
Harnessing Bacterial Endophytes for Promotion of Plant Growth and Biotechnological Applications: An Overview

 , , ,
, , ,  ,
,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Types of Bacterial Endophytes
3. Plant–Bacterial Endophyte Interactions
4. Applications of Bacterial Endophytes
4.1. Agricultural Applications
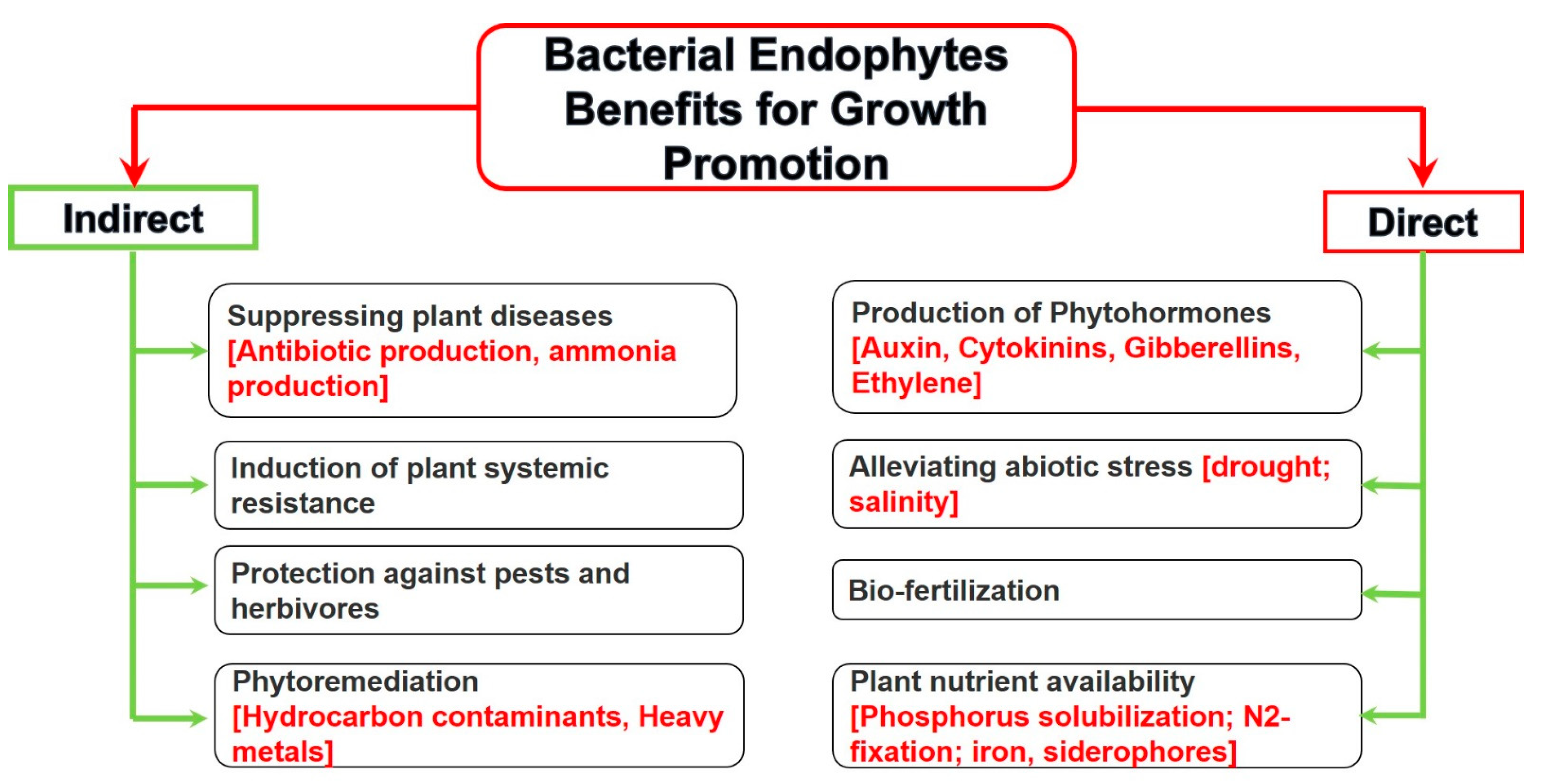
4.1.1. Plant Growth Promotion
Direct Plant Growth Promotion
- A.
- Phytohormone production
- B.
- Biofertilization
- a.
- Nitrogen fixation
- b.
- Phosphate Solubilization
- c.
- Siderophore Production
Indirect Plant Growth Promotion
- A.
- Stress Tolerance
- B.
- Endophyte-Based Phytoremediation
- C.
- Disease Control
- D.
- Competition for Space and Nutrients
- a.
- Antibiosis
- b.
- Parasitism
- c.
- Induced Systemic Resistance (ISR)
4.2. Biotechnological Applications
4.2.1. Production of Bioactive Metabolites for Agricultural and Medical Applications
Pharmaceutical Applications
4.2.2. Industrial Applications
4.2.3. Nano Biotechnology
5. Conclusions and Future Prospective
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tidke, S.A.; Kiran, S.; Giridhar, P.; Gokare, R.A. Current Understanding and Future Perspectives of Endophytic Microbes vis-a-vis Production of Secondary Metabolites. In Endophytes and Secondary Metabolites; Jha, S., Ed.; Springer International Publishing: Cham, Switzerland, 2018; pp. 1–16. [Google Scholar]
- Fouda, A.H.; Hassan, S.E.-D.; Eid, A.M.; Ewais, E.E.-D. Biotechnological applications of fungal endophytes associated with medicinal plant Asclepias sinaica (Bioss.). Ann. Agric. Sci. 2015, 60, 95–104. [Google Scholar] [CrossRef] [Green Version]
- Strobel, G. The Emergence of Endophytic Microbes and Their Biological Promise. J. Fungi 2018, 4, 57. [Google Scholar] [CrossRef] [Green Version]
- Khalil, A.M.A.; Hassan, S.E.-D.; Alsharif, S.M.; Eid, A.M.; Ewais, E.E.-D.; Azab, E.; Gobouri, A.A.; Elkelish, A.; Fouda, A. Isolation and Characterization of Fungal Endophytes Isolated from Medicinal Plant Ephedra pachyclada as Plant Growth-Promoting. Biomolecules 2021, 11, 140. [Google Scholar] [CrossRef] [PubMed]
- Eid, A.M.; Salim, S.S.; Hassan, S.E.-D.; Ismail, M.A.; Fouda, A. Role of Endophytes in Plant Health and Abiotic Stress Management. In Microbiome in Plant Health and Disease: Challenges and Opportunities; Kumar, V., Prasad, R., Kumar, M., Choudhary, D.K., Eds.; Springer: Singapore, 2019; pp. 119–144. [Google Scholar]
- Golinska, P.; Wypij, M.; Agarkar, G.; Rathod, D.; Dahm, H.; Rai, M. Endophytic actinobacteria of medicinal plants: Diversity and bioactivity. Antonie Van Leeuwenhoek 2015, 108, 267–289. [Google Scholar] [CrossRef] [Green Version]
- Sun, H.; He, Y.; Xiao, Q.; Ye, R.; Tian, Y. Isolation, characterization, and antimicrobial activity of endophytic bacteria from Polygonum cuspidatum. Afr. J. Microbiol. Res. 2013, 7, 1496–1504. [Google Scholar] [CrossRef] [Green Version]
- Hollants, J.; Leroux, O.; Leliaert, F.; Decleyre, H.; De Clerck, O.; Willems, A. Who Is in There? Exploration of Endophytic Bacteria within the Siphonous Green Seaweed Bryopsis (Bryopsidales, Chlorophyta). PLoS ONE 2011, 6, e26458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rangjaroen, C.; Sungthong, R.; Rerkasem, B.; Teaumroong, N.; Noisangiam, R.; Lumyong, S. Untapped Endophytic Colonization and Plant Growth-Promoting Potential of the Genus Novosphingobium to Optimize Rice Cultivation. Microbes Environ. 2017, 32, 84–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castanheira, N.L.; Dourado, A.C.; Pais, I.; Semedo, J.; Scotti-Campos, P.; Borges, N.; Carvalho, G.; Crespo, M.T.B.; Fareleira, P. Colonization and beneficial effects on annual ryegrass by mixed inoculation with plant growth promoting bacteria. Microbiol. Res. 2017, 198, 47–55. [Google Scholar] [CrossRef]
- Mercado-Blanco, J.; Lugtenberg, B.J.J. Biotechnological Applications of Bacterial Endophytes. Curr. Biotechnol. 2014, 3, 60–75. [Google Scholar] [CrossRef] [Green Version]
- Hardoim, P.R.; van Overbeek, L.S.; Elsas, J.D.v. Properties of bacterial endophytes and their proposed role in plant growth. Trends Microbiol. 2008, 16, 463–471. [Google Scholar] [CrossRef]
- Abreu-Tarazi, M.F.; Navarrete, A.A.; Andreote, F.D.; Almeida, C.V.; Tsai, S.M.; Almeida, M. Endophytic bacteria in long-term in vitro cultivated “axenic” pineapple microplants revealed by PCR–DGGE. World J. Microbiol. Biotechnol. 2010, 26, 555–560. [Google Scholar] [CrossRef]
- Li, J.; Zheng, B.; Hu, R.; Liu, Y.; Jing, Y.; Xiao, Y.; Sun, M.; Chen, W.; Zhou, Q. Pseudomonas species isolated from tobacco seed promote root growth and reduce lead contents in Nicotiana tobacum K326. Can J Microbiol 2019, 65, 214–223. [Google Scholar] [CrossRef] [PubMed]
- Annapurna, K.; Govindasamy, V.; Sharma, M.; Ghosh, A.; Chikara, S.K. Whole genome shotgun sequence of Bacillus paralicheniformis strain KMS 80, a rhizobacterial endophyte isolated from rice (Oryza sativa L.). 3 Biotech 2018, 8, 223. [Google Scholar] [CrossRef]
- Zhu, Y.; She, X. Evaluation of the plant-growth-promoting abilities of endophytic bacteria from the psammophyte Ammodendron bifolium. Can. J. Microbiol. 2018, 64, 253–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mareque, C.; da Silva, T.F.; Vollú, R.E.; Beracochea, M.; Seldin, L.; Battistoni, F. The Endophytic Bacterial Microbiota Associated with Sweet Sorghum (Sorghum bicolor) Is Modulated by the Application of Chemical N Fertilizer to the Field. Int. J. Genom. 2018, 2018, 7403670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez-Escobedo, R.; Briones-Roblero, C.I.; Pineda-Mendoza, R.M.; Rivera-Orduña, F.N.; Zúñiga, G. Bacteriome from Pinus arizonica and P. durangensis: Diversity, Comparison of Assemblages, and Overlapping Degree with the Gut Bacterial Community of a Bark Beetle That Kills Pines. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef]
- Nguyen, H.Q.; Vu, N.T.-H.; Chu, H.H.; Chu, S.K.; Hoang, H.; Tran, T.T.; Nguyen, C.; Dinh, L.T.-M.; Trinh, H.T.-T.; Phi, T.Q. Draft Genome Sequence of Streptomyces cavourensis YBQ59, an Endophytic Producer of Antibiotics Bafilomycin D, Nonactic Acid, Prelactone B, and 5,11-Epoxy-10-Cadinanol. Microbiol. Resour. Announc. 2018, 7, e01056-18. [Google Scholar] [CrossRef] [Green Version]
- Asaf, S.; Khan, A.L.; Khan, M.A.; Al-Harrasi, A.; Lee, I.-J. Complete genome sequencing and analysis of endophytic Sphingomonas sp. LK11 and its potential in plant growth. 3 Biotech 2018, 8, 389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haidar, B.; Ferdous, M.; Fatema, B.; Ferdous, A.S.; Islam, M.R.; Khan, H. Population diversity of bacterial endophytes from jute (Corchorus olitorius) and evaluation of their potential role as bioinoculants. Microbiol. Res. 2018, 208, 43–53. [Google Scholar] [CrossRef]
- Verma, S.K.; White, J.F. Indigenous endophytic seed bacteria promote seedling development and defend against fungal disease in browntop millet (Urochloa ramosa L.). J. Appl. Microbiol. 2018, 124, 764–778. [Google Scholar] [CrossRef]
- de Almeida Lopes, K.B.; Carpentieri-Pipolo, V.; Fira, D.; Balatti, P.A.; López, S.M.Y.; Oro, T.H.; Stefani Pagliosa, E.; Degrassi, G. Screening of bacterial endophytes as potential biocontrol agents against soybean diseases. J. Appl. Microbiol. 2018, 125, 1466–1481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bind, M.; Nema, S. Isolation and Molecular Characterization of Endophytic Bacteria from Pigeon Pea along with Antimicrobial Evaluation against Fusarium udum. Appl. Microbiol. Open Access 2019, 5, 163. [Google Scholar] [CrossRef]
- Soldan, R.; Mapelli, F.; Crotti, E.; Schnell, S.; Daffonchio, D.; Marasco, R.; Fusi, M.; Borin, S.; Cardinale, M. Bacterial endophytes of mangrove propagules elicit early establishment of the natural host and promote growth of cereal crops under salt stress. Microbiol. Res. 2019, 223–225, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Brígido, C.; Singh, S.; Menéndez, E.; Tavares, M.J.; Glick, B.R.; Félix, M.d.R.; Oliveira, S.; Carvalho, M. Diversity and Functionality of Culturable Endophytic Bacterial Communities in Chickpea Plants. Plants 2019, 8, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Wu, Y.D.; Chong, X.Y.; Xin, Q.H.; Wang, D.X.; Bian, K. Seed-borne endophytic Bacillus velezensis LHSB1 mediate the biocontrol of peanut stem rot caused by Sclerotium rolfsii. J. Appl. Microbiol. 2020, 128, 803–813. [Google Scholar] [CrossRef]
- Aquino, J.P.A.D.; Macedo Junior, F.B.D.; Antunes, J.E.L.; Figueiredo, M.D.V.B.; Alcântara Neto, F.D.; Araujo, A.S.F.D. Plant growth-promoting endophytic bacteria on maize and sorghum. Pesqui. Agropecuária Trop. 2019, 49. [Google Scholar] [CrossRef]
- Woźniak, M.; Gałązka, A.; Tyśkiewicz, R.; Jaroszuk-Ściseł, J. Endophytic Bacteria Potentially Promote Plant Growth by Synthesizing Different Metabolites and their Phenotypic/Physiological Profiles in the Biolog GEN III MicroPlateTM Test. Int. J. Mol. Sci. 2019, 20, 5283. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Li, P.; Meng, D.; Gu, Y.; Zheng, Z.; Yin, H.; Zhou, Q.; Li, J. Isolation, characterization and inoculation of Cd tolerant rice endophytes and their impacts on rice under Cd contaminated environment. Environ. Pollut. 2020, 260, 113990. [Google Scholar] [CrossRef]
- Borah, A.; Thakur, D. Phylogenetic and Functional Characterization of Culturable Endophytic Actinobacteria Associated With Camellia spp. for Growth Promotion in Commercial Tea Cultivars. Front. Microbiol. 2020, 11, 318. [Google Scholar] [CrossRef] [Green Version]
- Herrera, H.; Sanhueza, T.; Novotná, A.; Charles, T.C.; Arriagada, C. Isolation and Identification of Endophytic Bacteria from Mycorrhizal Tissues of Terrestrial Orchids from Southern Chile. Diversity 2020, 12, 55. [Google Scholar] [CrossRef] [Green Version]
- Gamalero, E.; Favale, N.; Bona, E.; Novello, G.; Cesaro, P.; Massa, N.; Glick, B.R.; Orozco-Mosqueda, M.d.C.; Berta, G.; Lingua, G. Screening of Bacterial Endophytes Able to Promote Plant Growth and Increase Salinity Tolerance. Appl. Sci. 2020, 10, 5767. [Google Scholar] [CrossRef]
- Khan, M.S.; Gao, J.; Chen, X.; Zhang, M.; Yang, F.; Du, Y.; Moe, T.S.; Munir, I.; Xue, J.; Zhang, X. Isolation and Characterization of Plant Growth-Promoting Endophytic Bacteria Paenibacillus polymyxa SK1 from Lilium lancifolium. Biomed Res. Int. 2020, 2020, 8650957. [Google Scholar] [CrossRef] [Green Version]
- Khaskheli, M.A.; Wu, L.; Chen, G.; Chen, L.; Hussain, S.; Song, D.; Liu, S.; Feng, G. Isolation and Characterization of Root-Associated Bacterial Endophytes and Their Biocontrol Potential against Major Fungal Phytopathogens of Rice (Oryza sativa L.). Pathogens 2020, 9, 172. [Google Scholar] [CrossRef]
- ALKahtani, M.D.F.; Fouda, A.; Attia, K.A.; Al-Otaibi, F.; Eid, A.M.; Ewais, E.E.-D.; Hijri, M.; St-Arnaud, M.; Hassan, S.E.-D.; Khan, N.; et al. Isolation and Characterization of Plant Growth Promoting Endophytic Bacteria from Desert Plants and Their Application as Bioinoculants for Sustainable Agriculture. Agronomy 2020, 10, 1325. [Google Scholar] [CrossRef]
- Schirawski, J.; Perlin, M.H. Plant–Microbe Interaction 2017—The Good, the Bad and the Diverse. Int. J. Mol. Sci. 2018, 19, 1374. [Google Scholar] [CrossRef] [Green Version]
- Hassan, S.E.-D. Plant growth-promoting activities for bacterial and fungal endophytes isolated from medicinal plant of Teucrium polium L. J Adv Res 2017, 8, 687–695. [Google Scholar] [CrossRef]
- Doty, S.L. Symbiotic Plant-Bacterial Endospheric Interactions. Microorganisms 2018, 6, 28. [Google Scholar] [CrossRef] [Green Version]
- Afzal, I.; Shinwari, Z.K.; Sikandar, S.; Shahzad, S. Plant beneficial endophytic bacteria: Mechanisms, diversity, host range and genetic determinants. Microbiol. Res. 2019, 221, 36–49. [Google Scholar] [CrossRef]
- Nair, D.N.; Padmavathy, S. Impact of Endophytic Microorganisms on Plants, Environment and Humans. Sci. World J. 2014, 2014, 250693. [Google Scholar] [CrossRef] [Green Version]
- Rosenblueth, M.; Martínez-Romero, E. Bacterial Endophytes and Their Interactions with Hosts. Mol. Plant-Microbe Interact. 2006, 19, 827–837. [Google Scholar] [CrossRef] [Green Version]
- Kandel, S.L.; Joubert, P.M.; Doty, S.L. Bacterial Endophyte Colonization and Distribution within Plants. Microorganisms 2017, 5. [Google Scholar] [CrossRef] [Green Version]
- Balsanelli, E.; Serrato, R.V.; De Baura, V.A.; Sassaki, G.; Yates, M.G.; Rigo, L.U.; Pedrosa, F.O.; De Souza, E.M.; Monteiro, R.A. Herbaspirillum seropedicae rfbB and rfbC genes are required for maize colonization. Environ. Microbiol. 2010, 12, 2233–2244. [Google Scholar] [PubMed]
- Reinhold-Hurek, B.; Maes, T.; Gemmer, S.; Van Montagu, M.; Hurek, T. An Endoglucanase Is Involved in Infection of Rice Roots by the Not-Cellulose-Metabolizing Endophyte Azoarcus Sp. Strain BH72. Mol. Plant-Microbe Interact. 2006, 19, 181–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fouda, A.; Abdel-Maksoud, G.; Saad, H.A.; Gobouri, A.A.; Mohammedsaleh, Z.M.; El-Sadany, M.A. The efficacy of silver nitrate (AgNO3) as a coating agentto protect paper against high deteriorating microbes. Catalysts 2021, 11, 310. [Google Scholar] [CrossRef]
- Suárez-Moreno, Z.R.; Devescovi, G.; Myers, M.; Hallack, L.; Mendonça-Previato, L.; Caballero-Mellado, J.; Venturi, V. Commonalities and Differences in Regulation of N-Acyl Homoserine Lactone Quorum Sensing in the Beneficial Plant-Associated Burkholderia Species Cluster. Appl. Environ. Microbiol. 2010, 76, 4302. [Google Scholar] [CrossRef] [Green Version]
- Compant, S.; Reiter, B.; Sessitsch, A.; Nowak, J.; Clément, C.; Ait Barka, E. Endophytic Colonization of Vitis vinifera L. by Plant Growth-Promoting Bacterium Burkholderia sp. Strain PsJN. Appl. Environ. Microbiol. 2005, 71, 1685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prieto, P.; Schilirò, E.; Maldonado-González, M.M.; Valderrama, R.; Barroso-Albarracín, J.B.; Mercado-Blanco, J. Root hairs play a key role in the endophytic colonization of olive roots by Pseudomonas spp. with biocontrol activity. Microb. Ecol. 2011, 62, 435–445. [Google Scholar] [CrossRef] [Green Version]
- White, J.F., Jr.; Torres, M.S.; Somu, M.P.; Johnson, H.; Irizarry, I.; Chen, Q.; Zhang, N.; Walsh, E.; Tadych, M.; Bergen, M. Hydrogen peroxide staining to visualize intracellular bacterial infections of seedling root cells. Microsc. Res. Tech. 2014, 77, 566–573. [Google Scholar] [CrossRef]
- Paungfoo-Lonhienne, C.; Rentsch, D.; Robatzek, S.; Webb, R.I.; Sagulenko, E.; Näsholm, T.; Schmidt, S.; Lonhienne, T.G.A. Turning the Table: Plants Consume Microbes as a Source of Nutrients. PLoS ONE 2010, 5, e11915. [Google Scholar] [CrossRef] [Green Version]
- Hardoim, P.R.; van Overbeek, L.S.; Berg, G.; Pirttilä, A.M.; Compant, S.; Campisano, A.; Döring, M.; Sessitsch, A. The Hidden World within Plants: Ecological and Evolutionary Considerations for Defining Functioning of Microbial Endophytes. Microbiol. Mol. Biol. Rev. 2015, 79, 293. [Google Scholar] [CrossRef] [Green Version]
- Kandel, S.L.; Herschberger, N.; Kim, S.H.; Doty, S.L. Diazotrophic Endophytes of Poplar and Willow for Growth Promotion of Rice Plants in Nitrogen-Limited Conditions. Crop Sci. 2015, 55, 1765–1772. [Google Scholar] [CrossRef] [Green Version]
- Compant, S.; Duffy, B.; Nowak, J.; Clément, C.; Barka, E.A. Use of plant growth-promoting bacteria for biocontrol of plant diseases: Principles, mechanisms of action, and future prospects. Appl. Environ. Microbiol. 2005, 71, 4951–4959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitter, B.; Pfaffenbichler, N.; Flavell, R.; Compant, S.; Antonielli, L.; Petric, A.; Berninger, T.; Naveed, M.; Sheibani-Tezerji, R.; von Maltzahn, G.; et al. A New Approach to Modify Plant Microbiomes and Traits by Introducing Beneficial Bacteria at Flowering into Progeny Seeds. Front. Microbiol. 2017, 8, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glassner, H.; Zchori-Fein, E.; Yaron, S.; Sessitsch, A.; Sauer, U.; Compant, S. Bacterial niches inside seeds of Cucumis melo L. Plant Soil 2018, 422, 101–113. [Google Scholar] [CrossRef]
- Koskimäki, J.J.; Pirttilä, A.M.; Ihantola, E.-L.; Halonen, O.; Frank, A.C. The Intracellular Scots Pine Shoot Symbiont Methylobacterium extorquens DSM13060 Aggregates around the Host Nucleus and Encodes Eukaryote-Like Proteins. mBio 2015, 6, e00039-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, P.; Reddy, K.M. Microscopic elucidation of abundant endophytic bacteria colonizing the cell wall–plasma membrane peri-space in the shoot-tip tissue of banana. AOB Plants 2013, 5. [Google Scholar] [CrossRef] [Green Version]
- Santoyo, G.; Moreno-Hagelsieb, G.; del Carmen Orozco-Mosqueda, M.; Glick, B.R. Plant growth-promoting bacterial endophytes. Microbiol. Res. 2016, 183, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Damam, M.; Kaloori, K.; Gaddam, B.; Kausar, R. Plant growth promoting substances (phytohormones) produced by rhizobacterial strains isolated from the rhizosphere of medicinal plants. Int. J. Pharm. Sci. Rev. Res. 2016, 37, 130–136. [Google Scholar]
- Selim, M.T.; Salem, S.S.; Mohamed, A.A.; El-Gamal, M.S.; Awad, M.F.; Fouda, A. Biological treatment of real textile effulent using Aspergillus flavus and Fusarium oxysorium and their consortium along with the evaluation of their phytotoxicity. J. Fungi. 2021, 7, 193. [Google Scholar] [CrossRef]
- Shahzad, R.; Waqas, M.; Khan, A.L.; Asaf, S.; Khan, M.A.; Kang, S.-M.; Yun, B.-W.; Lee, I.-J. Seed-borne endophytic Bacillus amyloliquefaciens RWL-1 produces gibberellins and regulates endogenous phytohormones of Oryza sativa. Plant Physiol. Biochem. 2016, 106, 236–243. [Google Scholar] [CrossRef]
- Brader, G.; Compant, S.; Mitter, B.; Trognitz, F.; Sessitsch, A. Metabolic potential of endophytic bacteria. Curr. Opin. Biotechnol. 2014, 27, 30–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reinhold-Hurek, B.; Hurek, T. Living inside plants: Bacterial endophytes. Curr. Opin. Plant Biol. 2011, 14, 435–443. [Google Scholar] [CrossRef] [PubMed]
- Asgher, M.; Khan, M.I.R.; Anjum, N.A.; Khan, N.A. Minimising toxicity of cadmium in plants—Role of plant growth regulators. Protoplasma 2015, 252, 399–413. [Google Scholar] [CrossRef]
- Spaepen, S.; Vanderleyden, J. Auxin and plant-microbe interactions. Cold Spring Harb. Perspect. Biol. 2011, 3, a001438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ismail, M.A.; Amin, M.A.; Eid, A.M.; Hassan, S.E.-D.; Mahgoub, H.A.M.; Lashin, I.; Abdelwahab, A.T.; Azab, E.; Gobouri, A.A.; ElKelish, A.; et al. Comparative Study between Exogenously Applied Plant Growth Hormones versus Metabolites of Microbial Endophytes as Plant Growth-Promoting for Phaseolus vulgaris L. Cells 2021, 10, 1059. [Google Scholar] [CrossRef]
- Rashid, S.; Charles, T.C.; Glick, B.R. Isolation and characterization of new plant growth-promoting bacterial endophytes. Appl. Soil Ecol. 2012, 61, 217–224. [Google Scholar] [CrossRef]
- Duca, D.; Lorv, J.; Patten, C.L.; Rose, D.; Glick, B.R. Indole-3-acetic acid in plant–microbe interactions. Antonie Van Leeuwenhoek 2014, 106, 85–125. [Google Scholar] [CrossRef]
- Apine, O.; Jadhav, J. Optimization of medium for indole-3-acetic acid production using Pantoea agglomerans strain PVM. J. Appl. Microbiol. 2011, 110, 1235–1244. [Google Scholar] [CrossRef]
- Costacurta, A.; Vanderleyden, J. Synthesis of phytohormones by plant-associated bacteria. Crit. Rev. Microbiol. 1995, 21, 1–18. [Google Scholar] [CrossRef]
- Fouda, A.; Eid, A.M.; Elsaied, A.; El-Belely, E.F.; Barghoth, M.G.; Azab, E.; Gobouri, A.A.; Hassan, S.E.-D. Plant Growth-Promoting Endophytic Bacterial Community Inhabiting the Leaves of Pulicaria incisa (Lam.) DC Inherent to Arid Regions. Plants 2021, 10, 76. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Wang, X.; Li, Y. Increased plant growth and copper uptake of host and non-host plants by metal-resistant and plant growth-promoting endophytic bacteria. Int. J. Phytoremediation 2016, 18, 494–501. [Google Scholar] [CrossRef]
- Sun, Y.; Cheng, Z.; Glick, B.R. The presence of a 1-aminocyclopropane-1-carboxylate (ACC) deaminase deletion mutation alters the physiology of the endophytic plant growth-promoting bacterium Burkholderia phytofirmans PsJN. FEMS Microbiol. Lett. 2009, 296, 131–136. [Google Scholar] [CrossRef] [Green Version]
- Albermann, S.; Linnemannstöns, P.; Tudzynski, B. Strategies for strain improvement in Fusarium fujikuroi: Overexpression and localization of key enzymes of the isoprenoid pathway and their impact on gibberellin biosynthesis. Appl. Microbiol. Biotechnol. 2013, 97, 2979–2995. [Google Scholar] [CrossRef]
- Rana, K.L.; Kour, D.; Kaur, T.; Devi, R.; Yadav, A.N.; Yadav, N.; Dhaliwal, H.S.; Saxena, A.K. Endophytic microbes: Biodiversity, plant growth-promoting mechanisms and potential applications for agricultural sustainability. Antonie Van Leeuwenhoek 2020, 113, 1075–1107. [Google Scholar] [CrossRef] [PubMed]
- Kieber, J.J. Tribute to Folke Skoog: Recent Advances in our Understanding of Cytokinin Biology. J. Plant Growth Regul. 2002, 21, 1–2. [Google Scholar] [CrossRef]
- Cohen, A.C.; Travaglia, C.N.; Bottini, R.; Piccoli, P.N. Participation of abscisic acid and gibberellins produced by endophytic Azospirillum in the alleviation of drought effects in maize. Botany 2009, 87, 455–462. [Google Scholar] [CrossRef]
- Bhore, S.J.; Ravichantar, N.; Loh, C.Y. Screening of endophytic bacteria isolated from leaves of Sambung nyawa [Gynura procumbens (Lour.) Merr.] for cytokinin-like compounds. Bioinformation 2010, 5, 191–197. [Google Scholar] [CrossRef] [Green Version]
- Arkhipova, T.; Veselov, S.; Melentiev, A.; Martynenko, E.; Kudoyarova, G. Ability of bacterium Bacillus subtilis to produce cytokinins and to influence the growth and endogenous hormone content of lettuce plants. Plant Soil 2005, 272, 201–209. [Google Scholar] [CrossRef]
- Peña-Yam, L.P.; Ruíz-Sánchez, E.; Barboza-Corona, J.E.; Reyes-Ramírez, A. Isolation of mexican Bacillus species and their effects in promoting growth of chili pepper (Capsicum annuum L. cv Jalapeño). Indian J. Microbiol. 2016, 56, 375–378. [Google Scholar] [CrossRef] [Green Version]
- Gowtham, H.; Murali, M.; Singh, S.B.; Lakshmeesha, T.; Murthy, K.N.; Amruthesh, K.; Niranjana, S. Plant growth promoting rhizobacteria-Bacillus amyloliquefaciens improves plant growth and induces resistance in chilli against anthracnose disease. Biol. Control 2018, 126, 209–217. [Google Scholar] [CrossRef]
- Sabaté, D.C.; Brandan, C.P.; Petroselli, G.; Erra-Balsells, R.; Audisio, M.C. Biocontrol of Sclerotinia sclerotiorum (Lib.) de Bary on common bean by native lipopeptide-producer Bacillus strains. Microbiol. Res. 2018, 211, 21–30. [Google Scholar] [CrossRef]
- de Melo Pereira, G.V.; Magalhães, K.T.; Lorenzetii, E.R.; Souza, T.P.; Schwan, R.F. A multiphasic approach for the identification of endophytic bacterial in strawberry fruit and their potential for plant growth promotion. Microb. Ecol. 2012, 63, 405–417. [Google Scholar] [CrossRef]
- Kumawat, K.C.; Sharma, P.; Sirari, A.; Singh, I.; Gill, B.S.; Singh, U.; Saharan, K. Synergism of Pseudomonas aeruginosa (LSE-2) nodule endophyte with Bradyrhizobium sp. (LSBR-3) for improving plant growth, nutrient acquisition and soil health in soybean. World J. Microbiol. Biotechnol. 2019, 35, 47. [Google Scholar] [CrossRef] [PubMed]
- Sandhya, V. Endophytes from maize with plant growth promotion and biocontrol activity under drought stress. Russ. Agric. Sci. 2017, 43, 22–34. [Google Scholar] [CrossRef]
- Rana, K.L.; Kour, D.; Kaur, T.; Sheikh, I.; Yadav, A.N.; Kumar, V.; Suman, A.; Dhaliwal, H.S. Endophytic Microbes from Diverse Wheat Genotypes and Their Potential Biotechnological Applications in Plant Growth Promotion and Nutrient Uptake. Proc. Natl. Acad. Sci. India Sect. B: Biol. Sci. 2020, 90, 969–979. [Google Scholar] [CrossRef]
- Singh, D.; Geat, N.; Rajawat, M.V.S.; Mahajan, M.M.; Prasanna, R.; Singh, S.; Kaushik, R.; Singh, R.N.; Kumar, K.; Saxena, A.K. Deciphering the Mechanisms of Endophyte-Mediated Biofortification of Fe and Zn in Wheat. J. Plant Growth Regul. 2018, 37, 174–182. [Google Scholar] [CrossRef]
- Zhao, L.; Xu, Y.; Lai, X. Antagonistic endophytic bacteria associated with nodules of soybean (Glycine max L.) and plant growth-promoting properties. Braz. J. Microbiol. 2018, 49, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Spaink, H.P.; Kondorosi, A.; Hooykaas, P.J. The Rhizobiaceae: Molecular Biology of Model Plant-Associated Bacteria; Springer Science & Business Media: Dordrecht, The Netherlands, 2012. [Google Scholar]
- Shridhar, B.S. Nitrogen fixing microorganisms. Int. J. Microbiol. Res. (IJMR) 2012, 3, 46–52. [Google Scholar]
- Ladha, J.K.; Reddy, P.M. Nitrogen fixation in rice systems: State of knowledge and future prospects. Plant Soil 2003, 252, 151–167. [Google Scholar] [CrossRef]
- Bhattacharjee, R.B.; Singh, A.; Mukhopadhyay, S. Use of nitrogen-fixing bacteria as biofertiliser for non-legumes: Prospects and challenges. Appl. Microbiol. Biotechnol. 2008, 80, 199–209. [Google Scholar] [CrossRef]
- Santi, C.; Bogusz, D.; Franche, C. Biological nitrogen fixation in non-legume plants. Ann. Bot. 2013, 111, 743–767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Triplett, E.W. Diazotrophic endophytes: Progress and prospects for nitrogen fixation in monocots. Plant Soil 1996, 186, 29–38. [Google Scholar] [CrossRef]
- Hurek, T.; Reinhold-Hurek, B. Azoarcus sp. strain BH72 as a model for nitrogen-fixing grass endophytes. J. Biotechnol. 2003, 106, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Govindarajan, M.; Kwon, S.-W.; Weon, H.-Y. Isolation, molecular characterization and growth-promoting activities of endophytic sugarcane diazotroph Klebsiella sp. GR9. World J. Microbiol. Biotechnol. 2007, 23, 997–1006. [Google Scholar] [CrossRef]
- Mirza, M.S.; Ahmad, W.; Latif, F.; Haurat, J.; Bally, R.; Normand, P.; Malik, K.A. Isolation, partial characterization, and the effect of plant growth-promoting bacteria (PGPB) on micro-propagated sugarcane in vitro. Plant Soil 2001, 237, 47–54. [Google Scholar] [CrossRef]
- Luo, T.; Ouyang, X.; Yang, L.; Li, Y. Effect of nitrogen-fixing bacteria inoculation on biological nitrogen fixation in sugarcane by 15 N isotope dilution technique. J. Nucl. Agric. Sci. 2010, 24, 1026–1031. [Google Scholar]
- Lin, L.; Li, Z.; Hu, C.; Zhang, X.; Chang, S.; Yang, L.; Li, Y.; An, Q. Plant growth-promoting nitrogen-fixing enterobacteria are in association with sugarcane plants growing in Guangxi, China. Microbes Environ. 2012, 27, 391–398. [Google Scholar] [CrossRef] [Green Version]
- Wei, C.-Y.; Lin, L.; Luo, L.-J.; Xing, Y.-X.; Hu, C.-J.; Yang, L.-T.; Li, Y.-R.; An, Q. Endophytic nitrogen-fixing Klebsiella variicola strain DX120E promotes sugarcane growth. Biol. Fertil. Soils 2014, 50, 657–666. [Google Scholar] [CrossRef]
- Xin, G.; Zhang, G.; Kang, J.W.; Staley, J.T.; Doty, S.L. A diazotrophic, indole-3-acetic acid-producing endophyte from wild cottonwood. Biol. Fertil. Soils 2009, 45, 669–674. [Google Scholar] [CrossRef]
- Khan, Z.; Guelich, G.; Phan, H.; Redman, R.; Doty, S. Bacterial and yeast endophytes from poplar and willow promote growth in crop plants and grasses. ISRN Agron. 2012, 2012, 890280. [Google Scholar] [CrossRef] [Green Version]
- Andrade, L.F. Analysis of the abilities of endophytic bacteria associated with banana tree roots to promote plant growth. J. Microbiol. 2014, 52, 27–34. [Google Scholar] [CrossRef]
- Patel, J.K.; Archana, G. Diverse culturable diazotrophic endophytic bacteria from Poaceae plants show cross-colonization and plant growth promotion in wheat. Plant Soil 2017, 417, 99–116. [Google Scholar] [CrossRef]
- Gupta, G.; Panwar, J.; Jha, P.N. Natural occurrence of Pseudomonas aeruginosa, a dominant cultivable diazotrophic endophytic bacterium colonizing Pennisetum glaucum (L.) R. Br. Appl. Soil Ecol. 2013, 64, 252–261. [Google Scholar] [CrossRef]
- Yan, X.; Wang, Z.; Mei, Y.; Wang, L.; Wang, X.; Xu, Q.; Peng, S.; Zhou, Y.; Wei, C. Isolation, Diversity, and Growth-Promoting Activities of Endophytic Bacteria From Tea Cultivars of Zijuan and Yunkang-10. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Rangjaroen, C.; Rerkasem, B.; Teaumroong, N.; Noisangiam, R.; Lumyong, S. Promoting plant growth in a commercial rice cultivar by endophytic diazotrophic bacteria isolated from rice landraces. Ann. Microbiol. 2015, 65, 253–266. [Google Scholar] [CrossRef]
- Padda, K.P.; Puri, A.; Chanway, C. Endophytic nitrogen fixation—A possible ‘hidden’ source of nitrogen for lodgepole pine trees growing at unreclaimed gravel mining sites. FEMS Microbiol. Ecol. 2019, 95. [Google Scholar] [CrossRef]
- Ji, S.H.; Gururani, M.A.; Chun, S.-C. Isolation and characterization of plant growth promoting endophytic diazotrophic bacteria from Korean rice cultivars. Microbiol. Res. 2014, 169, 83–98. [Google Scholar] [CrossRef] [PubMed]
- Szilagyi-Zecchin, V.J.; Ikeda, A.C.; Hungria, M.; Adamoski, D.; Kava-Cordeiro, V.; Glienke, C.; Galli-Terasawa, L.V. Identification and characterization of endophytic bacteria from corn (Zea mays L.) roots with biotechnological potential in agriculture. AMB Express 2014, 4, 26. [Google Scholar] [CrossRef] [PubMed]
- Anand, R.; Chanway, C.P. nif gene sequence and arrangement in the endophytic diazotroph Paenibacillus polymyxa strain P2b-2R. Biol. Fertil. Soils 2013, 49, 965–970. [Google Scholar] [CrossRef]
- Padda, K.P. Effect of GFP tagging of Paenibacillus polymyxa P2b-2R on its ability to promote growth of canola and tomato seedlings. Biol. Fertil. Soils 2016, 52, 377–387. [Google Scholar] [CrossRef]
- Puri, A.; Padda, K.P.; Chanway, C.P. Evidence of endophytic diazotrophic bacteria in lodgepole pine and hybrid white spruce trees growing in soils with different nutrient statuses in the West Chilcotin region of British Columbia, Canada. For. Ecol. Manag. 2018, 430, 558–565. [Google Scholar] [CrossRef]
- Doty, S.L.; Sher, A.W.; Fleck, N.D.; Khorasani, M.; Bumgarner, R.E.; Khan, Z.; Ko, A.W.K.; Kim, S.-H.; DeLuca, T.H. Variable Nitrogen Fixation in Wild Populus. PLoS ONE 2016, 11, e0155979. [Google Scholar] [CrossRef] [Green Version]
- Araújo, A.E.d.S.; Baldani, V.L.D.; Galisa, P.d.S.; Pereira, J.A.; Baldani, J.I. Response of traditional upland rice varieties to inoculation with selected diazotrophic bacteria isolated from rice cropped at the Northeast region of Brazil. Appl. Soil Ecol. 2013, 64, 49–55. [Google Scholar] [CrossRef] [Green Version]
- Schultz, N.; Silva, J.A.d.; Sousa, J.S.; Monteiro, R.C.; Oliveira, R.P.; Chaves, V.A.; Pereira, W.; Silva, M.F.d.; Baldani, J.I.; Boddey, R.M.; et al. Inoculation of sugarcane with diazotrophic bacteria. Rev. Bras. De Ciência Do Solo 2014, 38, 407–414. [Google Scholar] [CrossRef] [Green Version]
- Fang, K.; Bao, Z.-S.-N.; Chen, L.; Zhou, J.; Yang, Z.-P.; Dong, X.-F.; Zhang, H.-B. Growth-promoting characteristics of potential nitrogen-fixing bacteria in the root of an invasive plant Ageratina adenophora. PeerJ 2019, 7, e7099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gouda, S.; Kerry, R.G.; Das, G.; Paramithiotis, S.; Shin, H.-S.; Patra, J.K. Revitalization of plant growth promoting rhizobacteria for sustainable development in agriculture. Microbiol. Res. 2018, 206, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, H.; Fraga, R. Phosphate solubilizing bacteria and their role in plant growth promotion. Biotechnol. Adv. 1999, 17, 319–339. [Google Scholar] [CrossRef]
- Van Der Heijden, M.G.A.; Bardgett, R.D.; Van Straalen, N.M. The unseen majority: Soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems. Ecol. Lett. 2008, 11, 296–310. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.; Boukhris, I.; Pragya; Kumar, V.; Yadav, A.N.; Farhat-Khemakhem, A.; Kumar, A.; Singh, D.; Blibech, M.; Chouayekh, H.; et al. Contribution of microbial phytases to the improvement of plant growth and nutrition: A review. Pedosphere 2020, 30, 295–313. [Google Scholar] [CrossRef]
- Delvasto, P.; Valverde, A.; Ballester, A.; Muñoz, J.A.; González, F.; Blázquez, M.L.; Igual, J.M.; García-Balboa, C. Diversity and activity of phosphate bioleaching bacteria from a high-phosphorus iron ore. Hydrometallurgy 2008, 92, 124–129. [Google Scholar] [CrossRef]
- Yi, Y.; Huang, W.; Ge, Y. Exopolysaccharide: A novel important factor in the microbial dissolution of tricalcium phosphate. World J. Microbiol. Biotechnol. 2008, 24, 1059–1065. [Google Scholar] [CrossRef]
- Vendan, R.T.; Yu, Y.; Lee, S.; Rhee, Y. Diversity of endophytic bacteria in ginseng and their potential for plant growth promotion. J. Microbiol. 2010, 48, 559–565. [Google Scholar] [CrossRef] [PubMed]
- Palaniappan, P.; Chauhan, P.S.; Saravanan, V.S.; Anandham, R.; Sa, T. Isolation and characterization of plant growth promoting endophytic bacterial isolates from root nodule of Lespedeza sp. Biol. Fertil. Soils 2010, 46, 807–816. [Google Scholar] [CrossRef]
- Dias, A.C.F.; Costa, F.E.C.; Andreote, F.D.; Lacava, P.T.; Teixeira, M.A.; Assumpção, L.C.; Araújo, W.L.; Azevedo, J.L.; Melo, I.S. Isolation of micropropagated strawberry endophytic bacteria and assessment of their potential for plant growth promotion. World J. Microbiol. Biotechnol. 2009, 25, 189–195. [Google Scholar] [CrossRef] [Green Version]
- Puente, M.E.; Li, C.Y.; Bashan, Y. Endophytic bacteria in cacti seeds can improve the development of cactus seedlings. Environ. Exp. Bot. 2009, 66, 402–408. [Google Scholar] [CrossRef]
- Verma, P.; Yadav, A.N.; Khannam, K.S.; Panjiar, N.; Kumar, S.; Saxena, A.K.; Suman, A. Assessment of genetic diversity and plant growth promoting attributes of psychrotolerant bacteria allied with wheat (Triticum aestivum) from the northern hills zone of India. Ann. Microbiol. 2015, 65, 1885–1899. [Google Scholar] [CrossRef]
- Taurian, T.; Anzuay, M.S.; Angelini, J.G.; Tonelli, M.L.; Ludueña, L.; Pena, D.; Ibáñez, F.; Fabra, A. Phosphate-solubilizing peanut associated bacteria: Screening for plant growth-promoting activities. Plant Soil 2010, 329, 421–431. [Google Scholar] [CrossRef]
- Kurek, E.; Ozimek, E.; Sobiczewski, P.; Słomka, A.; Jaroszuk-Ściseł, J. Effect of Pseudomonas luteola on mobilization of phosphorus and growth of young apple trees (Ligol)—Pot experiment. Sci. Hortic. 2013, 164, 270–276. [Google Scholar] [CrossRef]
- Pereira, S.I.A.; Castro, P.M.L. Phosphate-solubilizing rhizobacteria enhance Zea mays growth in agricultural P-deficient soils. Ecol. Eng. 2014, 73, 526–535. [Google Scholar] [CrossRef]
- Otieno, N.; Lally, R.; Kiwanuka, S.; Lloyd, A.; Ryan, D.; Germaine, K.; Dowling, D. Plant growth promotion induced by phosphate solubilizing endophytic Pseudomonas isolates. Front. Microbiol. 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Joe, M.M.; Devaraj, S.; Benson, A.; Sa, T. Isolation of phosphate solubilizing endophytic bacteria from Phyllanthus amarus Schum & Thonn: Evaluation of plant growth promotion and antioxidant activity under salt stress. J. Appl. Res. Med. Aromat. Plants 2016, 3, 71–77. [Google Scholar] [CrossRef]
- Chen, Y.; Fan, J.-B.; Du, L.; Xu, H.; Zhang, Q.-H.; He, Y.-Q. The application of phosphate solubilizing endophyte Pantoea dispersa triggers the microbial community in red acidic soil. Appl. Soil Ecol. 2014, 84, 235–244. [Google Scholar] [CrossRef]
- Castro, R.A.; Dourado, M.N.; Almeida, J.R.d.; Lacava, P.T.; Nave, A.; Melo, I.S.d.; Azevedo, J.L.d.; Quecine, M.C. Mangrove endophyte promotes reforestation tree (Acacia polyphylla) growth. brazilian journal of microbiology 2018, 49, 59–66. [Google Scholar] [CrossRef]
- Ma, Y.; Rajkumar, M.; Zhang, C.; Freitas, H. Beneficial role of bacterial endophytes in heavy metal phytoremediation. J. Environ. Manag. 2016, 174, 14–25. [Google Scholar] [CrossRef] [PubMed]
- Saha, M.; Sarkar, S.; Sarkar, B.; Sharma, B.K.; Bhattacharjee, S.; Tribedi, P. Microbial siderophores and their potential applications: A review. Environ. Sci. Pollut. Res. 2016, 23, 3984–3999. [Google Scholar] [CrossRef] [PubMed]
- Neubauer, U.; Furrer, G.; Kayser, A.; Schulin, R. Siderophores, NTA, and Citrate: Potential Soil Amendments to Enhance Heavy Metal Mobility in Phytoremediation. Int. J. Phytoremediation 2000, 2, 353–368. [Google Scholar] [CrossRef]
- Senthilkumar, M.; Swarnalakshmi, K.; Govindasamy, V.; Lee, Y.K.; Annapurna, K. Biocontrol Potential of Soybean Bacterial Endophytes Against Charcoal Rot Fungus, Rhizoctonia bataticola. Curr. Microbiol. 2008, 58, 288. [Google Scholar] [CrossRef] [PubMed]
- Han, S.; Zhang, S.; Lin, T.; Gong, M. Screening for siderophore-producing endophytic bacteria against Fusarium oxysporum. Agric. Sci. Technol. Hunan 2011, 12, 994–996. [Google Scholar]
- Lacava, P.T.; Silva-Stenico, M.E.; Araújo, W.L.; Simionato, A.V.C.; Carrilho, E.; Tsai, S.M.; Azevedo, J.L. Detection of siderophores in endophytic bacteria Methylobacterium spp. associated with Xylella fastidiosa subsp. pauca. Pesqui. Agropecuária Bras. 2008, 43, 521–528. [Google Scholar] [CrossRef] [Green Version]
- Pandey, P.; Irulappan, V.; Bagavathiannan, M.V.; Senthil-Kumar, M. Impact of Combined Abiotic and Biotic Stresses on Plant Growth and Avenues for Crop Improvement by Exploiting Physio-morphological Traits. Front. Plant Sci. 2017, 8, 537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, N.; Rivero, R.M.; Shulaev, V.; Blumwald, E.; Mittler, R. Abiotic and biotic stress combinations. New Phytol. 2014, 203, 32–43. [Google Scholar] [CrossRef]
- Kumar, A.; Verma, J.P. Does plant—Microbe interaction confer stress tolerance in plants: A review? Microbiol. Res. 2018, 207, 41–52. [Google Scholar] [CrossRef]
- Meena, K.K.; Sorty, A.M.; Bitla, U.M.; Choudhary, K.; Gupta, P.; Pareek, A.; Singh, D.P.; Prabha, R.; Sahu, P.K.; Gupta, V.K.; et al. Abiotic Stress Responses and Microbe-Mediated Mitigation in Plants: The Omics Strategies. Front. Plant Sci. 2017, 8, 172. [Google Scholar] [CrossRef] [PubMed]
- Lata, R.; Chowdhury, S.; Gond, S.K.; White Jr, J.F. Induction of abiotic stress tolerance in plants by endophytic microbes. Lett. Appl. Microbiol. 2018, 66, 268–276. [Google Scholar] [CrossRef] [Green Version]
- Ullah, A.; Nisar, M.; Ali, H.; Hazrat, A.; Hayat, K.; Keerio, A.A.; Ihsan, M.; Laiq, M.; Ullah, S.; Fahad, S.; et al. Drought tolerance improvement in plants: An endophytic bacterial approach. Appl. Microbiol. Biotechnol. 2019, 103, 7385–7397. [Google Scholar] [CrossRef] [PubMed]
- Razzaghi Komaresofla, B.; Alikhani, H.A.; Etesami, H.; Khoshkholgh-Sima, N.A. Improved growth and salinity tolerance of the halophyte Salicornia sp. by co–inoculation with endophytic and rhizosphere bacteria. Appl. Soil Ecol. 2019, 138, 160–170. [Google Scholar] [CrossRef]
- Pandey, P.K.; Samanta, R.; Yadav, R.N.S. Inside the plant: Addressing bacterial endophytes in biotic stress alleviation. Arch. Microbiol. 2019, 201, 415–429. [Google Scholar] [CrossRef]
- Khan, N.A.; Khan, M.I.R. The ethylene: From senescence hormone to key player in plant metabolism. J. Plant Biochem. Physiol. 2014. [Google Scholar] [CrossRef]
- Tiwari, G.; Duraivadivel, P.; Sharma, S.; Hariprasad, P. 1-Aminocyclopropane-1-carboxylic acid deaminase producing beneficial rhizobacteria ameliorate the biomass characters of Panicum maximum Jacq. by mitigating drought and salt stress. Sci. Rep. 2018, 8, 17513. [Google Scholar] [CrossRef]
- Grobelak, A.; Kokot, P.; Świątek, J.; Jaskulak, M.; Rorat, A. Bacterial ACC Deaminase Activity in Promoting Plant Growth on Areas Contaminated with Heavy Metals. J. Ecol. Eng. 2018, 19, 150–157. [Google Scholar] [CrossRef]
- Afridi, M.S.; Mahmood, T.; Salam, A.; Mukhtar, T.; Mehmood, S.; Ali, J.; Khatoon, Z.; Bibi, M.; Javed, M.T.; Sultan, T. Induction of tolerance to salinity in wheat genotypes by plant growth promoting endophytes: Involvement of ACC deaminase and antioxidant enzymes. Plant Physiol. Biochem. 2019, 139, 569–577. [Google Scholar] [CrossRef]
- Glick, B.R. Bacteria with ACC deaminase can promote plant growth and help to feed the world. Microbiol. Res. 2014, 169, 30–39. [Google Scholar] [CrossRef]
- Ali, S.; Charles, T.C.; Glick, B.R. Amelioration of high salinity stress damage by plant growth-promoting bacterial endophytes that contain ACC deaminase. Plant Physiol. Biochem. 2014, 80, 160–167. [Google Scholar] [CrossRef] [PubMed]
- Złoch, M.; Thiem, D.; Gadzała-Kopciuch, R.; Hrynkiewicz, K. Synthesis of siderophores by plant-associated metallotolerant bacteria under exposure to Cd2+. Chemosphere 2016, 156, 312–325. [Google Scholar] [CrossRef] [PubMed]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant Drought Stress: Effects, Mechanisms and Management. In Sustainable Agriculture; Lichtfouse, E., Navarrete, M., Debaeke, P., Véronique, S., Alberola, C., Eds.; Springer: Dordrecht, The Netherlands, 2009; pp. 153–188. [Google Scholar]
- Barka, E.A.; Nowak, J.; Clément, C. Enhancement of chilling resistance of inoculated grapevine plantlets with a plant growth-promoting rhizobacterium, Burkholderia phytofirmans strain PsJN. Appl. Environ. Microbiol. 2006, 72, 7246–7252. [Google Scholar] [CrossRef] [Green Version]
- Fernandez, O.; Theocharis, A.; Bordiec, S.; Feil, R.; Jacquens, L.; Clément, C.; Fontaine, F.; Barka, E.A. Burkholderia phytofirmans PsJN Acclimates Grapevine to Cold by Modulating Carbohydrate Metabolism. Mol. Plant-Microbe Interact. 2012, 25, 496–504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naveed, M.; Mitter, B.; Reichenauer, T.G.; Wieczorek, K.; Sessitsch, A. Increased drought stress resilience of maize through endophytic colonization by Burkholderia phytofirmans PsJN and Enterobacter sp. FD17. Environ. Exp. Bot. 2014, 97, 30–39. [Google Scholar] [CrossRef]
- Vargas, L.; Santa Brígida, A.B.; Mota Filho, J.P.; de Carvalho, T.G.; Rojas, C.A.; Vaneechoutte, D.; Van Bel, M.; Farrinelli, L.; Ferreira, P.C.G.; Vandepoele, K.; et al. Drought tolerance conferred to sugarcane by association with Gluconacetobacter diazotrophicus: A transcriptomic view of hormone pathways. PLoS ONE 2014, 9, e114744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Timmusk, S.; El-Daim, I.A.A.; Copolovici, L.; Tanilas, T.; Kännaste, A.; Behers, L.; Nevo, E.; Seisenbaeva, G.; Stenström, E.; Niinemets, Ü. Drought-tolerance of wheat improved by rhizosphere bacteria from harsh environments: Enhanced biomass production and reduced emissions of stress volatiles. PLoS ONE 2014, 9, e96086. [Google Scholar] [CrossRef] [Green Version]
- Jiao, J.; Ma, Y.; Chen, S.; Liu, C.; Song, Y.; Qin, Y.; Yuan, C.; Liu, Y. Melatonin-Producing Endophytic Bacteria from Grapevine Roots Promote the Abiotic Stress-Induced Production of Endogenous Melatonin in Their Hosts. Front. Plant Sci. 2016, 7, 1387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Xin, K.; Liu, H.; Cheng, J.; Shen, X.; Wang, Y.; Zhang, L. Pantoea alhagi, a novel endophytic bacterium with ability to improve growth and drought tolerance in wheat. Sci. Rep. 2017, 7, 41564. [Google Scholar] [CrossRef] [Green Version]
- Vigani, G.; Rolli, E.; Marasco, R.; Dell’Orto, M.; Michoud, G.; Soussi, A.; Raddadi, N.; Borin, S.; Sorlini, C.; Zocchi, G.; et al. Root bacterial endophytes confer drought resistance and enhance expression and activity of a vacuolar H+-pumping pyrophosphatase in pepper plants. Environ. Microbiol. 2019, 21, 3212–3228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Z.; Chu, Y.; Zhang, W.; Lang, D.; Zhang, X. Bacillus pumilus alleviates drought stress and increases metabolite accumulation in Glycyrrhiza uralensis Fisch. Environ. Exp. Bot. 2019, 158, 99–106. [Google Scholar] [CrossRef]
- Jayakumar, A.; Padmakumar, P.; Nair, I.C.; Radhakrishnan, E.K. Drought tolerant bacterial endophytes with potential plant probiotic effects from Ananas comosus. Biologia 2020, 75, 1769–1778. [Google Scholar] [CrossRef]
- Naya, L.; Ladrera, R.; Ramos, J.; González, E.M.; Arrese-Igor, C.; Minchin, F.R.; Becana, M. The Response of Carbon Metabolism and Antioxidant Defenses of Alfalfa Nodules to Drought Stress and to the Subsequent Recovery of Plants. Plant Physiol. 2007, 144, 1104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yachana, J.; Subramanian, R.B.; Suchita, P. Combination of endophytic and rhizospheric plant growth promoting rhizobacteria in Oryza sativa shows higher accumulation of osmoprotectant against saline stress. Acta Physiol. Plant. 2011, 33, 797–802. [Google Scholar] [CrossRef]
- Han, Y.; Wang, R.; Yang, Z.; Zhan, Y.; Ma, Y.; Ping, S.; Zhang, L.; Lin, M.; Yan, Y. 1-Aminocyclopropane-1-Carboxylate Deaminase from Pseudomonas stutzeri A1501 Facilitates the Growth of Rice in the Presence of Salt or Heavy Metals. J. Microbiol. Biotechnol. 2015, 25, 1119–1128. [Google Scholar] [CrossRef] [PubMed]
- Sorty, A.M.; Meena, K.K.; Choudhary, K.; Bitla, U.M.; Minhas, P.S.; Krishnani, K.K. Effect of Plant Growth Promoting Bacteria Associated with Halophytic Weed (Psoralea corylifolia L) on Germination and Seedling Growth of Wheat Under Saline Conditions. Appl. Biochem. Biotechnol. 2016, 180, 872–882. [Google Scholar] [CrossRef]
- Ma, Y.; Rajkumar, M.; Freitas, H. Inoculation of plant growth promoting bacterium Achromobacter xylosoxidans strain Ax10 for the improvement of copper phytoextraction by Brassica juncea. J. Environ. Manag. 2009, 90, 831–837. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.-f.; He, L.-y.; Chen, Z.-j.; Wang, Q.-y.; Qian, M.; Sheng, X.-f. Characterization of ACC deaminase-producing endophytic bacteria isolated from copper-tolerant plants and their potential in promoting the growth and copper accumulation of Brassica napus. Chemosphere 2011, 83, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Abd_Allah, E.F.; Alqarawi, A.A.; Hashem, A.; Radhakrishnan, R.; Al-Huqail, A.A.; Al-Otibi, F.O.N.; Malik, J.A.; Alharbi, R.I.; Egamberdieva, D. Endophytic bacterium Bacillus subtilis (BERA 71) improves salt tolerance in chickpea plants by regulating the plant defense mechanisms. J. Plant Interact. 2018, 13, 37–44. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.A.; Asaf, S.; Khan, A.L.; Adhikari, A.; Jan, R.; Ali, S.; Imran, M.; Kim, K.M.; Lee, I.J. Plant growth-promoting endophytic bacteria augment growth and salinity tolerance in rice plants. Plant Biol. 2020, 22, 850–862. [Google Scholar] [CrossRef] [PubMed]
- Abdelshafy Mohamad, O.A.; Ma, J.-B.; Liu, Y.-H.; Zhang, D.; Hua, S.; Bhute, S.; Hedlund, B.P.; Li, W.-J.; Li, L. Beneficial Endophytic Bacterial Populations Associated With Medicinal Plant Thymus vulgaris Alleviate Salt Stress and Confer Resistance to Fusarium oxysporum. Front. Plant Sci. 2020, 11, 47. [Google Scholar] [CrossRef]
- Fan, M.; Liu, Z.; Nan, L.; Wang, E.; Chen, W.; Lin, Y.; Wei, G. Isolation, characterization, and selection of heavy metal-resistant and plant growth-promoting endophytic bacteria from root nodules of Robinia pseudoacacia in a Pb/Zn mining area. Microbiol. Res. 2018, 217, 51–59. [Google Scholar] [CrossRef]
- Jan, R.; Khan, M.A.; Asaf, S.; Lubna; Lee, I.-J.; Kim, K.M. Metal Resistant Endophytic Bacteria Reduces Cadmium, Nickel Toxicity, and Enhances Expression of Metal Stress Related Genes with Improved Growth of Oryza Sativa, via Regulating Its Antioxidant Machinery and Endogenous Hormones. Plants 2019, 8, 363. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Li, N.; Han, S.; Sun, Y.; Wang, L.; Qu, Z.; Dai, M.; Zhao, G. Characterization and bioremediation potential of nickel-resistant endophytic bacteria isolated from the wetland plant Tamarix chinensis. FEMS Microbiol. Lett. 2020, 367. [Google Scholar] [CrossRef]
- Feng, N.-X.; Yu, J.; Zhao, H.-M.; Cheng, Y.-T.; Mo, C.-H.; Cai, Q.-Y.; Li, Y.-W.; Li, H.; Wong, M.-H. Efficient phytoremediation of organic contaminants in soils using plant–endophyte partnerships. Sci. Total Environ. 2017, 583, 352–368. [Google Scholar] [CrossRef] [PubMed]
- Phillips, L.A.; Germida, J.J.; Farrell, R.E.; Greer, C.W. Hydrocarbon degradation potential and activity of endophytic bacteria associated with prairie plants. Soil Biol. Biochem. 2008, 40, 3054–3064. [Google Scholar] [CrossRef]
- Van Aken, B.; Yoon, J.M.; Schnoor, J.L. Biodegradation of nitro-substituted explosives 2,4,6-trinitrotoluene, hexahydro-1,3,5-trinitro-1,3,5-triazine, and octahydro-1,3,5,7-tetranitro-1,3,5-tetrazocine by a phytosymbiotic Methylobacterium sp. associated with poplar tissues (Populus deltoides x nigra DN34). Appl. Environ. Microbiol. 2004, 70, 508–517. [Google Scholar] [CrossRef] [Green Version]
- Germaine, K.J.; Liu, X.; Cabellos, G.G.; Hogan, J.P.; Ryan, D.; Dowling, D.N. Bacterial endophyte-enhanced phytoremediation of the organochlorine herbicide 2,4-dichlorophenoxyacetic acid. FEMS Microbiol. Ecol. 2006, 57, 302–310. [Google Scholar] [CrossRef]
- Ashraf, S.; Afzal, M.; Naveed, M.; Shahid, M.; Ahmad Zahir, Z. Endophytic bacteria enhance remediation of tannery effluent in constructed wetlands vegetated with Leptochloa fusca. Int. J. Phytoremediation 2018, 20, 121–128. [Google Scholar] [CrossRef]
- Ahsan, M.T.; Najam-ul-haq, M.; Idrees, M.; Ullah, I.; Afzal, M. Bacterial endophytes enhance phytostabilization in soils contaminated with uranium and lead. Int. J. Phytoremediation 2017, 19, 937–946. [Google Scholar] [CrossRef]
- Eevers, N.; Hawthorne, J.R.; White, J.C.; Vangronsveld, J.; Weyens, N. Endophyte-enhanced phytoremediation of DDE-contaminated using Cucurbita pepo: A field trial. Int. J. Phytoremediation 2018, 20, 301–310. [Google Scholar] [CrossRef]
- Mitter, E.K.; Kataoka, R.; de Freitas, J.R.; Germida, J.J. Potential use of endophytic root bacteria and host plants to degrade hydrocarbons. Int. J. Phytoremediation 2019, 21, 928–938. [Google Scholar] [CrossRef]
- Wu, T.; Xu, J.; Liu, J.; Guo, W.-H.; Li, X.-B.; Xia, J.-B.; Xie, W.-J.; Yao, Z.-G.; Zhang, Y.-M.; Wang, R.-Q. Characterization and Initial Application of Endophytic Bacillus safensis Strain ZY16 for Improving Phytoremediation of Oil-Contaminated Saline Soils. Front. Microbiol. 2019, 10, 991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Latha, P.; Karthikeyan, M.; Rajeswari, E. Endophytic Bacteria: Prospects and Applications for the Plant Disease Management. In Plant Health Under Biotic Stress: Volume 2: Microbial Interactions; Ansari, R.A., Mahmood, I., Eds.; Springer: Singapore, 2019; pp. 1–50. [Google Scholar]
- Lashin, I.; Fouda, A.; Gobouri, A.A.; Azab, E.; Mohammedsaleh, Z.M.; Makharita, R.R. Antimicrobial and In Vitro Cytotoxic Efficacy of Biogenic Silver Nanoparticles (Ag-NPs) Fabricated by Callus Extract of Solanum incanum L. Biomolecules 2021, 11, 341. [Google Scholar] [CrossRef]
- Ongena, M.; Jacques, P. Bacillus lipopeptides: Versatile weapons for plant disease biocontrol. Trends Microbiol. 2008, 16, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Fouda, A.; Hassan, S.E.D.; Eid, A.M.; El-Din Ewais, E. The Interaction Between Plants and Bacterial Endophytes Under Salinity Stress. In Endophytes and Secondary Metabolites; Jha, S., Ed.; Springer International Publishing: Cham, Switzerland, 2018; pp. 1–17. [Google Scholar]
- Blumenstein, K.; Albrectsen, B.R.; Martín, J.A.; Hultberg, M.; Sieber, T.N.; Helander, M.; Witzell, J. Nutritional niche overlap potentiates the use of endophytes in biocontrol of a tree disease. BioControl 2015, 60, 655–667. [Google Scholar] [CrossRef]
- Höfte, M. Classes of Microbial Siderophores; Academic Press: San Diego, CA, USA, 1993. [Google Scholar]
- Cordova-Albores, L.C.; Zapotitla, E.S.; Ríos, M.Y.; Barrera-Necha, L.L.; Hernández-López, M.; Bautista-Baños, S. Microscopic study of the morphology and metabolic activity of Fusarium oxysporum f. sp. gladioli treated with Jatropha curcas oil and derivatives. J. Microsc. Ultrastruct. 2016, 4, 28–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres, R.; Solsona, C.; Viñas, I.; Usall, J.; Plaza, P.; Teixidó, N. Optimization of packaging and storage conditions of a freeze-dried Pantoea agglomerans formulation for controlling postharvest diseases in fruit. J. Appl. Microbiol. 2014, 117, 173–184. [Google Scholar] [CrossRef]
- Fialho, M.B.; Moraes, M.H.D.d.; Tremocoldi, A.R.; Pascholati, S.F. Potential of antimicrobial volatile organic compounds to control Sclerotinia sclerotiorum in bean seeds. Pesqui. Agropecuária Bras. 2011, 46, 137–142. [Google Scholar] [CrossRef] [Green Version]
- Jasim, B.; Sreelakshmi, K.S.; Mathew, J.; Radhakrishnan, E.K. Surfactin, Iturin, and Fengycin Biosynthesis by Endophytic Bacillus sp. from Bacopa monnieri. Microb. Ecol. 2016, 72, 106–119. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, F.T.; Islam, M.R.; Islam, M.R.; Khan, H. Diversity of Plant Endophytic Volatile Organic Compound (VOC) and Their Potential Applications. In Endophytes and Secondary Metabolites; Jha, S., Ed.; Springer International Publishing: Cham, Switzerland, 2018; pp. 1–27. [Google Scholar]
- Spadaro, D.; Droby, S. Development of biocontrol products for postharvest diseases of fruit: The importance of elucidating the mechanisms of action of yeast antagonists. Trends Food Sci. Technol. 2016, 47, 39–49. [Google Scholar] [CrossRef]
- Piromyou, P.; Songwattana, P.; Greetatorn, T.; Okubo, T.; Chiba Kakizaki, K.; Prakamhang, J.; Tittabutr, P.; Boonkerd, N.; Teaumroong, N.; Minamisawa, K. The Type III Secretion System (T3SS) is a Determinant for Rice-Endophyte Colonization by Non-Photosynthetic Bradyrhizobium. Microbes Environ. 2015, advpub. [Google Scholar] [CrossRef] [Green Version]
- Backman, P.A.; Sikora, R.A. Endophytes: An emerging tool for biological control. Biol. Control 2008, 46, 1–3. [Google Scholar] [CrossRef]
- Miliute, I.; Buzaite, O.; Baniulis, D.; Stanys, V. Bacterial endophytes in agricultural crops and their role in stress tolerance: A review. Žemdirbystė (Agric.) 2015, 102, 465–478. [Google Scholar] [CrossRef] [Green Version]
- Rashid, M.H.-O.; Khan, A.; Hossain, M.T.; Chung, Y.R. Induction of Systemic Resistance against Aphids by Endophytic Bacillus velezensis YC7010 via Expressing PHYTOALEXIN DEFICIENT4 in Arabidopsis. Front. Plant Sci. 2017, 8, 211. [Google Scholar] [CrossRef] [Green Version]
- Lugtenberg, B.; Kamilova, F. Plant-Growth-Promoting Rhizobacteria. Annu. Rev. Microbiol. 2009, 63, 541–556. [Google Scholar] [CrossRef] [Green Version]
- Maloy, O.C.; Murray, T.D. Encyclopedia of Plant Pathology; Wiley: New York, NY, USA, 2001. [Google Scholar]
- Uroz, S.; D’Angelo-Picard, C.; Carlier, A.; Elasri, M.; Sicot, C.; Petit, A.; Oger, P.; Faure, D.; Dessaux, Y. Novel bacteria degrading N-acylhomoserine lactones and their use as quenchers of quorum-sensing-regulated functions of plant-pathogenic bacteria. Microbiol. (Read. Engl.) 2003, 149, 1981–1989. [Google Scholar] [CrossRef] [Green Version]
- Gao, Z.; Zhang, B.; Liu, H.; Han, J.; Zhang, Y. Identification of endophytic Bacillus velezensis ZSY-1 strain and antifungal activity of its volatile compounds against Alternaria solani and Botrytis cinerea. Biol. Control 2017, 105, 27–39. [Google Scholar] [CrossRef]
- Perneel, M.; D’Hondt, L.; De Maeyer, K.; Adiobo, A.; Rabaey, K.; Höfte, M. Phenazines and biosurfactants interact in the biological control of soil-borne diseases caused by Pythium spp. Environ. Microbiol. 2008, 10, 778–788. [Google Scholar] [CrossRef]
- Zeriouh, H.; Romero, D.; García-Gutiérrez, L.; Cazorla, F.M.; de Vicente, A.; Pérez-García, A. The Iturin-like Lipopeptides Are Essential Components in the Biological Control Arsenal of Bacillus subtilis Against Bacterial Diseases of Cucurbits. Mol. Plant-Microbe Interact. 2011, 24, 1540–1552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tabbene, O.; Ben Slimene, I.; Bouabdallah, F.; Mangoni, M.-L.; Urdaci, M.-C.; Limam, F. Production of Anti-Methicillin-Resistant Staphylococcus Activity from Bacillus subtilis sp. Strain B38 Newly Isolated from Soil. Appl. Biochem. Biotechnol. 2009, 157, 407–419. [Google Scholar] [CrossRef] [PubMed]
- Pliego, C.; Kamilova, F.; Lugtenberg, B. Plant Growth-Promoting Bacteria: Fundamentals and Exploitation. In Bacteria in Agrobiology: Crop Ecosystems; Maheshwari, D.K., Ed.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 295–343. [Google Scholar]
- Silva, H.S.A.; Romeiro, R.d.S.; Macagnan, D.; Halfeld-Vieira, B.d.A.; Pereira, M.C.B.; Mounteer, A. Rhizobacterial induction of systemic resistance in tomato plants: Non-specific protection and increase in enzyme activities. Biol. Control 2004, 29, 288–295. [Google Scholar] [CrossRef]
- Schuhegger, R.; Ihring, A.; Gantner, S.; Bahnweg, G.; Knappe, C.; Vogg, G.; Hutzler, P.; Schmid, M.; Van Breusegem, F.; Eberl, L.E.O.; et al. Induction of systemic resistance in tomato by N-acyl-L-homoserine lactone-producing rhizosphere bacteria. Plant Cell Environ. 2006, 29, 909–918. [Google Scholar] [CrossRef] [PubMed]
- Audenaert, K.; Pattery, T.; Cornelis, P.; Höfte, M. Induction of Systemic Resistance to Botrytis cinerea in Tomato by Pseudomonas aeruginosa 7NSK2: Role of Salicylic Acid, Pyochelin, and Pyocyanin. Mol. Plant-Microbe Interact. 2002, 15, 1147–1156. [Google Scholar] [CrossRef] [Green Version]
- Newman, D.J.; Cragg, G.M. Chapter 1 Natural Products as Drugs and Leads to Drugs: The Historical Perspective. In Natural Product Chemistry for Drug Discovery; The Royal Society of Chemistry: London, UK, 2009; pp. 3–27. [Google Scholar]
- Kapoor, N.; Jamwal, V.L.; Gandhi, S.G. Endophytes as a Source of High-Value, Bioactive Metabolites; Springer: Berlin, Germany, 2018. [Google Scholar] [CrossRef]
- Ek-Ramos, M.J.; Gomez-Flores, R.; Orozco-Flores, A.A.; Rodríguez-Padilla, C.; González-Ochoa, G.; Tamez-Guerra, P. Bioactive Products From Plant-Endophytic Gram-Positive Bacteria. Front. Microbiol. 2019, 10, 463. [Google Scholar] [CrossRef] [PubMed]
- Alvin, A.; Miller, K.I.; Neilan, B.A. Exploring the potential of endophytes from medicinal plants as sources of antimycobacterial compounds. Microbiol. Res. 2014, 169, 483–495. [Google Scholar] [CrossRef]
- Monowar, T.; Rahman, M.S.; Bhore, S.J.; Raju, G.; Sathasivam, K.V. Secondary Metabolites Profiling of Acinetobacter baumannii Associated with Chili (Capsicum annuum L.) Leaves and Concentration Dependent Antioxidant and Prooxidant Properties. Biomed Res. Int. 2019, 2019, 6951927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdalla, M.A.; Matasyoh, J.C. Endophytes as producers of peptides: An overview about the recently discovered peptides from endophytic microbes. Nat. Prod. Bioprospecting 2014, 4, 257–270. [Google Scholar] [CrossRef] [Green Version]
- Singh, M.; Kumar, A.; Singh, R.; Pandey, K.D. Endophytic bacteria: A new source of bioactive compounds. 3 Biotech 2017, 7, 315. [Google Scholar] [CrossRef] [PubMed]
- Hashem, A.; Abd_Allah, E.F.; Alqarawi, A.A.; Al-Huqail, A.A.; Shah, M.A. Induction of Osmoregulation and Modulation of Salt Stress in Acacia gerrardii Benth. by Arbuscular Mycorrhizal Fungi and Bacillus subtilis (BERA 71). Biomed Res. Int. 2016, 2016, 6294098. [Google Scholar] [CrossRef] [Green Version]
- Mitsuhashi, S. Current topics in the biotechnological production of essential amino acids, functional amino acids, and dipeptides. Curr. Opin. Biotechnol. 2014, 26, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Geddes, C.C.; Nieves, I.U.; Ingram, L.O. Advances in ethanol production. Curr. Opin. Biotechnol. 2011, 22, 312–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soliman, A.M.; Abdel-Latif, W.; Shehata, I.H.; Fouda, A.; Abdo, A.M.; Ahmed, Y.M. Green Approach to Overcome the Resistance Pattern of Candida spp. Using Biosynthesized Silver Nanoparticles Fabricated by Penicillium chrysogenum F9. Biological Trace Element Research 2021, 199, 800–811. [Google Scholar] [CrossRef] [PubMed]
- Shaligram, N.S.; Singhal, R.S. Surfactin—A review on biosynthesis, fermentation, purification and applications. Food Technol. Biotechnol. 2010, 48, 119–134. [Google Scholar]
- Yoon, Y.J.; Kim, E.S.; Hwang, Y.S.; Choi, C.Y. Avermectin: Biochemical and molecular basis of its biosynthesis and regulation. Appl. Microbiol. Biotechnol. 2004, 63, 626–634. [Google Scholar] [CrossRef]
- Ntabo, R.M.; Nyamache, A.K.; Lwande, W.; Kabii, J.; Nonoh, J. Enzymatic Activity of Endophytic Bacterial Isolates from Selected Mangrove Plants in Kenya. Open Microbiol. J. 2018, 12, 354–363. [Google Scholar] [CrossRef] [Green Version]
- Wu, T.; Xu, J.; Xie, W.; Yao, Z.; Yang, H.; Sun, C.; Li, X. Pseudomonas aeruginosa L10: A Hydrocarbon-Degrading, Biosurfactant-Producing, and Plant-Growth-Promoting Endophytic Bacterium Isolated From a Reed (Phragmites australis). Front. Microbiol. 2018, 9, 1087. [Google Scholar] [CrossRef]
- Baker, S.; Sahana, S.; Rakshith, D.; Kavitha, H.; Kavitha, K.; Satish, S. Biodecaffeination by endophytic Pseudomonas sp. isolated from Coffee arabica L. J. Pharm. Res. 2012, 5, 3654–3657. [Google Scholar]
- Mohamed, A.A.; Fouda, A.; Abdel-Rahman, M.A.; Hassan, S.E.D.; El-Gamal, M.S.; Salem, S.S.; Shaheen, T.I. Fungal strain impacts the shape, bioactivity and multifunctional properties of green synthesized zinc oxide nanoparticles. Biocatal. Agric. Biotechnol. 2019, 19. [Google Scholar] [CrossRef]
- Aref, M.S.; Salem, S.S. Bio-callus synthesis of silver nanoparticles, characterization, and antibacterial activities via Cinnamomum camphora callus culture. Biocatal. Agric. Biotechnol. 2020, 27. [Google Scholar] [CrossRef]
- Shaheen, T.I.; Fouda, A.; Salem, S.S. Integration of Cotton Fabrics with Biosynthesized CuO Nanoparticles for Bactericidal Activity in the Terms of Their Cytotoxicity Assessment. Ind. Eng. Chem. Res. 2021, 60, 1553–1563. [Google Scholar] [CrossRef]
- Fouda, A.; Abdel-Maksoud, G.; Abdel-Rahman, M.A.; Eid, A.M.; Barghoth, M.G.; El-Sadany, M.A.H. Monitoring the effect of biosynthesized nanoparticles against biodeterioration of cellulose-based materials by Aspergillus niger. Cellulose 2019, 26, 6583–6597. [Google Scholar] [CrossRef]
- Fouda, A.; Abdel-Maksoud, G.; Abdel-Rahman, M.A.; Salem, S.S.; Hassan, S.E.-D.; El-Sadany, M.A.-H. Eco-friendly approach utilizing green synthesized nanoparticles for paper conservation against microbes involved in biodeterioration of archaeological manuscript. Int. Biodeterior. Biodegrad. 2019, 142, 160–169. [Google Scholar] [CrossRef]
- Fouda, A.; Hassan, S.E.-D.; Saied, E.; Azab, M.S. An eco-friendly approach to textile and tannery wastewater treatment using maghemite nanoparticles (γ-Fe2O3-NPs) fabricated by Penicillium expansum strain (K-w). J. Environ. Chem. Eng. 2021, 9, 104693. [Google Scholar] [CrossRef]
- Fouda, A.; Hassan, S.E.-D.; Saied, E.; Hamza, M.F. Photocatalytic degradation of real textile and tannery effluent using biosynthesized magnesium oxide nanoparticles (MgO-NPs), heavy metal adsorption, phytotoxicity, and antimicrobial activity. J. Environ. Chem. Eng. 2021, 9, 105346. [Google Scholar] [CrossRef]
- Salem, S.S.; Fouda, A. Green Synthesis of Metallic Nanoparticles and Their Prospective Biotechnological Applications: An Overview. Biol. Trace Elem. Res. 2021, 199, 344–370. [Google Scholar] [CrossRef]
- El-Belely, E.F.; Farag, M.M.S.; Said, H.A.; Amin, A.S.; Azab, E.; Gobouri, A.A.; Fouda, A. Green Synthesis of Zinc Oxide Nanoparticles (ZnO-NPs) Using Arthrospira platensis (Class: Cyanophyceae) and Evaluation of their Biomedical Activities. Nanomaterials 2021, 11, 95. [Google Scholar] [CrossRef]
- Rahman, S.; Rahman, L.; Khalil, A.T.; Ali, N.; Zia, D.; Ali, M.; Shinwari, Z.K. Endophyte-mediated synthesis of silver nanoparticles and their biological applications. Appl. Microbiol. Biotechnol. 2019, 103, 2551–2569. [Google Scholar] [CrossRef]
- Monowar, T.; Rahman, M.S.; Bhore, S.J.; Raju, G.; Sathasivam, K.V. Silver Nanoparticles Synthesized by Using the Endophytic Bacterium Pantoea ananatis are Promising Antimicrobial Agents against Multidrug Resistant Bacteria. Molecules 2018, 23, 3220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fouda, A.; Hassan, S.E.D.; Abdo, A.M.; El-Gamal, M.S. Antimicrobial, Antioxidant and Larvicidal Activities of Spherical Silver Nanoparticles Synthesized by Endophytic Streptomyces spp. Biol. Trace Elem. Res. 2020, 195, 707–724. [Google Scholar] [CrossRef]
- Hassan, S.E.L.D.; Salem, S.S.; Fouda, A.; Awad, M.A.; El-Gamal, M.S.; Abdo, A.M. New approach for antimicrobial activity and bio-control of various pathogens by biosynthesized copper nanoparticles using endophytic actinomycetes. J. Radiat. Res. Appl. Sci. 2018, 11, 262–270. [Google Scholar] [CrossRef] [Green Version]
- Hassan, S.E.-D.; Fouda, A.; Radwan, A.A.; Salem, S.S.; Barghoth, M.G.; Awad, M.A.; Abdo, A.M.; El-Gamal, M.S. Endophytic actinomycetes Streptomyces spp mediated biosynthesis of copper oxide nanoparticles as a promising tool for biotechnological applications. JBIC J. Biol. Inorg. Chem. 2019, 24, 377–393. [Google Scholar] [CrossRef]
- Baker, S.; Satish, S. Biosynthesis of gold nanoparticles by Pseudomonas veronii AS41G inhabiting Annona squamosa L. Spectrochim. Acta Part A: Mol. Biomol. Spectrosc. 2015, 150, 691–695. [Google Scholar] [CrossRef]
- El-Moslamy, S.H. Bioprocessing strategies for cost-effective large-scale biogenic synthesis of nano-MgO from endophytic Streptomyces coelicolor strain E72 as an anti-multidrug-resistant pathogens agent. Sci. Rep. 2018, 8, 3820. [Google Scholar] [CrossRef] [PubMed]
- Yousef, N.; Mawad, A.; Abeed, A. Enhancement the Cellulase Activity Induced by Endophytic Bacteria Using Calcium Nanoparticles. Curr. Microbiol. 2019, 76, 346–354. [Google Scholar] [CrossRef]
- Rajabairavi, N.; Raju, C.S.; Karthikeyan, C.; Varutharaju, K.; Nethaji, S.; Hameed, A.S.H.; Shajahan, A. Biosynthesis of Novel Zinc Oxide Nanoparticles (ZnO NPs) Using Endophytic Bacteria Sphingobacterium thalpophilum. In Proceedings of the Recent Trends in Materials Science and Applications; Springer: Cham, Switzerland, 2017; pp. 245–254. [Google Scholar]
- Ibrahim, E.; Fouad, H.; Zhang, M.; Zhang, Y.; Qiu, W.; Yan, C.; Li, B.; Mo, J.; Chen, J. Biosynthesis of silver nanoparticles using endophytic bacteria and their role in inhibition of rice pathogenic bacteria and plant growth promotion. RSC Adv. 2019, 9, 29293–29299. [Google Scholar] [CrossRef] [Green Version]
- Eid, A.M.; Fouda, A.; Niedbała, G.; Hassan, S.E.D.; Salem, S.S.; Abdo, A.M.; Hetta, H.F.; Shaheen, T.I. Endophytic Streptomyces laurentii mediated green synthesis of Ag-NPs with antibacterial and anticancer properties for developing functional textile fabric properties. Antibiotics 2020, 9, 641. [Google Scholar] [CrossRef]
- Salem, S.S.; El-Belely, E.F.; Niedbała, G.; Alnoman, M.M.; Hassan, S.E.D.; Eid, A.M.; Shaheen, T.I.; Elkelish, A.; Fouda, A. Bactericidal and in-vitro cytotoxic efficacy of silver nanoparticles (Ag-NPs) fabricated by endophytic actinomycetes and their use as coating for the textile fabrics. Nanomaterials 2020, 10, 82. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, E.; Zhang, M.; Zhang, Y.; Hossain, A.; Qiu, W.; Chen, Y.; Wang, Y.; Wu, W.; Sun, G.; Li, B. Green-Synthesization of Silver Nanoparticles Using Endophytic Bacteria Isolated from Garlic and Its Antifungal Activity against Wheat Fusarium Head Blight Pathogen Fusarium graminearum. Nanomaterials 2020, 10, 219. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Endophytic Bacterial Species | Host Plant/ Organ | Plant Growth Promotion Attributes | References |
|---|---|---|---|
| Proteobacteria: Pseudomonas spp. | Nicotiana tabacum/seeds | Siderophores, IAA, ACC deaminase production, nitrogen fixation, phosphorus/potassium solubilization, and trace metal tolerance | [14] |
| Firmicutes: Bacillus paralicheniformis | Rice (Oryza sativa L.)/roots | Nitrogen fixation | [15] |
| Firmicutes: Bacillus mojavensis, Bacillus sp. | Ammodendron bifolium/roots and leaves | IAA, ACC deaminase, amylase, cellulase, protease, lipase production, phosphate solubilization, nitrogen fixation | [16] |
| Proteobacteria: Aquabacterium, Duganella, Massilia, Bordetella, Salmonella, Pantoea, Kosakonia, Klebsiella, Serratia, Pseudomonas, Agrobacterium, Stenotrophomonas, Brevundimonas, Ancylobacter, Pleomorphomonas. Actinobacterium: Curtobacterium, Microbacterium, Nocardia, Sediminihabitans. Firmicutes: Bacillus, Micrococcus, Staphylococcus, Exiguobacterium | Sorghum bicolor/roots and stems | IAA production, fungicidal and bactericidal activities, nitrogen fixation | [17] |
| Proteobacteria: Acetobacter, Burkholderia, Caulobacter, Pseudomonas, Ralstonia, Bradyrhizobium, Methylocapsa | Pinus arizonica; Pinus durangensis/roots, phloem, and bark | Production of active secondary metabolites, metabolism of vitamins and cofactors | [18] |
| Actinobacteria: Streptomyces cavourensis | Cinnamomum cassia/roots | Biosynthesis of active compounds with antimicrobial and cytotoxic properties and plant growth-promoting capabilities. | [19] |
| Proteobacteria: Sphingomonas sp. | Tephrosia apollinea/leaves | Drought tolerance | [20] |
| Actinobacteria: Kocuria sp., Micrococcus luteus | Corchorus olitorius/leaves, roots, seeds, and seedling | IAA and siderophore production. | [21] |
| Firmicutes: Actinobacteria Proteobacteria: Curtobacterium sp., Microbacterium sp., Methylobacterium sp., Bacillus amyloliquefaciens | Browntop millet/seeds | Auxin production, phosphate solubilization, inhibiting fungal pathogens | [22] |
| Proteobacteria: Enterobacter ludwigii, Enterobacter spp., Agrobacterium tumefaciens, Kosakonia cowardii, Variovorax sp., Burkholderia spp., Pantoea vagans, Serratia marcescens Firmicutes: Bacillus sp. | Soybean/roots, stems, and leaves | Antagonistic activity against soybean pathogenic fungi and bacteria | [23] |
| Chryseobacterium endophyticum, Paenibacillus castaneae, Streptomyces sp., Lactobacillus plantarum, Bacillus proteolyticus, Pseudomonas sp., Serratia rubidaea, Klebsiella aerogenes, Paraburkholderia sp., Burkholderia sp., Bacillus cereus, Bacillus subtilis, Enterobacter cloacae, Enterobacter sp., Arthrobacter sp., Bacillus thuringiensis, Bacillus sp. | Pigeonpea/stems, roots, and leaves | Antimicrobial activity against Fusarium wilt (Fusarium udum) | [24] |
| Actinobacteria: Micrococcus yunnanensis | Avicennia marina/Propagule teguments | IAA, ammonium, siderophore, and protease production | [25] |
| 59 bacterial isolates belonging to phyla: Proteobacteria, Firmicutes, and Actinobacteria | Chickpea (Cicer arietinum L.)/roots | IAA production, ammonia production, cellulase production, salt tolerance | [26] |
| Firmicutes: Bacillus velezensis | Peanut/seeds | Antagonistic against Sclerotium rolfsii | [27] |
| Firmicutes: Bacillus subtilis | Sugarcane/leaves and stalks | Promoting plant growth, increasing N and chlorophyll content | [28] |
| Proteobacteria: Delftia, Stenotrophomonas; Rhizobium; Brevundimonas, Variovorax; Achromobacter; Novosphingobium; Comamonas; and Collimonas | Zea mays L., Vicia faba L., Secale cereale L., Triticum aestivum L., Arctium lappa L., and Equisetum arvense L./roots and stems | IAA and siderophore production, nitrogen fixation, and phosphate solubilization | [29] |
| Proteobacteria: Enterobacter tabaci, Pantoea agglomerans, Stenotrophomonas maltophilia, Sphingomonas sanguinis, Enterobacter tabaci | rice/seeds | IAA production and Cd tolerance | [30] |
| Actinobacteria: Streptomyces niveus NRRL 2466 | Camellia spp. and related genera/roots and leaves | IAA, Ammonia, siderophores, ACC deaminase, chitinase, and protease, production. N2 fixation, P solubilization | [31] |
| 138 endophytic bacterial strains belonging to the phyla Proteobacteria (Pseudomonadales, Burkholderiales, and Xanthomonadales) Firmicutes, and Bacteroidetes (Bacillales and Flavobacteriales) | Six terrestrial orchid species/roots | Phosphate solubilization, siderophore production, IAA production, antagonistic activities against plant pathogenic fungi | [32] |
| Herbaspirillum lusitanum (2 species), Acinetobacter johnsonii (3 species), Stenotrophomonas rhizophila, Agrobacterium tumefaciens (4 species), Rhizobium radiobacter, Micrococcus yunnanensis, Paenibacillus graminis, Bacillus pumilus (2 species), Bacillus cereus; Bacillus muralis (2 species), Terribacillus goriensis | Cucumber/roots, shoots, and leaves | IAA production, siderophore production, phosphate solubilization, antibiotic production, salt tolerance | [33] |
| Bacillus cereus, Pseudomonas migulae (3 species), Pseudomonas spp. (2 species), Pseudomonas brassicacearum, Paenibacillus lautus, Brevibacterium frigoritolerans, Bacillus anthracis, Paenibacillus illinoisensis, Bacillus muralis, Bacillaceae bacterium, Micrococcus luteus | Sorghum/roots | IAA production, siderophore production, phosphate solubilization, antibiotic production, salt tolerance | [33] |
| Bacillus safensis, Acinetobacter lwoffii, Bacillus cereus (6 species), Bacillus thuringiensis (4 species), Bacillus muralis (2 species), Bacillus megaterium, Bacillus tequilensis, Bacillus aerophilus, Bacillaceae bacterium (2 species), Acinetobacter johnsonii (2 species), Microbacterium schleiferi, Bacillus subtilis, Paenibacillus sp., Bacillus niacin, Kochuria palustris | Tomato/roots, shoots, and leaves | IAA production, siderophore production, phosphate solubilization, antibiotic production, salt tolerance | [33] |
| Firmicutes: Paenibacillus polymyxa | Lilium lancifolium/bulbs | IAA, siderophore, ACC deaminase, and organic acid production; nitrogen fixation; phosphate solubilization; antifungal activities against fungal phytopathogens | [34] |
| Firmicutes and proteobacteria: Actinobacteria; Bacillus, Fictibacillus, Lysinibacillus, Paenibacillus, Cupriavidus, and Microbacterium | Different rice cultivars such as Xiushui-48, Y-003, and CO-39/roots | Antagonistic effect against rice fungal phytopathogens | [35] |
| Paenibacillus barengoltzii (2 species), Bacillus amyloliquefaciens (2 species), Bacillus thuringiensis (2 species), Bacillus cereus (4 species) | Fagonia mollis/leaves | Enzymatic activities, IAA production, ammonia production, phosphate solubilization, antibiotic activities | [36] |
| Brevibacillus agri (3 species) | Achillea fragrantissima/leaves | Enzymatic activities, IAA production, ammonia production, phosphate solubilization, antibiotic activities | [36] |
| Hormone | Producer Strain | Plant Source | Function/Effect | Reference |
|---|---|---|---|---|
| Gibberellins | Bacillus amyloliquefaciens strain RWL-1 | Oryza sativa L. (Poales: Poaceae) | Plant growth promotion, hormone regulation | [62] |
| Abscisic acid | Azospirillum lipoferum | Maize | Alleviating drought stress symptoms in maize | [78] |
| Cytokinin’s | Bacillus subtilis | lettuce plants | Increased plant shoot and root weight by approximately 30% | [80] |
| Auxin (indol acetic acid) | B. amyloliquefaciens B. cereus and Bacillus subtilis | Capsicum annuum L. (Solanales: Solanaceae) | Anthracnose control, plant growth promotion, and biomass improvement | [81,82] |
| Auxins | B. amyloliquefaciens strain B14 and Bacillus sp. strains B19 and P12 | Phaseolus vulgaris | Plant growth promotion, seed germination | [83] |
| Indol acetic acid | B. subtilis strain NA-108 | Fragaria ananassa Duchesne (Rosales: Rosaceae) | Plant growth promotion and biomass improvement | [84] |
| IAA | Pseudomonas aeruginosa. Bradyrhizobium sp. | Soybean | Plant growth-promoting | [85] |
| IAA, gibberellins, and cytokinin | Acitenobacter braumalli, Enterobacter asburiae, Pseudomonas aeruginosa, Pseudomonas fulva, Pseudomonas lini; Pseudomonas montelli, Pseudomonas putida, Pseudomonas thivervalensis, Sinorhizobium meliloti, Klebsiella pneumoniae | Maize | Plant growth-promoting, alleviating drought stress, biocontrol activity | [86] |
| IAA | Acinetobacter guillouiae | Wheat | Plant growth-promoting; | [87] |
| IAA | Arthrobacter sulfonivorans | Wheat | Plant growth-promoting | [88] |
| IAA | Acinetobacter calcoaceticus, Bacillus amyloliquefaciens, Enterobacter cloaca, Pseudomonas putida | Soybean | [89] |
| Diazotrophic Endophytic Bacteria | Plant Source | Inoculated in | Capacity of N-Fixing Confirmed by | Reference |
|---|---|---|---|---|
| Proteobacteria: (Acinetobacter calcoaceticus, Enterobacter cloacae, Pseudomonas putida). Firmicutes: (Bacillus cereus, Bacillus amyloliquefaciens) | Glycine max L. | In vitro assay |
| [89] |
| Firmicutes: (Bacillus subtilis EB-04, Bacillus pumilus EB-64, Bacillus pumilus EB-169, Paenibacillus sp. EB-144) | Banana tree | In vitro assay |
| [104] |
| Actinobacteria (Arthrobacter), Proteobacteria (Rhizobium), Firmicutes (Bacillus spp.) | Diverse Poaceae family plants (maize, wheat, pearl millet, sorghum, and rice) | Wheat |
| [105] |
| Proteobacteria(Pseudomonas aeruginosa PM389) | Pennisetum glaucum | Wheat |
| [106] |
| Proteobacteria (Herbaspirillum sp.) | Tea plants (Camellia sinensis var. assamic and C. sinensis) | In vitro assay |
| [107] |
| Proteobacteria (Burkholderia spp., Klebsiella spp., Novosphingobium spp., Sphingomonas spp.) | Rice (Oryza sativa) | Rice (Oryza sativa) |
| [108] |
| Proteobacteria (Pseudomonas spp., Caballeronia sordidicola, Rhizobium herbae) Actinobacteria (Rathayibacter tanaceti, Frigoribacterium endophyticum, Herbiconiux solani) Bacteroidetes (Flavobacterium aquidurense) | Lodgepole pine (Pinus contorta var. latifolia) | In vitro assay; lodgepole pine (Pinus contorta) |
| [109] |
| Firmicutes(Paenibacillus kribbensis HS-R01, Paenibacillus kribbensis HS-R14) | Rice (Oryza sativa var. japonica) | Rice (Oryza sativa var. japonica) |
| [110] |
| Firmicutes (Bacillus spp.) Proteobacteria (Enterobacter sp.) | Zea mays L. | Zea mays L. |
| [111] |
| Firmicutes (Paenibacillus polymyxa P2b-2R) | Lodgepole pine (Pinus contorta var. latifolia) | Zea mays L. |
| [112] |
| Firmicutes (Paenibacillus polymyxa P2b-2R) | Lodgepole pine (Pinus contorta var. latifolia) | Canola (Brassica napus L.) and tomato (Solanum lycopersicum) |
| [113] |
| Firmicutes: (Bacillus spp, Paenibacillus spp.) Proteobacteria: (Caballeronia spp., Pseudomonas spp.) | Spruce tree | In vitro assay |
| [114] |
| Proteobacteria: (Burkholderia, Sphingomonas, Bradyrhizobium sp., Azospirillum brasilens, Rhodospirillum rubrum, Rhodobacter capsulatus) Cyanobacteria: (Nostoc punctiforme) Euryarchaeota: (Methanococcus maripaludis, Methanosarcina acetivoran) | Populus trichocarpa | In vitro assay |
| [115] |
| Proteobacteria: (Azospirillum amazonense AR3122, Burkholderia vietnamiensis AR1122) | Rice (Oryza sativa L) | Rice (Oryza sativa L.) |
| [116] |
| Proteobacteria: (Gluconacetobacter diazotrophicus, Azospirillum, Herbaspirillum seropedicae, Herbaspirillum rubrisubalbicans, Burkholderia tropica) | Sugarcane | Sugarcane |
| [117] |
| Proteobacteria: (Pseudomonas spp., Rhizobium spp., Duganella spp.) | Ageratina adenophora | In vitro assay |
| [118] |
| Stress Condition | Bacterial Endophytes Used | Plant Host | Effect/Mechanism of Resistance | References |
|---|---|---|---|---|
| Drought stress | Azospirillum spp. | Maize |
| [78] |
| Cold tolerance | Burkholderia phytofrmans PsJN | Grapevine plant |
| [159,160] |
| Drought stress | Burkholderia phytofirmans PsJN, Enterobacter sp. PsJN | Maize |
| [161] |
| Drought stress | Gluconacetobacter diazotrophicus | Sugarcane (Saccharum officinarum)/shoot |
| [162] |
| Drought stress | Pseudomonas azotoformans | Alyssum serpyllifolium/leaves |
| [163] |
| Drought stress | Bacillus amyloliquifaciens | Grapevine/roots |
| [164] |
| Drought stress | Pantoea alhagi | Alhagi sparsifolia/ leaves |
| [165] |
| Drought stress | Bacillus subtilis and Paenibacillus illinoinensi | Capsicum annuum/root |
| [166] |
| Drought stress | Bacillus pumilus | Glycyrrhiza uralensis |
| [167] |
| Drought stress | Bacillus sp. strain Acb9, Providencia sp. strain Acb11, Staphylococcus sp. strain Acb12, Staphylococcus sp. strain Acb13 and Staphylococcus sp. strain Acb14 | Ananas comosus |
| [168] |
| Drought stress | Sinorhizobium meliloti | Medicago sativa/root |
| [169] |
| Salinity | Pseudomonas pseudoalcaligenes | Rice |
| [170] |
| Salinity | Pseudomonas fluorescens YsS6 and P. migulae 8R6 | Tomato plants |
| [156] |
| Salinity and trace metals | Pseudomonas stutzeri A1501 | Rice |
| [171] |
| Salinity | Bacillus sp., Pantoea sp., Marinobacterium sp., Acinetobacter sp., Enterobacter sp., Pseudomonas sp., Rhizobium sp. and Sinorhizobium sp. | Psoralea corylifolia L. |
| [172] |
| Trace metal (copper-contaminated soils) | Pantoea agglomerans Jp3-3 and Achromobacter xylosoxidans strain Ax 10 | Brassica sp |
| [173,174] |
| Salinity | Bacillus subtilis strain BERA 71 | Acacia gerrardii Benth./root |
| [175] |
| Salinity | Curtobacterium oceanosedimentum strain SAK1, Curtobacterium luteum strain SAK2, Enterobacter ludwigii strain SAK5, Bacillus cereus strain SA1, Micrococcus yunnanensis strain SA2, Enterobacter tabaci SA3 | Oenothera biennis L., Artemisia princeps Pamp, Chenopodium ficifolium Smith, and Echinochloa crusgalli/roots |
| [176] |
| Salinity | Bacillus spp., Enterobacter spp. | Thymus vulgaris/leaves, stems, and roots |
| [177] |
| Trace metals (Cd, Zn, Pb, and Cu) | Mesorhizobium loti HZ76 and Agrobacterium radiobacter HZ6 | Robinia pseudoacacia/root nodules |
| [178] |
| Trace metal (Cd and Ni) | Enterobacter ludwigii strain SAK5 and Exiguobacterium indicum strain SA22 | - |
| [179] |
| Trace metal (Ni) | Stenotrophomonas sp. S20, Pseudomonas sp. P21, and Sphingobium sp. S42 | Tamarix chinensis |
| [180] |
| Strategy Used | Mechanisms | References |
|---|---|---|
| Competition for space and nutrients | Competitive root colonization, capacity to stick onto the root, differentiating the growth phase, efficacy to utilize the organic acids existing in the root exudates and hence synthesize different components | [206,207] |
| Competition with ferric iron |
| [196] |
| Detoxification of virulence factors |
| [54,208] |
| Antibiosis |
| [200,209,210,211,212] |
| Induced systemic resistance (ISR) |
| [213,214,215,216] |
| Nanoparticles | Bacterial Endophytes | Plant | Applications | References |
|---|---|---|---|---|
| Au | Pseudomonas veronii | Annona squamosa | Antibacterial | [247] |
| Au | Pseudomonas fluorescens 417 | Coffea arabica | Antibacterial | [248] |
| CaCl2 | Lysinibacillus xylanilyticus | Chiliadenus montanus | Degradation of cellulase | [249] |
| Cu | Streptomyces capillispiralis | Convolvulus arvensis | Antibacterial and antifungal | [245] |
| CuO | Streptomyces zaomyceticus and Streptomyces pseudogriseolus | Oxalis corniculata L. | Antimicrobial, antiphytopathogen, in vitro cytotoxicity, larvicidal activity | [246] |
| MgO | Streptomyces coelicolor | Ocimum sanctum | Active against multidrug-resistant microbes | [248] |
| ZnO | Sphingobacterium thalpophilum | Withania somnifera (L.) | Antimicrobial | [250] |
| Ag | Bacillus siamensis C1 | Coriandrum sativum | Antibacterial | [251] |
| Ag | Pantoea ananatis | Monocot plants | Anti-multidrug-resistant | [243] |
| Ag | Streptomyces laurentii R-1 | Achillea fragrantissima | Antibacterial, in vitro cytotoxicity, and textile industry | [252] |
| Ag | Streptomyces antimycotics L-1 | Mentha longifolia L. | Antibacterial, in vitro cytotoxicity, and textile industry | [253] |
| Ag | Pseudomonas poae CO | Allium sativum | Antifungal | [254] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eid, A.M.; Fouda, A.; Abdel-Rahman, M.A.; Salem, S.S.; Elsaied, A.; Oelmüller, R.; Hijri, M.; Bhowmik, A.; Elkelish, A.; Hassan, S.E.-D. Harnessing Bacterial Endophytes for Promotion of Plant Growth and Biotechnological Applications: An Overview. Plants 2021, 10, 935. https://doi.org/10.3390/plants10050935
Eid AM, Fouda A, Abdel-Rahman MA, Salem SS, Elsaied A, Oelmüller R, Hijri M, Bhowmik A, Elkelish A, Hassan SE-D. Harnessing Bacterial Endophytes for Promotion of Plant Growth and Biotechnological Applications: An Overview. Plants. 2021; 10(5):935. https://doi.org/10.3390/plants10050935
Chicago/Turabian StyleEid, Ahmed M., Amr Fouda, Mohamed Ali Abdel-Rahman, Salem S. Salem, Albaraa Elsaied, Ralf Oelmüller, Mohamed Hijri, Arnab Bhowmik, Amr Elkelish, and Saad El-Din Hassan. 2021. "Harnessing Bacterial Endophytes for Promotion of Plant Growth and Biotechnological Applications: An Overview" Plants 10, no. 5: 935. https://doi.org/10.3390/plants10050935