
Echinacea purpurea L. (Moench) Hemagglutinin Effect on Immune Response In Vivo

 , , and
, , and
Abstract
:1. Introduction
2. Results
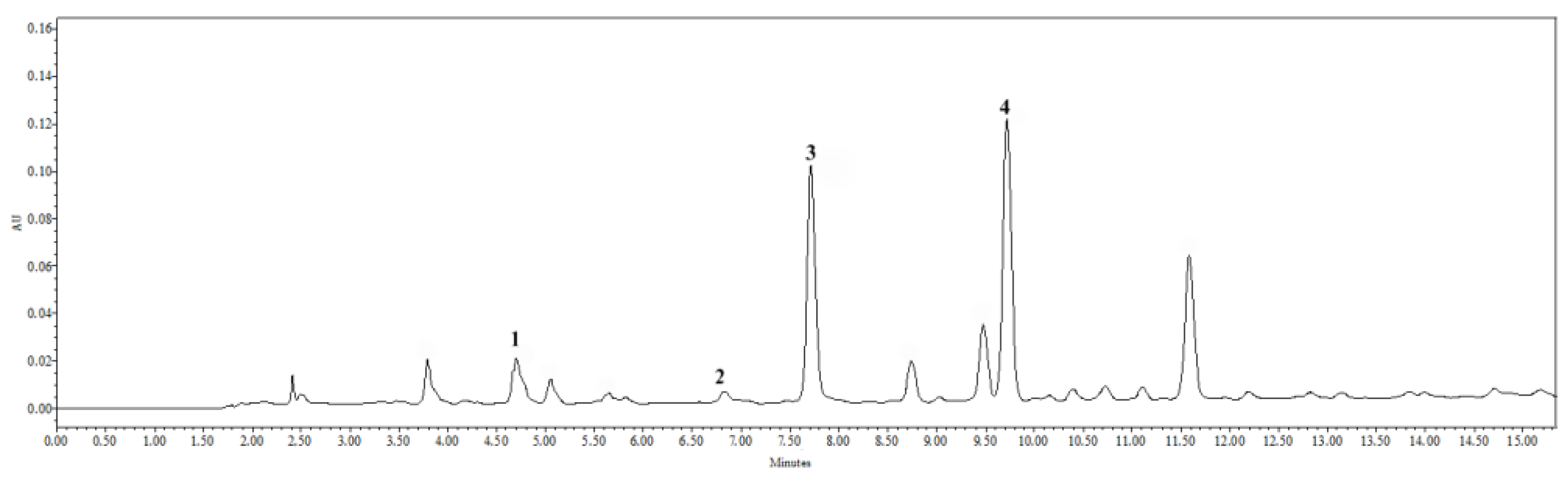
2.1. Results of Tincture Qualitative and Quantitative Assessment by HPLC
2.2. Animals during Experiment
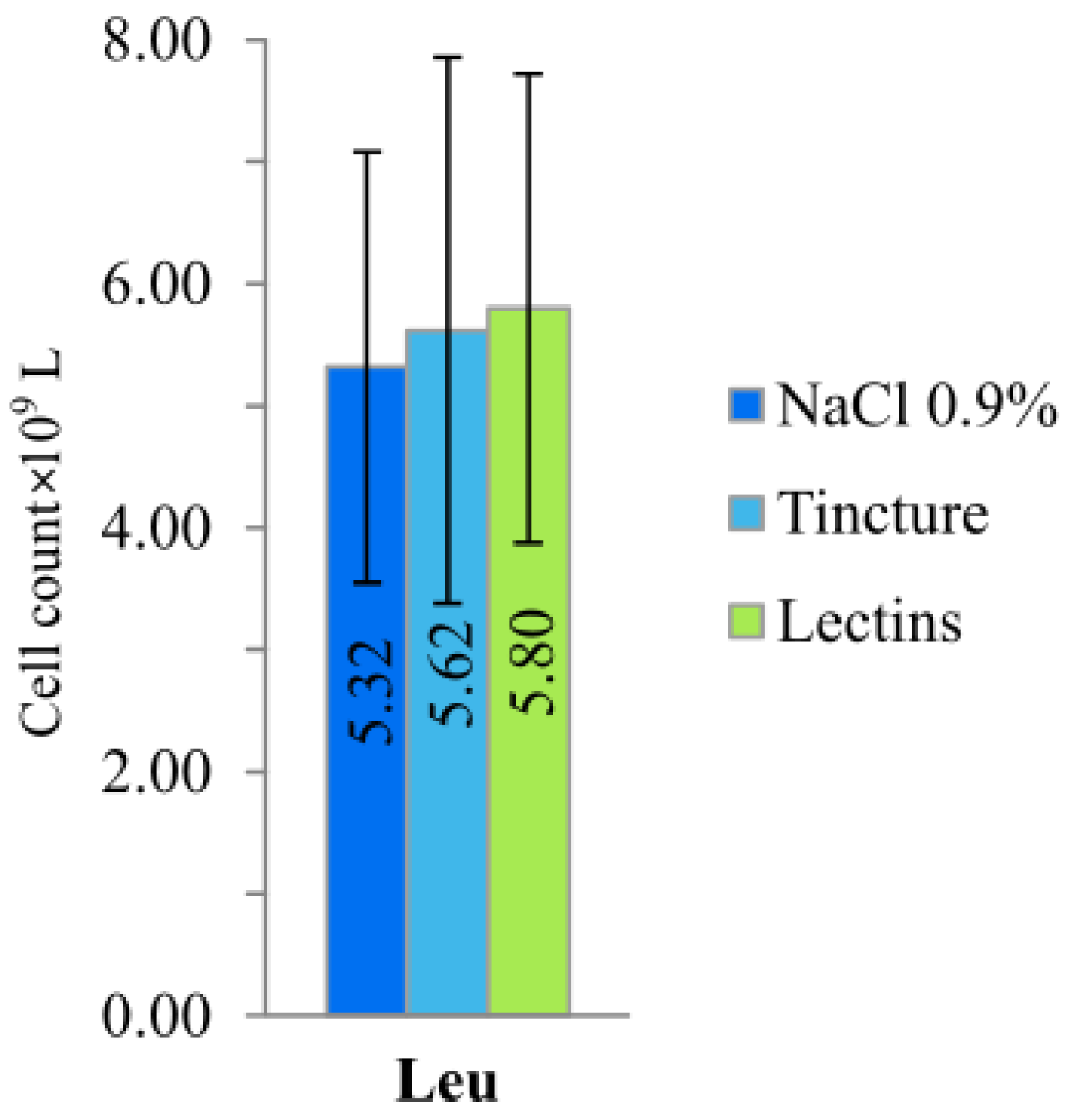
2.3. Blood Morphology and Cell Count
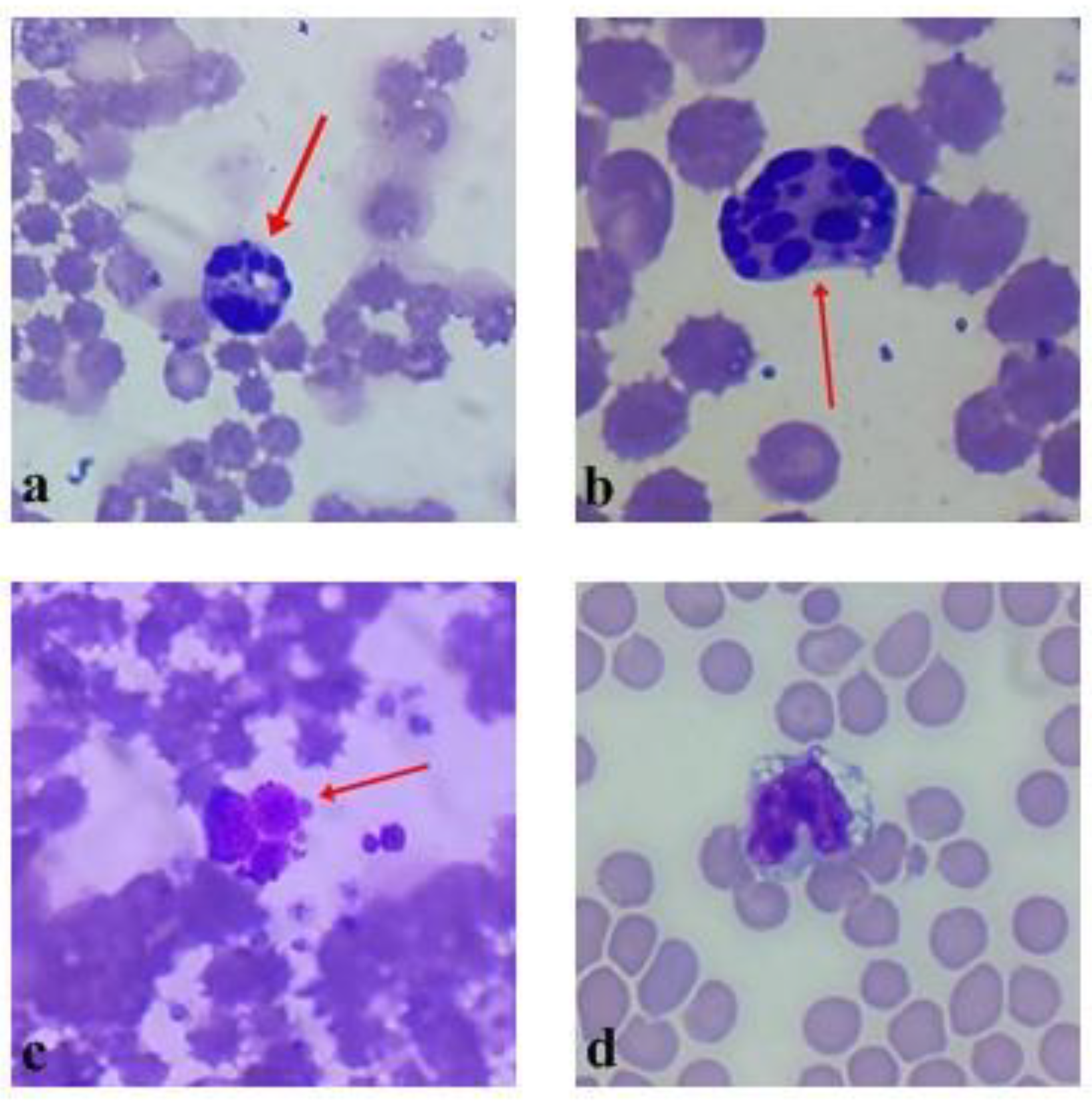
2.4. Spleen Immunohistochemical Investigation
3. Conclusions
4. Materials and Methods
4.1. Herbal Material Preparation
4.2. Lectin Purification and Identification
4.3. Echinacea purpurea (L.) Moench Fresh Roots’ Ttincture Preparation and Analysis
4.4. Animals
4.5. In Vivo Experiment Model
4.6. Blood Sample Analysis
4.7. Spleen
4.8. Evaluation of Spleen’s Sections
4.9. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hudson, J.; Vimalanathan, S.; Kang, L.; Amiguet, V.T.; Livesey, J.; Arnason, J.T. Characterization of Antiviral Activities in Echinacea Root Preparations. Pharm. Biol. 2005, 43, 790–796. [Google Scholar] [CrossRef]
- Sharifi-Rad, M.; Mnayer, D.; Morais-Braga, M.F.B.; Carneiro, J.N.P.; Bezerra, C.F.; Coutinho, H.D.M.; Salehi, B.; Martorell, M.; del Mar Contreras, M.; Soltani-Nejad, A.; et al. Echinacea Plants as Antioxidant and Antibacterial Agents: From Traditional Medicine to Biotechnological Applications. Phytother. Res. 2018, 32, 1653–1663. [Google Scholar] [CrossRef]
- Catanzaro, M.; Corsini, E.; Rosini, M.; Racchi, M.; Lanni, C. Immunomodulators Inspired by Nature: A Review on Curcumin and Echinacea. Mol. Basel Switz. 2018, 23, 2778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hudson, J.B. Applications of the Phytomedicine Echinacea purpurea (Purple Coneflower) in Infectious Diseases. J. Biomed. Biotechnol. 2011, 2012, e769896. [Google Scholar] [CrossRef] [Green Version]
- Dobrange, E.; Peshev, D.; Loedolff, B.; Van den Ende, W. Fructans as Immunomodulatory and Antiviral Agents: The Case of Echinacea. Biomolecules 2019, 9, 615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bauer, R. Echinacea Drugs--Effects and Active Ingredients. Z. Arztl. Fortbild. (Jena) 1996, 90, 111–115. [Google Scholar]
- Bany, J.; Siwicki, A.K.; Zdanowska, D.; Sokolnicka, I.; Skopińska-Rózewska, E.; Kowalczyk, M. Echinacea purpurea Stimulates Cellular Immunity and Anti-Bacterial Defence Independently of the Strain of Mice. Pol. J. Vet. Sci. 2003, 6, 3–5. [Google Scholar] [PubMed]
- Block, K.I.; Mead, M.N. Immune System Effects of Echinacea, Ginseng, and Astragalus: A Review. Integr. Cancer Ther. 2003, 2, 247–267. [Google Scholar] [CrossRef] [PubMed]
- Benson, J.M.; Pokorny, A.J.; Rhule, A.; Wenner, C.A.; Kandhi, V.; Cech, N.B.; Shepherd, D.M. Echinacea purpurea Extracts Modulate Murine Dendritic Cell Fate and Function. Food Chem. Toxicol. 2010, 48, 1170–1177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bronte, V.; Pittet, M.J. The Spleen in Local and Systemic Regulation of Immunity. Immunity 2013, 39, 806–818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, A.; Behura, A.; Mawatwal, S.; Kumar, A.; Naik, L.; Mohanty, S.S.; Manna, D.; Dokania, P.; Mishra, A.; Patra, S.K.; et al. Structure-Function and Application of Plant Lectins in Disease Biology and Immunity. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2019, 134, 110827. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, C.L. Lectins: Analytical Tools from Nature. In Lectins Analytical Technologies; Elsevier: Amsterdam, The Netherlands, 2007; pp. 1–13. [Google Scholar] [CrossRef]
- Coelho, L.C.B.B.; Silva, P.M.D.S.; Lima, V.L.D.M.; Pontual, E.V.; Paiva, P.M.G.; Napoleão, T.H.; Correia, M.T.D.S. Lectins, Interconnecting Proteins with Biotechnological/Pharmacological and Therapeutic Applications. Evid. Based Complement. Altern. Med. 2017, 2017, 1594074. [Google Scholar] [CrossRef]
- Balciunaite, G.; Haimi, P.-J.; Mikniene, Z.; Savickas, G.; Ragazinskiene, O.; Juodziukyniene, N.; Baniulis, D.; Pangonyte, D. Identification of Echinacea purpurea (L.) Moench Root LysM Lectin with Nephrotoxic Properties. Toxins 2020, 12, 88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Council of Europe; European Pharmacopoeia Commission. Echinaceae purpureae radix (01/2008:1824). In European Pharmacopoeia, 9.0; Council of Europe: Strasbourg, France, 2018; pp. 1488–1490. [Google Scholar]
- Jeong, J.A.; Wu, C.H.; Murthy, H.N.; Hahn, E.J.; Paek, K.Y. Application of an Airlift Bioreactor System for the Production of Adventitious Root Biomass and Caffeic Acid Derivatives of Echinacea purpurea. Biotechnol. Bioprocess Eng. 2009, 14, 91–98. [Google Scholar] [CrossRef]
- Cech, N.B.; Eleazer, M.S.; Shoffner, L.T.; Crosswhite, M.R.; Davis, A.C.; Mortenson, A.M. High Performance Liquid Chromatography/Electrospray Ionization Mass Spectrometry for Simultaneous Analysis of Alkamides and Caffeic Acid Derivatives from Echinacea purpurea Extracts. J. Chromatogr. A 2006, 1103, 219–228. [Google Scholar] [CrossRef] [Green Version]
- Freier, D.O.; Wright, K.; Klein, K.; Voll, D.; Dabiri, K.; Cosulich, K.; George, R. Enhancement of the Humoral Immune Response by Echinacea purpurea in Female Swiss Mice. Immunopharmacol. Immunotoxicol. 2003, 25, 551–560. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, S.J.; Hume, D.A. Homeostasis in the Mononuclear Phagocyte System. Trends Immunol. 2014, 35, 358–367. [Google Scholar] [CrossRef] [PubMed]
- Kolaczkowska, E.; Kubes, P. Neutrophil Recruitment and Function in Health and Inflammation. Nat. Rev. Immunol. 2013, 13, 159–175. [Google Scholar] [CrossRef]
- Italiani, P.; Boraschi, D. From Monocytes to M1/M2 Macrophages: Phenotypical vs. Functional Differentiation. Front. Immunol. 2014, 5, 514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mills, C.D.; Kincaid, K.; Alt, J.M.; Heilman, M.J.; Hill, A.M. M-1/M-2 Macrophages and the Th1/Th2 Paradigm. J. Immunol. Baltim. Md 1950 2000, 164, 6166–6173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, V. Macrophages: The Potent Immunoregulatory Innate Immune Cells. Macrophage Act. Biol. Dis. 2019. [Google Scholar] [CrossRef] [Green Version]
- Poltavets, A.S.; Vishnyakova, P.A.; Elchaninov, A.V.; Sukhikh, G.T.; Fatkhudinov, T.K. Macrophage Modification Strategies for Efficient Cell Therapy. Cells 2020, 9, 1535. [Google Scholar] [CrossRef] [PubMed]
- Imig, J.D.; Ryan, M.J. Immune and Inflammatory Role in Renal Disease. Compr. Physiol. 2013, 3, 957–976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tritarelli, A.; Oricchio, E.; Ciciarello, M.; Mangiacasale, R.; Palena, A.; Lavia, P.; Soddu, S.; Cundari, E. P53 Localization at Centrosomes during Mitosis and Postmitotic Checkpoint Are ATM-Dependent and Require Serine 15 Phosphorylation. Mol. Biol. Cell 2004, 15, 3751–3757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kotsias, F.; Cebrian, I.; Alloatti, A. Antigen Processing and Presentation. Int. Rev. Cell Mol. Biol. 2019, 348, 69–121. [Google Scholar] [CrossRef] [PubMed]
- Chaplin, D.D. Overview of the Immune Response. J. Allergy Clin. Immunol. 2010, 125, S3–S23. [Google Scholar] [CrossRef] [PubMed]
- Netea, M.G.; Schlitzer, A.; Placek, K.; Joosten, L.A.B.; Schultze, J.L. Innate and Adaptive Immune Memory: An Evolutionary Continuum in the Host’s Response to Pathogens. Cell Host Microbe 2019, 25, 13–26. [Google Scholar] [CrossRef] [Green Version]
- Mebius, R.E.; Kraal, G. Structure and Function of the Spleen. Nat. Rev. Immunol. 2005, 5, 606–616. [Google Scholar] [CrossRef] [PubMed]
- Matthias, A.; Banbury, L.; Bone, K.M.; Leach, D.N.; Lehmann, R.P. Echinacea Alkylamides Modulate Induced Immune Responses in T-Cells. Fitoterapia 2008, 79, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, T.A.; Mariano, V.S.; Sardinha-Silva, A.; De Souza, M.A.; Mineo, T.W.P.; Roque-Barreira, M.C. IL-17 Induction by ArtinM Is Due to Stimulation of IL-23 and IL-1 Release and/or Interaction with CD3 in CD4+T Cells. PLoS ONE 2016, 11, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamma, S.M.L.; Kalyanaraman, V.S.; Pahwa, S.; Dominguez, P.; Modesto, R.R. The Lectin Jacalin Induces Phosphorylation of ERK and JNK in CD4+ T Cells. J. Leukoc. Biol. 2003, 73, 682–688. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, T.A.; de Souza, M.A.; Cecílio, N.T.; Roque-Barreira, M.C. ristina Activation of Spleen Cells by ArtinM May Account for Its Immunomodulatory Properties. Cell Tissue Res. 2014, 357, 719–730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ditamo, Y.; Rupil, L.L.; Sendra, V.G.; Nores, G.A.; Roth, G.A.; Irazoqui, F.J. In Vivo Immunomodulatory Effect of the Lectin from Edible Mushroom Agaricus Bisporus. Food Funct. 2016, 7, 262–269. [Google Scholar] [CrossRef] [PubMed]
- Brown, P.N.; Columbia, B.; Ave, W.; Chan, M.; Betz, J.M. Determination of Major Phenolic Compounds in Echinacea Spp. Raw Materials and Finished Products by High-Performance Liquid Chromatography with Ultraviolet Detection: Single- Laboratory Validation Matrix Extension. J. AOAC Int. 2013, 94, 1400–1410. [Google Scholar] [CrossRef] [Green Version]
- Council of Europe; European Pharmacopoeia Commission. Dry residue of extracts (2.8.17.). In European Pharmacopoeia, 9.0; Council of Europe: Strasbourg, France, 2018; p. 289. [Google Scholar]
- Smalinskiene, A.; Lesauskaite, V.; Ryselis, S.; Abdrakhmanov, O.; Kregzdyte, R.; Sadauskiene, I.; Ivanov, L.; Savickiene, N.; Zitkevičius, V.; Savickas, A. Assessment of the Effect of Echinacea purpurea (L.) Moench on Apoptotic and Mitotic Activity of Liver Cells during Intoxication by Cadmium. Ann. N. Y. Acad. Sci. 2007, 1095, 574–584. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nr. | RT | Compound | Linear Range, μg/mL | Regression Equation | R2 | LOD, μg/mL | LOQ, μg/mL | Quantity, μg/mL | Quantity, μg/g, in Fresh Root |
|---|---|---|---|---|---|---|---|---|---|
| 1. | 4.69 | Caftaric acid | 1.56–50.00 | y = 357,000x – 574,000 | 0.997 | 0.158 | 0.527 | 1.95 ± 0.10 | 0.18 ± 0.01 |
| 2. | 6.83 | Chlorogenic acid | 0.646 | 2.154 | 1.75 ± 0.09 | 3.16 ± 0.13 | |||
| 3. | 7.712 | Caffeic acid | 0.025 | 0.085 | 3.78 ± 0.23 | 6.80 ± 0.34 | |||
| 4. | 9.719 | Cichoric acid | 0.031 | 0.104 | 3.47 ± 0.14 | 6.24 ± 0.25 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Balčiūnaitė-Murzienė, G.; Miknienė, Z.; Ragažinskienė, O.; Juodžiukynienė, N.; Savickas, A.; Savickienė, N.; Pangonytė, D. Echinacea purpurea L. (Moench) Hemagglutinin Effect on Immune Response In Vivo. Plants 2021, 10, 936. https://doi.org/10.3390/plants10050936
Balčiūnaitė-Murzienė G, Miknienė Z, Ragažinskienė O, Juodžiukynienė N, Savickas A, Savickienė N, Pangonytė D. Echinacea purpurea L. (Moench) Hemagglutinin Effect on Immune Response In Vivo. Plants. 2021; 10(5):936. https://doi.org/10.3390/plants10050936
Chicago/Turabian StyleBalčiūnaitė-Murzienė, Gabrielė, Zoja Miknienė, Ona Ragažinskienė, Nomeda Juodžiukynienė, Arūnas Savickas, Nijolė Savickienė, and Dalia Pangonytė. 2021. "Echinacea purpurea L. (Moench) Hemagglutinin Effect on Immune Response In Vivo" Plants 10, no. 5: 936. https://doi.org/10.3390/plants10050936