
Genetic Engineering of Lesquerella with Increased Ricinoleic Acid Content in Seed Oil

 ,
,
Abstract
:1. Introduction
2. Results
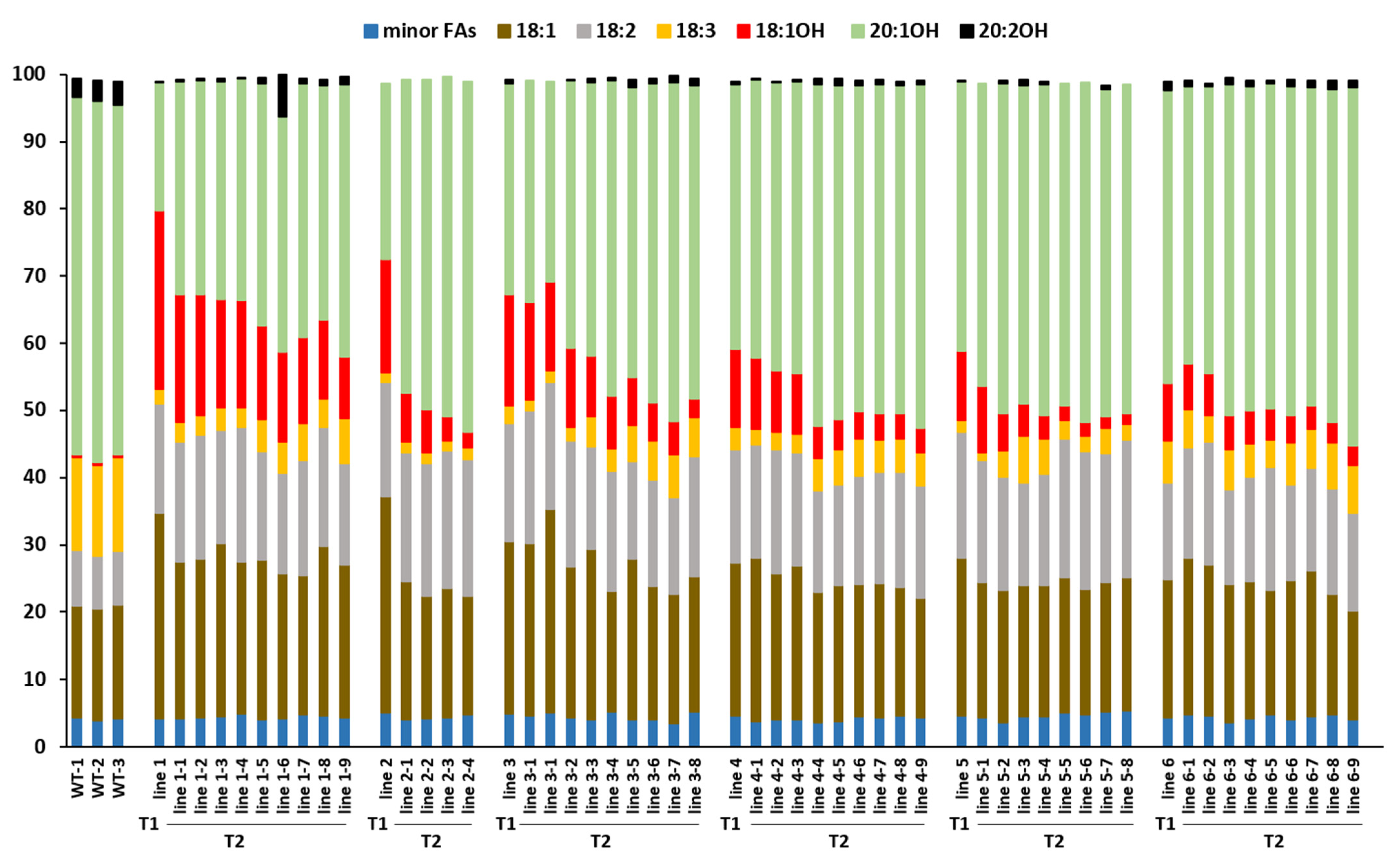
2.1. Changes of FA Composition in Transgenic Lesquerella Expressing Two dsRNAs, AtFAD3 RNAi + CsFAE1 RNAi
2.2. Changes of FA Composition in Transgenic Lesquerella Expressing Three dsRNAs, CsFAD2 RNAi + AtFAD3 RNAi + CsFAE1 RNAi
2.3. Correlations between FA Levels among Transgenic Lines
3. Discussion
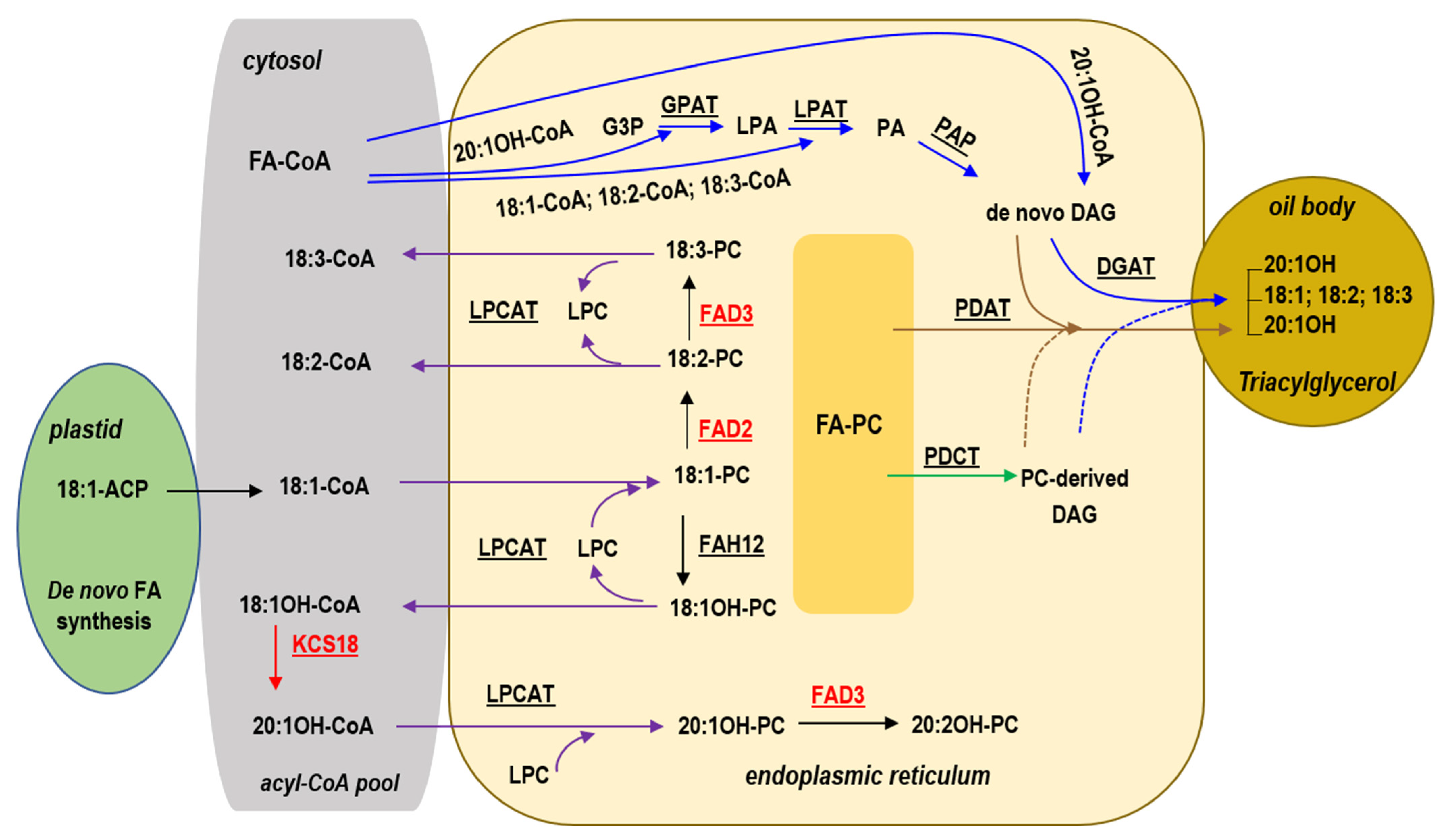
3.1. High Levels of 18:1OH Accumulate in Lesquerella by Blocking Elongation and Desaturation of Fatty Acids
3.2. Constrains and Potential for Production of a High 18:1OH-Containing Oil in Lesquerella
4. Materials and Methods
4.1. Construction of pBinGlyBar1 + AtFAD3 RNAi + CsFAE1 RNAi and pBinGlyBar1 CsFAD2 RNAi + AtFAD3 RNAi + CsFAE1RNAi
4.2. Plant Transformation and Growth Condition
4.3. Analysis of Fatty Acid Composition
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Caupin, H.J. Products from castor oil: Past, present and future. In Lipid Technologies and Applications; Gunstone, F.D., Padley, F.B., Eds.; Marcel Dekker: New York, NY, USA, 1997. [Google Scholar]
- Patel, V.R.; Dumancas, G.G.; Viswanath, L.C.K.; Maples, R.; Subong, B.J.J. Castor Oil: Properties, Uses, and Optimization of Processing Parameters in Commercial Production. Lipid Insights 2016, 9, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, G.Q.; He, X.; McKeon, T.A. A simple and sensitive assay for distinguishing the expression of ricin and Ricinus communis agglutinin genes in developing castor seed (R. communis L.). J. Agric. Food Chem. 2005, 53, 2358–2361. [Google Scholar] [CrossRef] [PubMed]
- Lord, M.J.; Jolliffe, N.A.; Marsden, C.J.; Pateman, C.S.C.; Smith, D.C.; Spooner, R.A.; Watson, P.D.; Roberts, L.M. Ricin: Mechanisms of cytotoxicity. Toxicol. Rev. 2003, 22, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.Q.; He, X.; Liao, L.P.; McKeon, T.A. 2S albumin gene expression in castor plant (Ricinus communis L.). J. Am. Oil Chem. Soc. 2004, 81, 867–872. [Google Scholar] [CrossRef]
- Machado, O.L.T.; Silva, J.G., Jr. An allergenic 2S storage protein from Ricinus communis seeds which is a part of the 2S albumin precursor predicted by C-DNA data. Braz. J. Med Biol. Res. 1992, 25, 567–582. [Google Scholar]
- Youle, R.J.; Huang, A.H.C. Albumin Storage Proteins in the Protein Bodies of Castor Bean. Plant Physiol. 1978, 61, 13–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roetheli, J.C.; Carlson, K.D.; Kleiman, R.; Thompson, A.E.; Dierig, D.; Glaser, L.K.; Blase, M.G.; Goodell, J. Lesquerella as a Source of Hydroxy Fatty Acids for Industrial Products; Growing Industrial Materials Series; USDA CSRS: Washington, DC, USA, 1991. Available online: https://www.ars.usda.gov/ARSUserFiles/53442010/NewCrops/Lesquerella/PDF/LesquerellaBooklet.pdf (accessed on 27 May 2021).
- Hayes, D.G.; Carlson, K.D.; Kleiman, R. The isolation of hydroxy acids from lesquerella oil lipolysate by a saponification/extraction technique. J. Am. Oil Chem. Soc. 1996, 73, 1113–1119. [Google Scholar] [CrossRef]
- Isbell, T.A.; Mund, M.S.; Evangelista, R.L.; Dierig, D.A. Method for analysis of fatty acid distribution and oil content on a single Lesquerella fendleri seed. Ind. Crop. Prod. 2008, 28, 231–236. [Google Scholar] [CrossRef]
- Chen, G.Q.; Lin, J.T.; Lu, C. Hydroxy fatty acid synthesis and lipid gene expression during seed development in Lesquerella fendleri. Ind. Crop. Prod. 2011, 34, 1286–1292. [Google Scholar] [CrossRef]
- Dierig, D.A.; Wang, G.; McCloskey, W.B.; Thorp, K.R.; Isbell, T.A.; Ray, D.T.; Foster, M.A. Lesquerella: New crop development and commercialization in the U.S. Ind. Crop. Prod. 2011, 34, 1381–1385. [Google Scholar] [CrossRef]
- Von Cruz, M.V.; Dierig, D.A. Germplasm improvement to develop commercially viable lines of the new oilseed crop lesquerella. In Industrial Crops: Breeding for Bioenergy and Bioproducts; Springer: New York, NY, USA, 2015; pp. 315–334. [Google Scholar] [CrossRef]
- Chen, G.Q. Effective reduction of chimeric tissue in transgenics for the stable genetic transformation of lesquerella fendleri. HortScience 2011, 46, 86–90. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.Q.; van Erp, H.; Martin-Moreno, J.; Johnson, K.; Morales, E.; Browse, J.; Eastmond, P.J.; Lin, J.T. Expression of castor LPAT2 enhances ricinoleic acid content at the sn-2 position of triacylglycerols in lesquerella seed. Int. J. Mol. Sci. 2016, 17, 507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li-Beisson, Y.; Shorrosh, B.; Beisson, F.; Andersson, M.X.; Arondel, V.; Bates, P.D.; Baud, S.; Bird, D.; DeBono, A.; Durrett, T.P.; et al. Acyl-Lipid Metabolism. Arab. Book Am. Soc. Plant Biol. 2013, 11, e0161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.U.; Chen, G.Q. Identification of hydroxy fatty acid and triacylglycerol metabolism-related genes in lesquerella through seed transcriptome analysis. BMC Genom. 2015, 16, 230. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.Q.; Kim, W.N.; Johnson, K.; Park, M.E.; Lee, K.R.; Kim, H.U. Transcriptome Analysis and Identification of Lipid Genes in Physaria lindheimeri, a Genetic Resource for Hydroxy Fatty Acids in Seed Oil. Int. J. Mol. Sci. 2021, 22, 514. [Google Scholar] [CrossRef]
- Bates, P.D. Understanding the control of acyl flux through the lipid metabolic network of plant oil biosynthesis. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2016, 1861, 1214–1225. [Google Scholar] [CrossRef] [PubMed]
- Lager, I.; Yilmaz, J.L.; Zhou, X.R.; Jasieniecka, K.; Kazachkov, M.; Wang, P.; Zou, J.; Weselake, R.; Smith, M.A.; Bayon, S.; et al. Plant acyl-CoA:lysophosphatidylcholine acyltransferases (LPCATs) have different specificities in their forward and reverse reactions. J. Biol. Chem. 2013, 288, 36902–36914. [Google Scholar] [CrossRef] [Green Version]
- Lands, W.E. Lipid Metabolism. Annu. Rev. Biochem. 1965, 34, 313–346. [Google Scholar] [CrossRef]
- Bafor, M.; Smith, M.A.; Jonsson, L.; Stobart, K.; Stymne, S. Ricinoleic acid biosynthesis and triacylglycerol assembly in microsomal preparations from developing castor-bean (Ricinus communis) endosperm. Biochem. J. 1991, 280, 507–514. [Google Scholar] [CrossRef] [Green Version]
- Moreau, R.A.; Stumpf, P.K. Recent Studies of the Enzymic Synthesis of Ricinoleic Acid by Developing Castor Beans. Plant Physiol. 1981, 67, 672–676. [Google Scholar] [CrossRef] [Green Version]
- Van De Loo, F.J.; Broun, P.; Turner, S.; Somerville, C. An oleate 12-hydroxylase from Ricinus communis L. is a fatty acyl desaturase homolog. Proc. Natl. Acad. Sci. USA 1995, 92, 6743–6747. [Google Scholar] [CrossRef] [Green Version]
- Broun, P.; Boddupalli, S.; Somerville, C. A bifunctional oleate 12-hydroxylase: Desaturase from Lesquerella fendleri. Plant J. 1998, 13, 201–210. [Google Scholar] [CrossRef]
- Bayon, S.; Chen, G.; Weselake, R.J.; Browse, J. A small phospholipase A2-α from castor catalyzes the removal of hydroxy fatty acids from phosphatidylcholine in transgenic Arabidopsis seeds. Plant Physiol. 2015, 167, 1259–1270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moon, H.; Smith, M.A.; Kunst, L. A condensing enzyme from the seeds of Lesquerella fendleri that specifically elongates hydroxy fatty acids. Plant Physiol. 2001, 127, 1635–1643. [Google Scholar] [CrossRef]
- Okuley, J.; Lightner, J.; Feldmann, K.; Yadav, N.; Lark, E.; Browse, J. Arabidopsis FAD2 gene encodes the enzyme that is essential for polyunsaturated lipid synthesis. Plant Cell 1994, 6, 147. [Google Scholar] [CrossRef]
- Arondel, V.; Lemieux, B.; Hwang, I.; Gibson, S.; Goodman, H.M.; Somerville, C.R. Map-based cloning of a gene controlling omega-3 fatty acid desaturation in Arabidopsis. Science 1992, 258, 1353–1355. [Google Scholar] [CrossRef] [PubMed]
- Engeseth, N.; Stymne, S. Desaturation of oxygenated fatty acids in Lesquerella and other oil seeds. Planta 1996, 198, 238–245. [Google Scholar] [CrossRef]
- Reed, D.W.; Taylor, D.C.; Covello, P.S. Metabolism of hydroxy fatty acids in developing seeds in the genera Lesquerella (Brassicaceae) and Linum (Linaceae). Plant Physiol. 1997, 114, 63–68. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.-R.; Kim, E.-H.; Jeon, I.; Lee, Y.; Chen, G.Q.; Kim, H.U. Lesquerella FAD3-1 gene is responsible for the biosynthesis of trienoic acid and dienoic hydroxy fatty acids in seed oil. Ind. Crop. Prod. 2019, 134, 257–264. [Google Scholar] [CrossRef] [Green Version]
- Kennedy, E.P. Biosynthesis of complex lipids. Fed. Proc. 1961, 20, 934–940. [Google Scholar] [PubMed]
- Hayes, D.G.; Kleiman, R. 1,3-specific lipolysis of lesquerella fendleri oil by immobilized and reverse-micellar encapsulated enzymes. J. Am. Oil Chem. Soc. 1993, 70, 1121–1127. [Google Scholar] [CrossRef]
- Lin, J.-T.; Chen, G.Q. Quantification of the Molecular Species of TAG and DAG in Lesquerella (Physaria fendleri) Oil by HPLC and MS. J. Am. Oil Chem. Soc. 2014, 91, 1417–1424. [Google Scholar] [CrossRef]
- Lin, J.-T.; Fagerquist, C.K.; Chen, G.Q. Ratios of Regioisomers of the Molecular Species of Triacylglycerols in Lesquerella (Physaria fendleri) Oil Estimated by Mass Spectrometry. J. Am. Oil Chem. Soc. 2016, 93, 183–191. [Google Scholar] [CrossRef]
- Chen, G.Q.; Lin, J.-T.; van Erp, H.; Johnson, K.; Lu, C. Regiobiochemical analysis reveals the role of castor LPAT2 in the accumulation of hydroxy fatty acids in transgenic lesquerella seeds. Biocatal. Agric. Biotechnol. 2020, 25, 101617. [Google Scholar] [CrossRef]
- Frentzen, M. Acyltransferases from basic science to modified seed oils. Fett 1998, 100, 161–166. [Google Scholar] [CrossRef]
- Lu, C.; Xin, Z.; Ren, Z.; Miquel, M.; Browse, J. An enzyme regulating triacylglycerol composition is encoded by the ROD1 gene of Arabidopsis. Proc. Natl. Acad. Sci. USA 2009, 106, 18837–18842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bates, P.D.; Browse, J. The pathway of triacylglycerol synthesis through phosphatidylcholine in Arabidopsis produces a bottleneck for the accumulation of unusual fatty acids in transgenic seeds. Plant J. 2011, 68, 387–399. [Google Scholar] [CrossRef]
- Hu, Z.; Ren, Z.; Lu, C. The phosphatidylcholine diacylglycerol cholinephosphotransferase is required for efficient hydroxy fatty acid accumulation in transgenic Arabidopsis. Plant Physiol. 2012, 158, 1944–1954. [Google Scholar] [CrossRef] [Green Version]
- Slack, C.R.; Campbell, L.C.; Browse, J.A.; Roughan, P.G. Some evidence for the reversibility of the cholinephosphotransferasecatalysed reaction in developing linseed cotyledons in vivo. Biochim. Biophys. Acta Lipids Lipid Metab. 1983, 754, 10–20. [Google Scholar] [CrossRef]
- Aryal, N.; Lu, C. A Phospholipase C-Like Protein From Ricinus communis Increases Hydroxy Fatty Acids Accumulation in Transgenic Seeds of Camelina sativa. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Dahlqvist, A.; Stahl, U.; Lenman, M.; Banas, A.; Lee, M.; Sandager, L.; Ronne, H.; Stymne, S. Phospholipid:diacylglycerol acyltransferase: An enzyme that catalyzes the acyl-CoA-independent formation of triacylglycerol in yeast and plants. Proc. Natl. Acad. Sci. USA 2000, 97, 6487–6492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.U.; Lee, K.R.; Go, Y.S.; Jung, J.H.; Suh, M.C.; Kim, J.B. Endoplasmic reticulum-located PDAT1-2 from castor bean enhances hydroxy fatty acid accumulation in transgenic plants. Plant Cell Physiol. 2011, 52, 983–993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Erp, H.; Bates, P.D.; Burgal, J.; Shockey, J.; Browse, J. Castor phospholipid:diacylglycerol acyltransferase facilitates efficient metabolism of hydroxy fatty acids in transgenic Arabidopsis. Plant Physiol. 2011, 155, 683–693. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.-R.; Chen, G.Q.; Kim, H.U. Current progress towards the metabolic engineering of plant seed oil for hydroxy fatty acids production. Plant Cell Rep. 2015. [Google Scholar] [CrossRef]
- Broun, P.; Somerville, C.; Morell, M.K.; Blennow, A.; Kosar-Hashemi, B.; Samuel, M.S. Accumulation of ricinoleic, lesquerolic, and densipolic acids in seeds of transgenic arabidopsis plants that express a fatty acyl hydroxylase cDNA from castor bean. Plant Physiol. 1997, 113, 933–942. [Google Scholar] [CrossRef] [Green Version]
- Lu, C.; Fulda, M.; Wallis, J.G.; Browse, J. A high-throughput screen for genes from castor that boost hydroxy fatty acid accumulation in seed oils of transgenic Arabidopsis. Plant J. 2006, 45, 847–856. [Google Scholar] [CrossRef]
- Burgal, J.; Shockey, J.; Lu, C.; Dyer, J.; Larson, T.; Graham, I.; Browse, J. Metabolic engineering of hydroxy fatty acid production in plants: RcDGAT2 drives dramatic increases in ricinoleate levels in seed oil. Plant Biotechnol. J. 2008, 6, 819–831. [Google Scholar] [CrossRef] [Green Version]
- Shockey, J.; Lager, I.; Stymne, S.; Kotapati, H.K.; Sheffield, J.; Mason, C.; Bates, P.D. Specialized lysophosphatidic acid acyltransferases contribute to unusual fatty acid accumulation in exotic Euphorbiaceae seed oils. Planta 2019, 249, 1285–1299. [Google Scholar] [CrossRef]
- Kim, H.U.; Park, M.-E.; Lee, K.-R.; Suh, M.-C.; Chen, G.Q. Variant Castor Lysophosphatidic Acid Acyltransferases Acylate Ricinoleic Acid in Seed Oil. Ind. Crop. Prod. 2020, 150, 112245. [Google Scholar] [CrossRef]
- Lunn, D.; Wallis, J.G.; Browse, J. Tri-Hydroxy-Triacylglycerol Is Efficiently Produced by Position-Specific Castor Acyltransferases. Plant Physiol. 2019, 179, 1050–1063. [Google Scholar] [CrossRef] [Green Version]
- Baulcombe, D. RNA silencing in plants. Nature 2004, 431, 356–363. [Google Scholar] [CrossRef]
- Brodersen, P.; Voinnet, O. The diversity of RNA silencing pathways in plants. Trends Genet. 2006, 22, 268–280. [Google Scholar] [CrossRef]
- Kang, J.; Snapp, A.R.; Lu, C. Identification of three genes encoding microsomal oleate desaturases (FAD2) from the oilseed crop Camelina sativa. Plant Physiol. Biochem. 2011, 49, 223–229. [Google Scholar] [CrossRef]
- Nguyen, H.T.; Silva, J.E.; Podicheti, R.; Macrander, J.; Yang, W.; Nazarenus, T.J.; Nam, J.W.; Jaworski, J.G.; Lu, C.; Scheffler, B.E.; et al. Camelina seed transcriptome: A tool for meal and oil improvement and translational research. Plant Biotechnol. J. 2013, 11, 759–769. [Google Scholar] [CrossRef] [Green Version]
- Kumssa, T.T.; Nazarenus, T.J.; Koster, K.L.; Nguyen, H.T.; Cahoon, R.E.; Lu, C.; Cahoon, E.B. Field Performance and Enhanced Oil Oxidative Stability of the Biofuel and Industrial Oilseed Camelina Engineered for Reduced Fatty Acid Polyunsaturation. 2021; in preparation. [Google Scholar]
- Hu, Q.A.; Ailhas, J.; Blevins, T.; Klahre, U.; Vazquez, F.; Mette, M.F.; Meins, F. Epigenetic transgenerational effects on RNAi in Arabidopsis. bioRxiv 2020. [Google Scholar] [CrossRef]
- McGinnis, K.; Murphy, N.; Carlson, A.R.; Akula, A.; Akula, C.; Basinger, H.; Carlson, M.; Hermanson, P.; Kovacevic, N.; McGill, M.A.; et al. Assessing the Efficiency of RNA Interference for Maize Functional Genomics. Plant Physiol. 2007, 143, 1441–1451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, G.Q.; Turner, C.; He, X.; Nguyen, T.; McKeon, T.A.; Laudencia-Chingcuanco, D. Expression profiles of genes involved in fatty acid and triacylglycerol synthesis in castor bean (Ricinus communis L.). Lipids 2007, 42, 263–274. [Google Scholar] [CrossRef] [PubMed]
- Dauk, M.; Lam, P.; Kunst, L.; Smith, M.A. A FAD2 homologue from Lesquerella lindheimeri has predominantly fatty acid hydroxylase activity. Plant Sci. 2007, 173, 43–49. [Google Scholar] [CrossRef] [Green Version]
- Waschburger, E.; Kulcheski, F.R.; Veto, N.M.; Margis, R.; Margis-Pinheiro, M.; Turchetto-Zolet, A.C. Genome-wide analysis of the Glycerol-3-Phosphate Acyltransferase (GPAT) gene family reveals the evolution and diversification of plant GPATs. Genet. Mol. Biol. 2018, 41, 355–370. [Google Scholar] [CrossRef] [PubMed]
- Kroon, J.T.M.; Wei, W.; Simon, W.J.; Slabas, A.R. Identification and functional expression of a type 2 acyl-CoA:diacylglycerol acyltransferase (DGAT2) in developing castor bean seeds which has high homology to the major triglyceride biosynthetic enzyme of fungi and animals. Phytochemistry 2006, 67, 2541–2549. [Google Scholar] [CrossRef]
- Snapp, A.R.; Kang, J.; Qi, X.; Lu, C. A fatty acid condensing enzyme from Physaria fendleri increases hydroxy fatty acid accumulation in transgenic oilseeds of Camelina sativa. Planta 2014, 240, 599–610. [Google Scholar] [CrossRef]
- Lu, C.; Kang, J. Generation of transgenic plants of a potential oilseed crop Camelina sativa by Agrobacterium-mediated transformation. Plant Cell Rep. 2008, 27, 273–278. [Google Scholar] [CrossRef] [PubMed]
- Moire, L.; Rezzonico, E.; Goepfert, S.; Poirier, Y. Impact of unusual fatty acid synthesis on futile cycling through beta-oxidation and on gene expression in transgenic plants. Plant Physiol. 2004, 134, 432–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stahl, U.; Carlsson, A.S.; Lenman, M.; Dahlqvist, A.; Huang, B.; Banas, W.; Banas, A.; Stymne, S. Cloning and functional characterization of a phospholipid:diacylglycerol acyltransferase from Arabidopsis. Plant Physiol. 2004, 135, 1324–1335. [Google Scholar] [CrossRef] [Green Version]
- Pidkowich, M.S.; Nguyen, H.T.; Heilmann, I.; Ischebeck, T.; Shanklin, J. Modulating seed beta-ketoacyl-acyl carrier protein synthase II level converts the composition of a temperate seed oil to that of a palm-like tropical oil. Proc. Natl. Acad. Sci. USA 2007, 104, 4742–4747. [Google Scholar] [CrossRef] [Green Version]
- Wesley, S.V.; Helliwell, C.A.; Smith, N.A.; Wang, M.B.; Rouse, D.T.; Liu, Q.; Gooding, P.S.; Singh, S.P.; Abbott, D.; Stoutjesdijk, P.A.; et al. Construct design for efficient, effective and high-throughput gene silencing in plants. Plant J. 2001, 27, 581–590. [Google Scholar] [CrossRef]
- Dierig, D.A.; Tomasi, P.M.; Dahlquist, G.H. Registration of WCL-LY2 high oil Lesquerella fendleri germplasm. Crop Sci. 2001, 41, 604–605. [Google Scholar] [CrossRef]
- Lazo, G.R.; Stein, P.A.; Ludwig, R.A. A DNA transformation-competent Arabidopsis genomic library in Agrobacterium. Nat. Biotechnol. 1991, 9, 963–967. [Google Scholar] [CrossRef]
- Butte, W. Rapid method for the determination of fatty acid profiles from fats and oils using trimethylsulphonium hydroxide for transesterification. J. Chromatogr. A 1983, 261, 142–145. [Google Scholar] [CrossRef]
- Kim, H.J.; Silva, J.E.; Vu, H.S.; Mockaitis, K.; Nam, J.W.; Cahoon, E.B. Toward production of jet fuel functionality in oilseeds: Identification of FatB acyl-acyl carrier protein thioesterases and evaluation of combinatorial expression strategies in Camelina seeds. J. Exp. Bot. 2015, 66, 4251–4265. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Line | Total Minor Fatty Acid a | 18:1 | 18:2 | 18:3 | 18:1OH | 20:1OH | 20:2OH | Total Hydroxy Fatty Acid |
|---|---|---|---|---|---|---|---|---|
| wild-type | 4.7 ± 0.4 | 17.0 ± 0.4 | 7.6 ± 0.4 | 13.3 ± 0.6 | 0.6 ± 0.2 | 51.2 ± 1.0 | 4.3 ± 0.6 | 56.0 ± 0.5 |
| line 1 | 4.3 ± 0.2 | 30.5 ± 2.8 *** | 16.2 ± 1.7 *** | 2.2 ± 0.7 *** | 26.6 ± 0.2 *** | 19.0 ± 2.0 *** | 0.2 ± 0.2 *** | 45.8 ± 1.9 *** |
| line 2 | 5.1 ± 0.2 | 32.1 ± 1.3 ** | 17.0 ± 0.5 *** | 1.7 ± 0.5 *** | 16.8 ± 0.5 *** | 26.1 ± 1.0 *** | 0.0 ± 0.0 *** | 42.9 ± 0.5 *** |
| line 3 | 5.0 ± 0.2 | 25.5 ± 1.9 ** | 17.6 ± 1.2 *** | 2.6 ± 1.4 *** | 16.6 ± 1.0 *** | 31.4 ± 2.1 *** | 0.5 ± 0.4 *** | 48.4 ± 1.5 *** |
| line 4 | 4.7 ± 0.4 | 22.7 ± 2.7 * | 16.8 ± 0.6 *** | 3.3 ± 0.8 *** | 11.7 ± 2.7 ** | 39.3 ± 5.0 ** | 0.4 ± 0.2 *** | 51.4 ± 2.6 * |
| line 5 | 4.7 ± 0.2 | 23.4 ± 2.0 ** | 18.7 ± 0.2 *** | 1.8 ± 0.6 *** | 10.2 ± 0.4 *** | 40.2 ± 1.4 *** | 0.1 ± 0.1 *** | 50.5 ± 1.5 *** |
| line 6 | 4.4 ± 0.1 | 20.5 ± 1.8 * | 14.4 ±1.0 *** | 6.2 ± 0.5 *** | 8.5 ± 0.4 *** | 43.7 ± 1.4 ** | 1.2 ± 0.1 *** | 53.4 ± 1.5 * |
| line 7 | 4.1 ± 0.1 | 20.5 ± 0.9 ** | 16.7 ± 2.0 *** | 4.9 ± 1.7 *** | 8.0 ± 1.3 *** | 45.5 ± 2.0 ** | 0.3 ± 0.2 *** | 53.8 ± 1.2 * |
| line 8 | 4.5 ± 0.0 | 17.6 ± 0.5 | 16.1 ± 0.5 *** | 7.6 ± 0.4 *** | 7.5 ± 0.5 *** | 45.0 ± 0.5 *** | 1.3 ± 0.2 *** | 53.8 ± 0.2 ** |
| line 9 | 4.4 ± 0.1 | 17.9 ± 0.5 | 17.2 ± 0.7 *** | 4.3 ± 0.7 *** | 4.9 ± 0.4 *** | 49.3 ± 0.6 * | 0.7 ± 0.2 *** | 54.9 ± 0.5 |
| line 10 | 4.8 ± 0.0 | 19.0 ± 1.0 * | 13.0 ± 0.5 *** | 9.6 ± 0.5 *** | 4.7 ± 1.2 ** | 46.9 ± 0.6 * | 1.2 ± 0.1 *** | 52.8 ± 1.4 * |
| line 11 | 4.3 ± 0.0 | 16.5 ± 0.4 | 16.3 ± 0.2 *** | 5.4 ± 0.3 *** | 3.6 ± 0.3 *** | 51.9 ± 0.4 | 0.8 ± 0.2 *** | 56.2 ± 0.0 |
| line 12 | 4.1 ± 0.2 | 16.3 ± 0.5 | 14.1 ± 0.9 *** | 7.7 ± 1.2 ** | 1.8 ± 0.3 ** | 53.2 ± 1.0 | 1.8 ± 0.2 ** | 56.7 ± 1.1 |
| line 13 | 3.9 ± 0.3 | 17.1 ± 2.1 | 9.5 ± 0.2 ** | 13.3 ± 0.4 | 1.2 ± 0.1 ** | 51.1 ± 2.7 | 2.2 ± 0.1 ** | 54.5 ± 2.7 |
| line 14 | 4.3 ± 0.3 | 16.2 ± 0.5 | 20.0 ± 0.7 *** | 1.5 ± 0.1 *** | 0.5 ± 0.1 | 56.2 ± 1.2 ** | 0.1 ± 0.0 *** | 56.8 ± 1.3 |
| line 15 | 4.0 ± 0.4 | 16.1 ± 0.5 | 14.2 ± 1.2 *** | 6.6 ± 0.9 *** | 0.5 ± 0.1 | 55.5 ± 0.7 ** | 1.8 ± 0.4 ** | 57.7 ± 0.8 * |
| line 16 | 4.1 ± 0.1 | 15.4 ± 0.4 ** | 13.4 ± 1.5 ** | 8.3 ± 1.3 ** | 0.5 ± 0.1 | 55.6 ± 0.2 ** | 1.7 ± 0.3 ** | 57.8 ± 0.5 * |
| average of transgenics | 4.4 ± 0.1 | 20.5 ± 0.9 | 15.7 ± 0.5 | 5.4 ± 0.4 | 7.7 ± 0.7 | 44.4 ± 1.2 | 0.9 ± 0.1 | 53.0 ± 0.8 |
| Line | Total Minor Fatty Acid a | 18:1 | 18:2 | 18:3 | 18:1OH | 20:1OH | 20:2OH | Total Hydroxy Fatty Acid |
|---|---|---|---|---|---|---|---|---|
| wild-type | 4.2 ± 0.2 | 16.7 ± 0.2 | 8.0 ± 0.2 | 13.7 ± 0.2 | 0.40 ± 0.0 | 53.0 ± 0.9 | 3.1 ± 0.5 | 56.5 ± 0.7 |
| line 1 | 4.2 ± 0.1 | 27.7 ± 0.3 *** | 13.8 ± 0.4 *** | 4.8 ± 0.5 *** | 15.4 ± 0.7 *** | 33.3 ± 0.8 *** | 0.9 ± 0.1 *** | 49.6 ± 0.1 *** |
| line 2 | 4.3 ± 0.1 | 26.4 ± 2.4 ** | 13.6 ± 0.3 *** | 5.5 ± 0.8 *** | 10.3 ± 0.9 *** | 38.8 ± 2.5 *** | 1.1 ± 0.1 ** | 50.1 ± 1.9 ** |
| line 3 | 5.2 ± 0.0 *** | 35.7 ± 1.6 ** | 15.1 ± 0.4 *** | 3.1 ± 1.0 *** | 8.2 ± 1.1 *** | 32.4 ± 1.8 *** | 0.5 ± 0.3 ** | 40.9 ± 1.1 *** |
| line 4 | 4.1 ± 0.3 | 22.6 ± 0.6 *** | 15.8 ± 1.5 *** | 4.3 ± 2.5 ** | 7.5 ± 1.0 *** | 44.9 ± 0.7 *** | 0.7 ± 0.6 ** | 53.2 ± 1.3 ** |
| line 5 | 5.4 ± 0.1 *** | 30.2 ± 1.0 *** | 13.2 ± 1.1 *** | 3.8 ± 0.3 *** | 6.5 ± 0.8 *** | 37.3 ± 1.0 *** | 0.7 ± 0.1 *** | 44.5 ± 1.8 *** |
| line 6 | 4.5 ± 0.0 * | 35.8 ± 4.1 *** | 15.1 ± 0.2 *** | 5.0 ± 0.7 *** | 6.3 ± 0.4 *** | 35.6 ± 3.7 *** | 0.7 ± 0.3 *** | 42.6 ± 4.1 ** |
| line 7 | 4.1 ± 0.1 | 17.5 ± 0.8 | 17.3 ± 1.2 *** | 3.9 ± 0.9 *** | 6.3 ± 0.5 *** | 50.2 ± 0.5 ** | 0.7 ± 0.0 *** | 57.2 ± 0.6 |
| line 8 | 4.3 ± 0.2 * | 38.8 ± 3.5 *** | 14.4 ± 0.9 *** | 1.7 ± 0.3 *** | 4.6 ± 0.3 *** | 36.1 ± 2.8 *** | 0 ± 0.3 *** | 40.7 ± 2.6 *** |
| line 9 | 3.9 ± 0.2 | 24.6 ± 2.1 ** | 13.3 ± 0.5 *** | 6.0 ± 0.5 *** | 1.9 ± 0.2 *** | 48.9 ± 1.4 ** | 1.2 ± 0.2 ** | 52.1 ± 1.4 ** |
| line 10 | 4.5 ± 0.8 | 32.3 ± 2.8 *** | 10.7 ± 1.3 *** | 7.1 ± 1.1 *** | 1.1 ± 0.2 *** | 43.1 ± 2.2 ** | 1.5 ± 0.3 ** | 45.7 ± 1.8 *** |
| line 11 | 4.8 ± 0.3 * | 22.9 ± 0.8 *** | 13.9 ± 0.1 *** | 7.0 ± 0.7 *** | 0.7 ± 0.1 ** | 49.7 ± 0.7 ** | 0.9 ± 0.1 ** | 51.3 ± 0.6 *** |
| line 12 | 4.3 ± 0.2 | 28.4 ± 1.4 *** | 16.0 ± 0.7 *** | 3.0 ± 0.2 *** | 0.4 ± 0.0 | 47.9 ± 0.7 ** | 0.0 ± 0.0 *** | 48.3 ± 0.7 *** |
| line 13 | 4.4 ± 0.1 | 22.7 ± 2.8 * | 17.4 ± 0.6 *** | 1.9 ± 0.4 *** | 0.4 ± 0.1 | 53.2 ± 2.0 | 0.0 ± 0.0 *** | 53.6 ± 2.0 |
| line 14 | 4.3 ± 0.3 | 24.3 ± 2.0 ** | 14.6 ± 1.1 *** | 4.5 ± 0.9 *** | 0.4 ± 0.0 | 50.9 ± 1.1* | 1.1 ± 0.5 ** | 52.4 ± 1.6 ** |
| line 15 | 4.0 ± 0.0 *** | 26.3 ± 1.6 *** | 14.5 ± 0.4 *** | 4.0 ± 0.6 *** | 0.4 ± 0.0 | 50.1 ± 0.9 * | 0.7 ± 0.2 *** | 51.2 ± 1.0 ** |
| average of transgenic line | 4.4 ± 0.4 | 27.8 ± 5.9 | 14.6 ± 1.7 | 4.4 ± 1.6 | 4.7 ± 4.5 | 43.5 ± 7.2 | 0.7 ± 0.4 | 48.9 ± 5.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, G.Q.; Johnson, K.; Nazarenus, T.J.; Ponciano, G.; Morales, E.; Cahoon, E.B. Genetic Engineering of Lesquerella with Increased Ricinoleic Acid Content in Seed Oil. Plants 2021, 10, 1093. https://doi.org/10.3390/plants10061093
Chen GQ, Johnson K, Nazarenus TJ, Ponciano G, Morales E, Cahoon EB. Genetic Engineering of Lesquerella with Increased Ricinoleic Acid Content in Seed Oil. Plants. 2021; 10(6):1093. https://doi.org/10.3390/plants10061093
Chicago/Turabian StyleChen, Grace Q., Kumiko Johnson, Tara J. Nazarenus, Grisel Ponciano, Eva Morales, and Edgar B. Cahoon. 2021. "Genetic Engineering of Lesquerella with Increased Ricinoleic Acid Content in Seed Oil" Plants 10, no. 6: 1093. https://doi.org/10.3390/plants10061093