The Genus Citrus; CRC Press: Boca Raton, FL, USA, 2002; pp. 318–354. [Google Scholar]
- Rana, V.S.; Blazquez, M.A. Compositions of the volatile oils of Citrus macroptera and C. maxima. Nat. Prod. Commun. 2012, 7, 1371–1372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miah, M.N.; Bachar, S.C.; Nahar, L.; Rahman, M.S.; Rashid, M.A.; Hadiuzzaman, S.; Sarker, S.D. Composition of the volatiles of Citrus macroptera var. annamensis and evaluation of bioactivity. J. Essent. Oil Bear. Plants 2010, 13, 211–218. [Google Scholar] [CrossRef]
- Lota, M.-L.; de Rocca Serra, D.; Tomi, F.; Casanova, J. Chemical variability of peel and leaf essential oils of mandarins from Citrus reticulata Blanco. Biochem. Syst. Ecol. 2000, 28, 61–78. [Google Scholar] [CrossRef]
- Venturini, N.; Curk, F.; Desjobert, J.-M.; Karp, D.; Costa, J.; Paolini, J. Chemotaxonomic Investigations of peel and petitgrain essential oils from 17 citron cultivars. Chem. Biodivers. 2010, 7, 736–751. [Google Scholar] [CrossRef]
- Lota, M.-L.; de Rocca Serra, D.; Tomi, F.; Jacquemond, C.; Casanova, J. Volatile components of peel and leaf oils of lemon and lime species. J. Agric. Food Chem. 2002, 50, 796–805. [Google Scholar] [CrossRef] [PubMed]
- Tomi, F.; Barzalona, M.; Casanova, J.; Luro, F. Chemical variability of the leaf oil of 113 hybrids from Citrus clementina (Commun) × Citrus deliciosa (Willow Leaf). Flavour Fragr. J. 2008, 23, 152–163. [Google Scholar] [CrossRef]
- Agouillal, F.; Taher, Z.M.; Moghrani, H.; Nasrallah, N.; El Enshasy, H. A Review of genetic taxonomy, biomolecules chemistry and bioactivities of Citrus hystrix DC. Biosci. Biotechnol. Res. Asia 2017, 14, 285. [Google Scholar] [CrossRef]
- Nicolosi, E.; Deng, Z.N.; Gentile, A.; La Malfa, S.; Continella, G.; Tribulato, E. Citrus phylogeny and genetic origin of important species as investigated by molecular markers. Theor. Appl. Genet. 2000, 100, 1155–1166. [Google Scholar] [CrossRef]
- Penjor, T.; Yamamoto, M.; Uehara, M.; Ide, M.; Matsumoto, N.; Matsumoto, R.; Nagano, Y. Phylogenetic relationships of citrus and its relatives based on MatK gene sequences. PLoS ONE 2013, 8, e62574. [Google Scholar]
- Sutour, S.; Bradesi, P.; Luro, F.; Casanova, J.; Tomi, F. Germacra-1 (10), 5-dien-4α-ol in Fortunella sp. leaf oils. Flavour Fragr. J. 2015, 30, 445–450. [Google Scholar] [CrossRef]
- Kasali, A.A.; Olaniyan, A.A. Citrus essential oil of Nigeria Part III volatile constituents of Citrus jambhiri lush leaf oil. J. Essent. Oil Bear. Plants 2009, 12, 690–693. [Google Scholar] [CrossRef]
- Chen, H.P.; Yang, K.; You, C.X.; Du, S.S.; Cai, Q.; He, Q.; Geng, Z.F.; Deng, Z.W. Chemical constituents and biological activities against Tribolium castaneum (Herbst) of the essential oil from Citrus wilsonii leaves. J. Serb. Chem. Soc. 2014, 79, 1213–1222. [Google Scholar] [CrossRef]
- Kamiyama, S. Studies on leaf oils of citrus species: Part IV composition of leaf oils from Funadoko-Mikan, Sanbokan, Kawabata-Mikan, Shiikuwasha, Yuzu and Otaheite-Orange. Agric. Biol. Chem. 1970, 34, 1561–1568. [Google Scholar] [CrossRef] [Green Version]
- Lota, M.-L.; de Rocca Serra, D.; Tomi, F.; Casanova, J. Chemical variability of peel and leaf essential oils of 15 species of mandarins. Biochem. Syst. Ecol. 2001, 29, 77–104. [Google Scholar] [CrossRef]
- Tomi, F.; Bradesi, P.; Bighelli, A.; Casanova, J. Computer-aided identification of individual components of essential oils using Carbon-13 NMR spectroscopy. J. Magn. Reson. Anal 1995, 1, 25–34. [Google Scholar]
- McLafferty, F.W.; Stauffer, D.B. The Wiley/NBS Registry of Mass Spectral Data; Wiley: New York, NY, USA, 1989; Volume 1. [Google Scholar]
- König, W.A.; Hochmuth, D.H.; Joulain, D. Terpenoids and Related Constituents of Essential Oils. Library of Mass Finder 2.1; Institute of Organic Chemistry: Hamburg, Germany, 2001. [Google Scholar]
- McLafferty, F.W.; Stauffer, D.B. Wiley Registry of Mass Spectral Data, Mass Spectrometry Library Search System Bench-Top/PBM, Version 3.10 d; Wiley-Interscience: Palisade Newfield, NJ, USA, 1994. [Google Scholar]
- Joulain, D.; König, W.A. The Atlas of Spectral Data of Sesquiterpene Hydrocarbons; EB-Verlag: Hamburg, Germany, 1998; 658p. [Google Scholar]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectrometry; Allured Publishing Corporation: Carol Stream, IL, USA, 2007; Volume 456, 698p. [Google Scholar]
- Tomi, F.; Casanova, J. 13C-NMR as a Tool for Identification of Individual Components of Essential Oils from Labiate—A Review. In Proceedings of the I International Symposium on the Labiatae: Advances in Production, Biotechnology and Utilisation, Sanremo, Italy, 22–25 February 2006; Volume 723, pp. 185–192. [Google Scholar]
- Bighelli, A.; Casanova, J. Analytical tools for analyzing Cymbopogon oils. In Essential Oil Bearing Grasses—The Genus Cymbopogon; CRC Press: Boca Raton, FL, USA, 2009. [Google Scholar]
- Hammer, O.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Paleontol. Electron. 2001, 4, 1–9. [Google Scholar]



{kind=link}
{kind=link}
| N° | RI A | RI P | Name | hys | mic | mapt | ich-2 | ich-3 | lat | wil | jun | maph |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 923 | 1022 | α-thujene | 0.2 | 0.1 | 0.1 | - | 0.1 | 1.0 | 0.4 | 0.4 | 0.3 |
| 2 | 931 | 1020 | α-pinene | 3.0 | 2.4 | 1.3 | - | 2.1 | 2.4 | 2.2 | 1.6 | 1.0 |
| 3 | 945 | 1070 | camphene | 0.2 | 0.3 | - | - | - | tr | tr | - | - |
| 4 | 966 | 1127 | sabinene | 22.7 | 1.0 | 12.4 | 0.1 | 9.6 | 0.3 | 0.1 | 0.1 | 0.1 |
| 5 | 972 | 1116 | β-pinene | 35.0 | 33.4 | 3.9 | - | 0.7 | 1.9 | 3.5 | 0.8 | 0.6 |
| 6 | 977 | 1221 | butyl butyrate | - | - | - | 2.4 | - | - | - | - | - |
| 7 | 981 | 1166 | myrcene | 0.9 | 1.0 | 2.1 | 0.2 | 2.1 | 18.8 | 1.6 | 1.9 | 1.6 |
| 8 | 996 | UD | hexyl acetate | - | - | - | - | - | - | 0.3 | - | - |
| 9 | 998 | 1170 | α-phellandrene | - | 0.1 | - | - | 0.3 | - | 0.2 | 0.4 | - |
| 10 | 1010 | 1185 | α-terpinene | - | 0.4 | 0.3 | - | 0.1 | 0.2 | 0.1 | 0.2 | tr |
| 11 | 1013 | 1276 | p-cymene | 0.7 | 0.8 | 1.1 | 0.3 | 4.4 | 6.5 | 7.0 | 1.3 | 2.6 |
| 12 | 1022 | 1215 | β-phellandrene * | 0.3 | 1.0 | 0.5 | - | 8.8 | 0.2 | 3.9 | 2.6 | 0.1 |
| 13 | 1022 | 1205 | limonene * | 25.2 | 20.7 | 53.8 | 42.3 | 58.2 | 50.4 | 66.9 | 79.9 | 81.4 |
| 14 | 1026 | 1237 | (Z)-β-ocimene | - | - | - | - | tr | - | - | - | 0.5 |
| 15 | 1037 | 1255 | (E)-β-ocimene | - | 0.4 | 0.2 | - | 0.1 | 0.1 | 0.2 | 0.4 | 0.8 |
| 16 | 1049 | 1251 | γ-terpinene | - | 1.3 | 1.0 | - | 1.0 | 16.2 | 10.1 | 8.8 | 5.0 |
| 17 | 1062 | 1446 | cis-linalool oxide THF form | 0.5 | 0.3 | 1.2 | - | tr | - | tr | - | 0.1 |
| 18 | 1075 | 1474 | trans-linalool oxide THF form | 0.3 | 0.2 | 0.6 | - | - | - | - | - | 0.1 |
| 19 | 1079 | 1288 | terpinolene | - | 1.1 | 0.3 | - | 0.3 | 0.7 | 0.5 | 0.4 | 0.2 |
| 20 | 1086 | 1551 | linalool | 0.9 | 2.2 | 11.8 | 0.4 | 0.7 | 0.1 | 0.3 | 1.0 | 0.3 |
| 21 | 1111 | 1565 | cis-p-menth-2-en-1-ol | tr | - | 0.2 | - | 0.3 | - | - | - | - |
| 22 | 1133 | 1574 | isopulegol | - | 1.5 | - | - | - | - | - | - | - |
| 23 | 1133 | 1483 | citronellal | 3.4 | 1.5 | - | - | - | - | - | - | tr |
| 24 | 1144 | 1567 | isoneopulegol | - | 1.0 | - | - | - | - | - | - | - |
| 25 | 1160 | 1672 | cryptone | - | - | - | - | 0.4 | - | 0.1 | - | - |
| 26 | 1163 | 1604 | terpinen-4-ol | 1.2 | 3.8 | 4.3 | tr | 7.3 | 0.1 | 0.5 | 0.1 | 0.1 |
| 27 | 1175 | 1699 | α-terpineol | 0.8 | 6.6 | 0.4 | 0.3 | 0.5 | 0.2 | 0.7 | 0.1 | 0.2 |
| 28 | 1173 | 1415 | butyl hexanoate | - | - | - | 1.5 | - | - | - | - | - |
| 29 | 1175 | 1417 | hexyl butyrate | - | - | - | 0.8 | - | - | - | - | - |
| 30 | 1199 | 1837 | trans-carveol | 0.2 | - | - | 0.6 | - | - | tr | - | tr |
| 31 | 1212 | 1769 | citronellol | 0.1 | 6.8 | - | - | - | - | - | - | - |
| 32 | 1217 | 1683 | neral | - | - | - | - | - | - | - | - | 0.5 |
| 33 | 1237 | 1851 | geraniol | tr | 1.0 | 0.1 | - | - | - | - | - | - |
| 34 | 1245 | 1753 | geranial | - | - | - | - | - | - | - | - | 0.6 |
| 35 | 1309 | 2275 | limonene-1,2-diol | - | - | - | 0.6 | tr | - | 0.1 | - | - |
| 36 | 1333 | 1697 | α-terpinyl acetate | - | - | - | - | - | - | 0.3 | - | - |
| 37 | 1334 | 1664 | citronellyl acetate | - | 3.1 | - | - | - | - | - | - | - |
| 38 | 1361 | 1759 | geranyl acetate | 1.1 | 2.1 | - | - | 0.2 | tr | tr | - | 0.1 |
| 39 | 1369 | 1611 | hexyl hexanoate | 0.1 | - | - | 0.5 | - | - | - | - | - |
| 40 | 1370 | 1614 | butyl octanoate | - | - | - | 0.8 | - | - | - | - | - |
| 41 | 1375 | 1492 | α-copaene | 0.3 | 0.2 | 0.4 | 0.1 | - | - | - | - | tr |
| 42 | 1387 | 1591 | β-elemene | 0.2 | 0.2 | 0.4 | - | tr | - | - | - | 0.1 |
| 43 | 1410 | 1569 | cis-α-bergamotene | - | - | - | 0.3 | - | - | - | - | tr |
| 44 | 1417 | 1597 | (E)-β-caryophyllene | 0.1 | 0.1 | 1.7 | - | - | 0.1 | - | - | 0.3 |
| 45 | 1417 | 1572 | α-santalene | - | - | - | - | 0.5 | - | - | - | - |
| 46 | 1432 | 1586 | trans-α-bergamotene | - | - | - | 3.2 | - | - | - | - | 0.6 |
| 47 | 1447 | 1667 | (E)-β-farnesene | - | - | - | 1.8 | - | - | 0.2 | 0.1 | tr |
| 48 | 1469 | 1688 | γ-muurolene | - | - | - | 3.1 | - | - | - | - | - |
| 49 | 1475 | 1708 | germacrene D | - | 0.2 | 0.3 | - | 0.1 | 0.7 | - | - | 0.6 |
| 50 | 1481 | 1718 | β-selinene | - | tr | - | 2.0 | - | - | - | - | - |
| 51 | 1488 | 1718 | valencene | - | - | - | 2.7 | - | - | 0.1 | - | - |
| 52 | 1490 | 1723 | α-selinene | tr | tr | 0.1 | 0.7 | - | - | - | - | - |
| 53 | 1495 | 1750 | (E,E)-α-farnesene | - | 1.7 | - | - | - | - | - | - | 0.1 |
| 54 | 1500 | 1727 | β-bisabolene | - | - | - | 18.4 | - | - | - | - | 0.9 |
| 55 | 1505 | 1758 | γ-cadinene | - | - | - | 1.1 | - | - | - | - | - |
| 56 | 1513 | 1757 | δ-cadinene | 0.1 | 0.3 | 0.5 | - | - | - | tr | - | 0.1 |
| 57 | 1548 | 2043 | (E)-nerolidol | - | tr | 0.2 | 1.6 | 0.9 | - | - | - | - |
| 58 | 1550 | 1826 | germacrene B | - | 0.3 | - | - | - | - | - | - | tr |
| 59 | 1611 | 2254 | alismol | - | 0.3 | 0.1 | 1.0 | - | - | - | - | 0.1 |
| 60 | 1641 | 2229 | intermedeol | - | - | - | 4.7 | - | - | - | - | - |
| Monoterpene hydrocarbon | 88.2 | 63.7 | 76.9 | 42.8 | 87.7 | 98.7 | 96.7 | 98.7 | 94.3 | |||
| Oxygenated monoterpene | 8.5 | 30.1 | 18.6 | 1.9 | 9.4 | 0.4 | 2.0 | 1.2 | 2.0 | |||
| Sesquiterpene hydrocarbon | 0.7 | 3.2 | 3.4 | 33.4 | 0.6 | 0.7 | 0.3 | 0.1 | 2.6 | |||
| Oxygenated sesquiterpene | 0.0 | 0.3 | 0.3 | 7.3 | 0.9 | 0.0 | 0.0 | 0.0 | 0.1 | |||
| Acyclic compound | 0.1 | 0.0 | 0.0 | 6.0 | 0.0 | 0.0 | 0.3 | 0.0 | 0.0 | |||
| TOTAL | 97.5 | 97.3 | 99.3 | 91.5 | 98.6 | 99.9 | 99.3 | 99.9 | 98.9 |
| N° | RI A | RI P | Name | hys | mic | mapt | ich-1 | ich-2 | ich-3 | lat | wil | jun | maph |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 923 | 1022 | α-thujene | tr | - | 0.3 | - | tr | 0.5 | - | 0.8 | 2.0 | 0.2 |
| 2 | 931 | 1020 | α-pinene | 0.1 | - | 2.0 | tr | tr | 2.2 | 0.1 | 2.0 | 4.8 | 0.5 |
| 3 | 965 | 1342 | 6-methyl-5-hepten-2-one | - | - | - | - | - | - | - | 0.3 | - | 0.5 |
| 4 | 966 | 1127 | sabinene | 3.0 | tr | 32.4 | - | tr | 44.6 | 0.2 | 1.4 | 0.7 | 0.1 |
| 5 | 972 | 1116 | β-pinene | 0.5 | 0.1 | 15.7 | - | tr | 1.6 | 0.3 | 9.7 | 4.1 | 0.5 |
| 6 | 981 | 1166 | myrcene | 0.5 | 0.4 | 1.8 | 1.7 | 1.1 | 2.3 | 0.7 | 0.8 | 1.4 | 0.6 |
| 7 | 998 | 1170 | α-phellandrene | - | - | 0.2 | 0.1 | 0.1 | 1.3 | - | 0.3 | 1.7 | - |
| 8 | 1006 | 1153 | δ-3-carene | tr | - | 1.9 | 0.4 | 0.7 | - | - | - | - | tr |
| 9 | 1010 | 1185 | α-terpinene | 0.1 | - | 1.0 | - | - | 2.2 | - | 0.3 | 0.8 | tr |
| 10 | 1013 | 1276 | p-cymene | tr | - | 0.1 | - | tr | 0.2 | 0.2 | 5.1 | 11.4 | 4.3 |
| 11 | 1022 | 1215 | β-phellandrene * | 0.1 | - | 0.7 | 0.1 | 0.1 | 11.7 | tr | 3.4 | 11.2 | - |
| 12 | 1022 | 1205 | limonene * | 2.4 | 0.1 | 2.7 | 0.4 | 0.1 | 3.1 | 41.0 | 4.0 | 4.7 | 17.7 |
| 13 | 1026 | 1237 | (Z)-β-ocimene | tr | 0.1 | 1.8 | 13.0 | 18.2 | 0.7 | 0.1 | 0.1 | 0.2 | 0.3 |
| 14 | 1037 | 1255 | (E)-β-ocimene | 0.2 | 0.6 | 8.6 | 32.4 | 62.7 | 3.7 | 2.2 | 3.0 | 5.1 | 0.6 |
| 15 | 1049 | 1251 | γ-terpinene | 0.5 | - | 1.6 | tr | - | 3.5 | 0.1 | 19.5 | 28.2 | 6.2 |
| 16 | 1057 | 1467 | trans-sabinene hydrate | tr | tr | 0.2 | - | - | 0.6 | - | - | tr | tr |
| 17 | 1062 | 1446 | cis-linalool oxide THF form | tr | tr | 1.1 | - | tr | 0.2 | 0.1 | tr | - | 0.2 |
| 18 | 1073 | 1442 | p-cymenene | - | - | - | - | tr | - | tr | - | 6.2 | - |
| 19 | 1075 | 1474 | trans-linalool oxide THF form | tr | - | 0.6 | - | tr | 0.1 | tr | - | - | 0.1 |
| 20 | 1079 | 1288 | terpinolene | 0.1 | tr | 0.7 | 0.3 | tr | 0.8 | tr | 0.9 | 2.0 | 0.2 |
| 21 | 1086 | 1551 | linalool | 3.4 | 1.2 | 18.2 | 9.3 | 0.2 | 1.0 | 24.6 | 6.1 | 10.4 | 4.3 |
| 22 | 1087 | 1550 | cis-sabinene hydrate | - | - | - | - | - | 0.5 | - | - | - | - |
| 23 | 1111 | 1565 | cis-p-menth-2-en-1-ol | - | - | 0.2 | - | - | 0.5 | - | - | - | - |
| 24 | 1117 | 1375 | allo-ocimene | - | - | - | 0.4 | 0.4 | - | - | - | - | - |
| 25 | 1126 | 1630 | trans-p-menth-2-en-1-ol | - | - | 0.1 | - | - | 0.3 | - | - | 0.6 | - |
| 26 | 1133 | 1574 | isopulegol | 0.8 | 0.9 | - | - | - | - | 0.2 | - | - | 0.1 |
| 27 | 1133 | 1483 | citronellal | 78.1 | 76.1 | - | - | - | 0.3 | 14.1 | 1.0 | - | 3.5 |
| 28 | 1145 | 1567 | isoneopulegol | 0.3 | 0.3 | - | - | - | - | tr | - | - | - |
| 29 | 1159 | UD | isogeranial | - | - | - | - | - | - | - | 0.2 | tr | 0.4 |
| 30 | 1163 | 1604 | terpinen-4-ol | 0.3 | - | 3.8 | tr | tr | 8.4 | - | 0.4 | 0.2 | 0.2 |
| 31 | 1175 | 1699 | α-terpineol | 0.1 | - | 0.2 | 3.1 | tr | 0.3 | 0.1 | 0.3 | 0.1 | 0.6 |
| 32 | 1212 | 1769 | citronellol | 3.4 | 4.4 | - | - | - | 0.1 | 1.8 | 0.2 | - | 0.1 |
| 33 | 1212 | 1804 | nerol | 0.1 | - | - | 0.8 | - | 0.1 | 0.1 | 2.3 | - | 0.2 |
| 34 | 1215 | 1597 | thymyl methyl oxide | - | - | - | - | - | - | - | - | 0.3 | - |
| 35 | 1217 | 1683 | neral | tr | - | - | - | - | 0.1 | 0.8 | 11.6 | - | 18.9 |
| 36 | 1237 | 1851 | geraniol | 0.6 | 1.2 | - | 2.3 | tr | 0.3 | 0.1 | 0.4 | - | 0.6 |
| 37 | 1241 | 1560 | linalyl acetate | - | - | - | 10.8 | - | - | 0.1 | - | - | 0.2 |
| 38 | 1245 | 1753 | geranial | 0.1 | - | - | - | tr | 0.2 | 1.0 | 15.2 | - | 24.7 |
| 39 | 1268 | 2192 | thymol | - | - | - | - | - | - | - | - | 1.1 | - |
| 40 | 1303 | 1697 | methyl geranate | - | - | - | - | - | 0.7 | - | - | - | - |
| 41 | 1334 | 1664 | citronellyl acetate | 0.7 | 5.1 | - | - | - | 0.3 | 1.0 | - | - | 0.4 |
| 42 | 1335 | 1472 | δ-elemene | - | 0.3 | - | - | - | - | - | 0.3 | 0.1 | 0.1 |
| 43 | 1343 | 1728 | neryl acetate | 0.1 | tr | - | 1.2 | - | 0.2 | 0.1 | 1.1 | - | 0.5 |
| 44 | 1361 | 1759 | geranyl acetate | 1.2 | 2.9 | - | 2.4 | - | 5.0 | 0.1 | 0.1 | - | 2.1 |
| 45 | 1375 | 1492 | α-copaene | 0.2 | 0.3 | - | 0.1 | tr | - | tr | - | - | 0.1 |
| 46 | 1387 | 1591 | β-elemene | tr | 0.5 | - | - | tr | - | tr | 0.3 | tr | 0.3 |
| 47 | 1399 | 1872 | 2,5-dimethoxy-p-cymene | - | - | - | - | - | - | - | - | 1.4 | - |
| 48 | 1417 | 1597 | (E)-β-caryophyllene | 1.1 | 0.8 | 0.5 | 1.4 | - | - | 2.9 | 0.1 | 0.1 | 2.7 |
| 49 | 1427 | 1638 | γ-elemene | tr | - | - | - | 0.3 | - | - | - | tr | - |
| 50 | 1432 | 1586 | trans-α-bergamotene | 0.1 | - | - | 0.1 | 0.1 | - | 1.1 | - | - | 0.6 |
| 51 | 1447 | 1667 | (E)-β-farnesene | - | 0.1 | - | 0.1 | 0.3 | - | - | tr | tr | tr |
| 52 | 1449 | 1667 | α-humulene | 0.1 | 0.1 | 0.1 | 0.2 | 0.2 | - | 0.2 | 0.1 | tr | 0.3 |
| 53 | 1469 | 1688 | γ-muurolene | - | 0.1 | - | 0.2 | 0.3 | - | - | - | - | tr |
| 54 | 1471 | 1668 | guaia-6,10(14)-diene | - | - | - | 0.3 | 0.4 | - | - | - | - | - |
| 55 | 1475 | 1708 | germacrene D | 0.1 | 0.2 | - | - | - | 0.1 | 0.6 | 1.2 | tr | 0.9 |
| 56 | 1481 | 1718 | β-selinene | - | - | - | 0.4 | 0.5 | - | tr | - | - | tr |
| 57 | 1490 | 1723 | α-selinene | - | - | - | tr | 0.3 | - | - | - | - | - |
| 58 | 1490 | 1732 | bicyclogermacrene | 0.3 | 0.2 | 0.4 | - | - | 0.1 | 0.3 | 0.1 | 0.1 | 0.4 |
| 59 | 1495 | 1750 | (E,E)-α-farnesene | 0.2 | 0.8 | 0.3 | - | 0.4 | 0.1 | - | - | - | 0.2 |
| 60 | 1500 | 1727 | β-bisabolene | 0.1 | - | - | 0.7 | 1.2 | - | 1.5 | 0.1 | - | 0.9 |
| 61 | 1513 | 1757 | δ-cadinene | 0.3 | 0.3 | 0.1 | 0.5 | 0.3 | - | tr | - | tr | 0.2 |
| 62 | 1534 | 2079 | β-elemol | tr | 0.1 | - | - | - | - | - | 1.1 | - | - |
| 63 | 1548 | 2043 | (E)-nerolidol | 0.2 | tr | 0.3 | 0.5 | 0.6 | 1.3 | tr | 0.3 | tr | tr |
| 64 | 1549 | 1825 | germacrene B | - | 0.5 | - | 0.4 | 0.6 | tr | - | 0.4 | tr | 0.2 |
| 65 | 1563 | 2121 | spathulenol | - | - | 0.1 | 0.1 | tr | 0.1 | 0.1 | 0.1 | - | 0.7 |
| 66 | 1570 | 1978 | caryophyllene oxide | tr | - | - | 1.8 | - | - | 0.2 | - | - | 0.4 |
| 67 | 1592 | 2033 | humulene oxide II | - | - | - | 0.3 | 0.2 | - | - | - | - | tr |
| 68 | 1611 | 2254 | alismol | - | 0.1 | - | 1.7 | 1.6 | 0.1 | - | 0.4 | - | 0.3 |
| 69 | 1616 | 2197 | eremoligenol | - | - | - | 0.4 | - | - | 0.1 | 0.3 | - | - |
| 70 | 1618 | 2176 | γ-eudesmol | - | tr | - | 0.1 | - | - | tr | 0.6 | - | - |
| 71 | 1625 | 2169 | τ-cadinol | - | - | 0.1 | 0.3 | 0.1 | - | - | 0.2 | tr | 0.1 |
| 72 | 1634 | 2225 | β-eudesmol | - | - | - | 0.7 | 0.8 | - | 0.2 | 0.6 | tr | - |
| 73 | 1639 | 2216 | α-eudesmol | - | - | - | 1.1 | 0.1 | - | tr | 0.5 | - | - |
| 74 | 1651 | 2145 | β-bisabolol | - | - | - | 0.4 | - | - | - | - | - | - |
| 75 | 1668 | 2215 | α-bisabolol | - | - | - | 0.4 | 0.5 | - | 0.1 | - | - | 0.1 |
| 76 | 2098 | 2610 | (E)-phytol | - | 0.3 | 0.7 | 3.0 | 1.4 | tr | 1.1 | 0.2 | 0.1 | 0.5 |
| Monoterpene hydrocarbon | 7.4 | 1.3 | 71.5 | 48.6 | 83.4 | 78.3 | 44.9 | 51.2 | 84.4 | 31.1 | |||
| Oxygenated monoterpene | 88.9 | 92.2 | 24.4 | 30.0 | 0.2 | 19.1 | 44.1 | 39.1 | 14.1 | 57.2 | |||
| Sesquiterpene hydrocarbon | 2.5 | 4.2 | 1.3 | 4.3 | 4.8 | 0.3 | 6.6 | 2.4 | 0.3 | 7.0 | |||
| Oxygenated sesquiterpene | 0.2 | 0.2 | 0.6 | 7.7 | 3.9 | 1.5 | 0.6 | 4.1 | 0.0 | 1.5 | |||
| Oxygenated diterpene | 0.0 | 0.3 | 0.7 | 3.0 | 1.4 | 0.0 | 1.1 | 0.2 | 0.1 | 0.5 | |||
| TOTAL | 99.0 | 98.3 | 98.5 | 93.6 | 93.8 | 99.3 | 97.2 | 97.1 | 98.9 | 97.3 | |||
| Yields (%; w/w) | 0.05 | 0.02 | 0.02 | 0.18 | 0.17 | 0.08 | 0.08 | 0.03 | 0.04 | 0.10 |
| Scientific Name | Common Name | Sample | ICVN |
|---|---|---|---|
| C. hystrix DC. | Combava | hys | 0100630 |
| C. macroptera Montr. | Melanesian papeda | mapt | 0100686 |
| C. micrantha Wester | Biasong | mic | 0101140 |
| C. ichangensis Swingle | Ichang papeda | ich-1 | 0100687 |
| C. ichangensis Swingle | Ichang papeda | ich-2 | 0110241 |
| C. ichangensis Swingle | Ichang papeda | ich-3 | 0110240 |
| C. latipes (Swingle) Tanaka | Khasi papeda | lat | 0110243 |
| C. wilsonii Tanaka | Ichang lemon | wil | 0100844 |
| C. junos Siebold ex Tanaka | Yuzu | jun | 0100988 |
| C. macrophylla Wester | Alemow | maph | 0110058 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baccati, C.; Gibernau, M.; Paoli, M.; Ollitrault, P.; Tomi, F.; Luro, F. Chemical Variability of Peel and Leaf Essential Oils in the Citrus Subgenus Papeda (Swingle) and Few Relatives. Plants 2021, 10, 1117. https://doi.org/10.3390/plants10061117
Baccati C, Gibernau M, Paoli M, Ollitrault P, Tomi F, Luro F. Chemical Variability of Peel and Leaf Essential Oils in the Citrus Subgenus Papeda (Swingle) and Few Relatives. Plants. 2021; 10(6):1117. https://doi.org/10.3390/plants10061117
Chicago/Turabian StyleBaccati, Clémentine, Marc Gibernau, Mathieu Paoli, Patrick Ollitrault, Félix Tomi, and François Luro. 2021. "Chemical Variability of Peel and Leaf Essential Oils in the Citrus Subgenus Papeda (Swingle) and Few Relatives" Plants 10, no. 6: 1117. https://doi.org/10.3390/plants10061117