
Evaluation and Selection of Interspecific Lines of Groundnut (Arachis hypogaea L.) for Resistance to Leaf Spot Disease and for Yield Improvement

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Leaf Spots
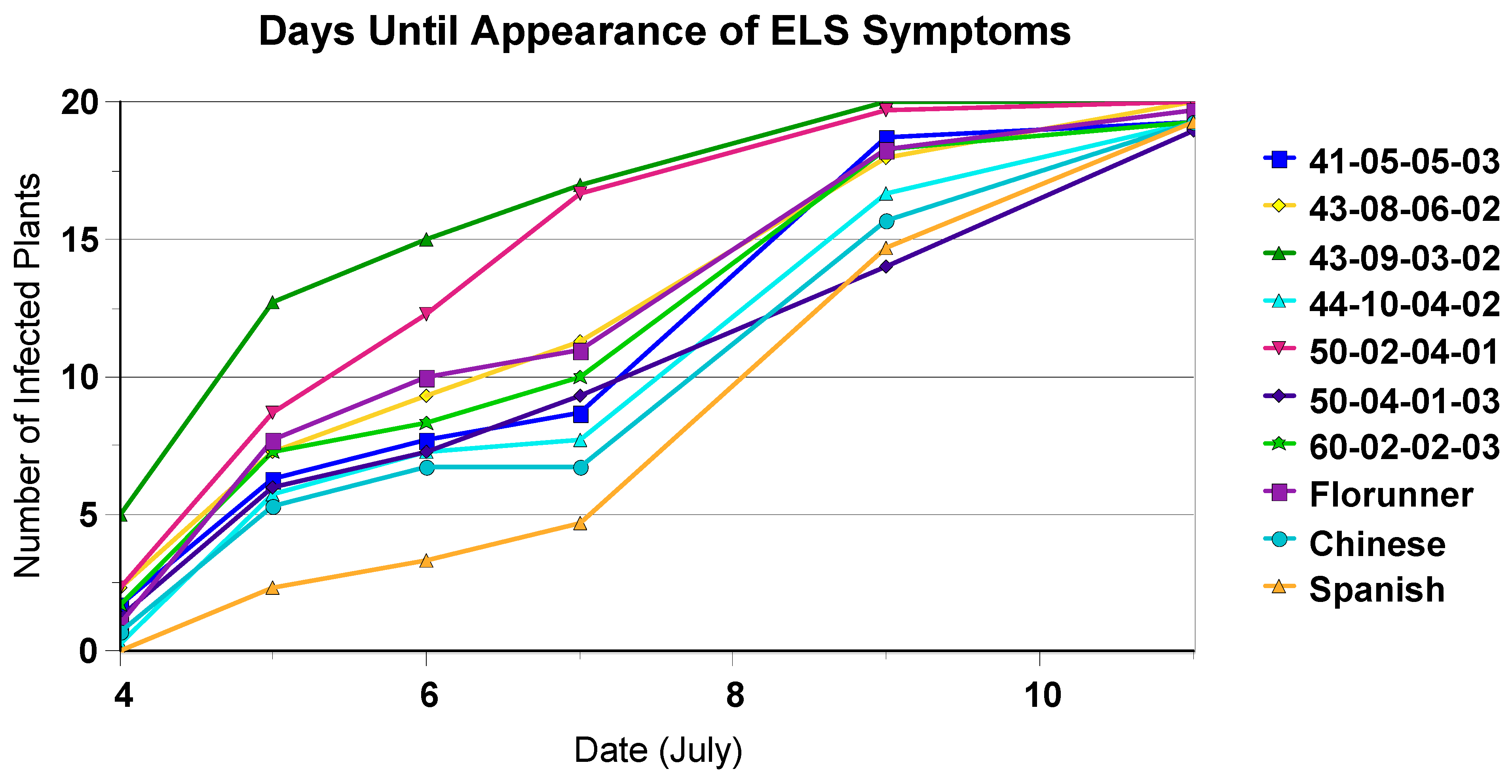
2.2. Time to Appearance of Symptoms
2.3. Pod Yield
2.4. Correlations
3. Discussion
3.1. Test of Hypotheses
3.1.1. Introgression
3.1.2. Breaking Linkage Drag
3.2. Selections
3.3. Early and Late Leaf Spot Ratings
3.4. Time to Appearance of Symptoms and Disease Incidence
3.5. Correlations
3.6. Utility of the AB-QTL Method
3.7. Continuing Work
4. Materials and Methods
4.1. Plant Materials
4.2. Field Plot Design and Operations
4.3. Defoliation Ratings
4.4. Time to Appearance of Symptoms
4.5. Pod Yield
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shokes, F.M.; Culbreath, A.K. Early and late leaf spots. In Compendium of Peanut Diseases, 2nd ed.; Kokalis-Burelle, N., Porter, D.M., Rodriguez-Kabana, R., Smith, D.H., Subrahmanyam, P., Eds.; APS Press: Saint Paul, MN, USA, 1997; pp. 17–20. [Google Scholar]
- Smith, D.H. Early and late leaf spots. In Compendium of Peanut Diseases; Porter, D.M., Smith, D.H., Rodriguez-Kabana, R., Eds.; APS Press: Saint Paul, MN, USA, 1984. [Google Scholar]
- McDonald, D.; Subrahmanyam, P.; Gibbons, R.W.; Smith, D.H. Early and Late Leaf Spot of Groundnut; Information Bulletin No. 21; International Crops Research Institute for the Semi-Tropics: Patancheru, India, 1985; p. 24. [Google Scholar]
- Waliyar, F. Evaluation of yield losses due to groundnut leaf diseases in West Africa. In Proceedings of the Summary Proceedings of the Second ICRISAT Regional Groundnut Meeting for West Africa, Niamey, Niger, 11–14 September 1990; ICRISAT: Patancheru, India, 1991; pp. 32–33. [Google Scholar]
- Waliyar, F.; Adamou, M.; Traoré, A. Rational Use of Fungicide Applications to Maximize Peanut Yield Under Foliar Disease Pressure in West Africa. Plant Dis. 2000, 84, 1203–1211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wynne, J.C.; Beute, M.K.; Nigam, S.N. Breeding for Disease Resistance in Peanut (Arachis Hypogaea L.). Annu. Rev. Phytopathol. 1991, 29, 279–303. [Google Scholar] [CrossRef]
- Coffelt, T.A.; Porter, D.M. Field Screening of Reciprocal Chico x Florigiant Peanut Populations for Resistance to Leafspot in Virginia1. Peanut Sci. 1986, 13, 57–60. [Google Scholar] [CrossRef]
- Naab, J.B.; Tsigbey, F.K.; Prasaad, P.V.V.; Boote, K.J.; Bailey, J.E.; Brandenburg, R.L. Effects of sowing date and fungicide application on yield of early and late maturing peanut cultivars under rainfed conditions in Ghana. Crop Prot. 2005, 24, 325–332. [Google Scholar] [CrossRef]
- Kokalis-Burelle, N.; Backman, P.A.; Rodríguez-Kábana, R.; Ploper, L.D. Potential for biological control of early leafspot of peanut using Bacillus cereus and chitin as foliar amendments. Biol. Control. 1992, 2, 321–328. [Google Scholar] [CrossRef]
- Kishore, G.K.; Pande, S.; Podile, A.R. Management of late leaf spot of Groundnut (Arachis hypogaea) with chlorothalonil-tolerant isolates of Pseudomonas aeruginosa. Plant Pathol. 2015, 54, 401–408. [Google Scholar] [CrossRef]
- Nana, T.A. Essai de Lutte Intégrée par Association de la lutte Génétique et de la Lutte Biologique pour le Contrôle de la Rouille et des Cercosporioses de L’arachide (Arachis hypogaea L.) au Burkina Faso. Bachelor’s Thesis, Université de Ouagadougou, Ouagadougou, Burkina Faso, 2015; 169p. [Google Scholar]
- Abudulai, M.; Naab, J.; Seini, S.S.; Dzomeku, I.; Boote, K.; Brandenburg, R.; Jordan, D. Peanut (Arachis hypogaea) response to weed and disease management in northern Ghana. Int. J. Pest Manag. 2017, 64, 204–209. [Google Scholar] [CrossRef]
- Sanon, E.; Dao, K.; Neya, F.B.; Zagré, B.; Burow, M.; Sankara, P. Essai de Lutte Intégrée Combinant Les Pratiques Culturales, la Résistance Variétale et Chlorothalonil Pour le Contrôle des Cercosporioses de l’arachide au Burkina Faso; Annale de l’Université Joseph KI-ZERBO Série C: Ouagadougou, Burkina Faso, 2019; Volume 15. [Google Scholar]
- Smartt, J.; Stalker, H.T. Speciation and cytogenetics in Arachis. In Peanut Science and Technology; Pattee, H.E., Young, C.T., Eds.; American Peanut Research and Education Society: Yoakum, TX, USA, 1982; pp. 21–49. [Google Scholar]
- Abdou, Y.A.-M.; Gregory, W.C.; Cooper, W.E. Sources and nature of resistance to Cercospora arachidicola Hori and Cercosporidium personatum (Berk & Curtis) Deighton in Arachis species. Peanut. Sci. 1974, 1, 6–11. [Google Scholar]
- Stalker, H.; Moss, J. Speciation, Cytogenetics, and Utilization of Arachis Species. Adv. Agron. 1987, 41, 1–40. [Google Scholar] [CrossRef] [Green Version]
- Simpson, C.E.; Nelson, S.C.; Starr, J.L.; Woodard, K.E.; Smith, O.D. Registration of TxAG-6 and TxAG-7 Peanut Germplasm Lines. Crop Sci. 1993, 33, 1418. [Google Scholar] [CrossRef]
- Smartt, J.; Gregory, W.C. Interspecific cross-compatibility between the cultivated peanut Arachis hypogaea L. and other members of the genus Arachis. Oléagineux 1967, 22, 455–459. [Google Scholar]
- Stalker, H.T.; Wynne, J.C.; Company, M. Variation in progenies of an Arachis hypogaea x diploid wild species hybrid. Euphytica 1979, 28, 675–684. [Google Scholar] [CrossRef]
- Garcia, G.M.; Stalker, H.T.; Shroeder, E.; Kochert, G. Identification of RAPD, SCAR, and RFLP markers tightly linked to nematode resistance genes introgressed from Arachis cardenasii into Arachis hypogaea. Genome 1996, 39, 836–845. [Google Scholar] [CrossRef]
- Smartt, J.; Gregory, W.C.; Gregory, M.P. The genomes of Arachis hypogaea 2. The implications in interspecific breeding. Euphytica 1978, 27, 677–680. [Google Scholar] [CrossRef]
- Norden, A.J.; Lipscomb, R.W.; Carver, W.A. Registration of Florunner Peanuts 1 (Reg. No. 2). Crop. Sci. 1969, 9, 850. [Google Scholar] [CrossRef]
- Simpson, C.E.; Starr, J.L. Registration of ‘COAN’ Peanut. Crop Sci. 2001, 41, 918. [Google Scholar] [CrossRef]
- Simpson, C.E.; Starr, J.L.; Church, G.T.; Burow, M.D.; Paterson, A.H. Registration of ‘NemaTAM’ Peanut. Crop Sci. 2003, 43, 1561. [Google Scholar] [CrossRef]
- Holbrook, C.C.; Timper, P.; Culbreath, A.K.; Kvien, C.K. Registration of ‘Tifguard’ peanut. J. Plant Regist. 2008, 2, 92–94. [Google Scholar] [CrossRef]
- Simpson, C.E.; Starr, J.L.; Baring, M.R.; Burow, M.D.; Cason, J.M.; Wilson, J.N. Registration of ‘Webb’ peanut. J. Plant Regist. 2013, 7, 265–268. [Google Scholar] [CrossRef]
- Holbrook, C.C., Jr.; Ozias-Akins, P.; Chu, Y.; Culbreath, A.K.; Kvien, C.K.; Brenneman, T.B. Registration of ‘TifNV-High O/L’ peanut. J. Plant Regist. 2017, 11, 228–230. [Google Scholar] [CrossRef]
- Branch, W.D.; Brenneman, T.B. Registration of ‘Georgia-14N’ Peanut. J. Plant Regist. 2015, 9, 159–161. [Google Scholar] [CrossRef]
- Burow, M.D.; Simpson, C.E.; Paterson, A.H.; Starr, J.L. Identification of peanut (Arachis hypogaea L.) RAPD markers diagnostic of root-knot nematode (Meloidogyne arenaria) (Neal) Chitwood) resistance. Mol. Breed. 1996, 2, 369–379. [Google Scholar] [CrossRef]
- Church, G.T.; Simpson, C.E.; Burow, M.D.; Paterson, A.H.; Starr, J.L. Use of RFLP markers for identification of individuals homozygous for resistance to Meloidogyne arenaria in peanut. Nematology 2000, 2, 575–580. [Google Scholar] [CrossRef]
- Chu, Y.; Holbrook, C.C.; Timper, P.; Ozias-Akins, P. Development of a PCR-Based Molecular Marker to Select for Nematode Resistance in Peanut. Crop. Sci. 2007, 47, 841–845. [Google Scholar] [CrossRef] [Green Version]
- Clevenger, J.; Chu, Y.; Guimaraes, L.A.; Maia, T.; Bertioli, D.; Leal-Bertioli, S.; Timper, P.; Holbrook, C.C.; Ozias-Akins, P. Gene expression profiling describes the genetic regulation of Meloidogyne arenaria resistance in Arachis hypogaea and reveals a candidate gene for resistance. Sci. Rep. 2017, 7, 1317. [Google Scholar] [CrossRef]
- Tanksley, S.D.; Nelson, J.C. Advanced backcross QTL analysis: A method for the simultaneous discovery and transfer of valuable QTLs from unadapted germplasm into elite breeding lines. Theor. Appl. Genet. 1996, 92, 191–203. [Google Scholar] [CrossRef] [PubMed]
- Burow, M.D.; Starr, J.L.; Park, C.-H.; Simpson, C.E.; Paterson, A.H. Introgression of homoeologous quantitative trait loci (QTLs) for resistance to the root-knot nematode [Meloidogyne arenaria (Neal) Chitwood] in an advanced backcross-QTL population of peanut (Arachis hypogaea L.). Mol. Breed. 2014, 34, 393–406. [Google Scholar] [CrossRef]
- Kornegay, J.L.; Beute, M.K.; Wynne, J.C. Inheritance of Resistance to Cercospora arachidicola and Cercosporidium personatum in Six Virginia-type Peanut Lines1,3. Peanut Sci. 1980, 7, 4–9. [Google Scholar] [CrossRef]
- Jogloy, S.; Wynne, J.C.; Beute, M.K. Inheritance of Late Leafspot Resistance and Agronomic Traits in Peanut1. Peanut Sci. 1987, 14, 86–90. [Google Scholar] [CrossRef]
- Anderson, W.F.; Holbrook, C.C.; Wynne, J.C. Heritability and Early-Generation Selection for Resistance to Early and Late Leafspot in Peanut. Crop. Sci. 1991, 31, 588–593. [Google Scholar] [CrossRef]
- Green, C.C.; Wynne, J.C. Field and greenhouse evaluations of the components of partial resistance to early leafspot in peanut. Euphytica 1986, 35, 561–573. [Google Scholar]
- Anderson, W.F.; Holbrook, C.C.; Brenneman, T.B. Resistance to Cercosporidium Personatum Within Peanut Germplasm1. Peanut Sci. 1993, 20, 53–57. [Google Scholar] [CrossRef] [Green Version]
- Waliyar, F.; McDonald, D.W.; Rao, P.V.S.; Reddy, P.M. Components of Resistance to an Indian Source of Cercospora Arachidicola in Selected Peanut Lines1. Peanut Sci. 1993, 20, 93–96. [Google Scholar] [CrossRef] [Green Version]
- Waliyar, F.B.; Shew, B.; Shidamed, R.; Beute, M.K. Effects of host resistance on germination of Cercospora arachidicola on peanut leaf surfaces. Peanut Sci. 1995, 22, 154–157. [Google Scholar] [CrossRef] [Green Version]
- Iroume, R.N.; Knauft, D.A. Heritabilities and correlations for early leafspot resistance in peanut (Arachis hypogaea L.): Implications for early generation selection. Peanut Sci. 1987, 14, 46–50. [Google Scholar] [CrossRef]
- Khedikar, Y.P.; Gowda, M.V.C.; Sarvamangala, C.; Patgar, K.V.; Upadhyaya, H.D.; Varshney, R.K. A QTL study on late leaf spot and rust revealed one major QTL for molecular breeding for rust resistance in groundnut (Arachis hypogaea L.). Theor. Appl. Genet. 2010, 121, 971–984. [Google Scholar] [CrossRef] [Green Version]
- Clevenger, J.; Chu, Y.; Chavarro, C.; Agarwal, G.; Bertioli, D.J.; Leal-Bertioli, S.C.; Pandey, M.K.; Vaughn, J.; Abernathy, B.; Barkley, N.A.; et al. Genome-wide SNP Genotyping Resolves Signatures of Selection and Tetrasomic Recombination in Peanut. Mol. Plant 2017, 10, 309–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isleib, T.G.; Milla-Lewis, S.R.; Pattee, H.E.; Copeland, S.C.; Zuleta, M.C.; Shew, B.B.; Hollowell, J.E.; Sanders, T.H.; Dean, L.O.; Hendrix, K.W.; et al. Registration of ‘Bailey’ peanut. J. Plant Regist. 2011, 5, 27–39. [Google Scholar] [CrossRef]
- Padi, F.K.; Frimpong, A.; Kombiok, J.; Salifu, A.B.; Marfo, K.O. Registration of ‘Nkatiesari’ Peanut. Crop Sci. 2006, 46, 1397–1398. [Google Scholar] [CrossRef]
- Leal-Bertioli, S.C.M.; José, A.C.V.F.; Alves-Freitas, D.M.T.; Moretzson, M.C.; Guimarães, P.M.; Nielsen, S.; Vidigal, B.S.; Pereira, R.W.; Pike, J.; Fávero, A.P.; et al. Identification of candidate genome regions controlling disease resistance in Arachis. BMC Plant Biol. 2009, 9, 112. [Google Scholar] [CrossRef] [Green Version]
- Burow, M.D.; Simpson, C.E.; Starr, J.L.; Paterson, A.H. Transmission genetics of chromatin from a synthetic amphidiploid in cultivated peanut (Arachis hypogaea L.): Broadening the gene pool of a monophyletic polyploid species. Genetics 2001, 159, 823–837. [Google Scholar] [PubMed]
- Burow, M.D.; Starr, J.L.; Park, H.; Simpson, C.E.; Paterson, A.H. Identification of QTLs for resistance to early leafspot (Cercospora arachidicola S. Hori.) in an introgression population of peanut (Arachis hypogaea L.). In Proceedings of the Plant & Animal Genome XVI Conference, San Diego, CA, USA, 14 January 2008. [Google Scholar]
- Burow, M.D.; M’Bi Zagré, B.; Denwar, N.N.; Sankara, P.; Neya, F.; Sanon, E.; Baring, M.R.; Grichar, J.; Simpson, C.E. Evaluation of a peanut interspecific introgression population for resistance to leafspot and rust, and for yield and maturity. (in preparation)
- Branch, W.D.; Brenneman, T.B.; Noe, J.P. Evidence for a second RKN resistance gene in peanut. Peanut Sci. 2016, 43, 49–51. [Google Scholar] [CrossRef]
- Wynne, J.C.; Halward, T. Cytogenetics and genetics of Arachis. Crit. Rev. Plant Sci. 1989, 8, 189–220. [Google Scholar] [CrossRef]
- Nigam, D.J. A review of the present status of the genetic resources of the ICRISAT Regional Groundnut Improvement Program, of the Southern African Cooperative Regional Yield Trials, and of rosette virus resistance breeding. In Proceedings of the Second Regional Groundnut Workshop for Southern Africa. Harare, Zimbabwe, 10–14 February 1986, 2nd ed.; International Crops Research Institute for the Semi-Arid Tropics: Patancheru, Andhra Pradesh, India, 1986; pp. 15–30. [Google Scholar]
- Ouedraogo, M. Yield and Leafspot Responses of Interspecific Peanut Crosses in Early Generation Tests. Ph.D. Thesis, Soil and Crop Science, Department, Texas A.&M University, College Station, TX, USA, 1995; 186p. [Google Scholar]
- Pande, S.; Rao, J.N.; Dwivedi, S.L. Components of resistance to late leaf spot caused by Phaeoisariopsis personata in inter-specific derivatives of groundnut. Indian Phytopath. 2002, 55, 444–450. [Google Scholar]
- Tuggle, J.C. Selection of Agronomically Acceptable, Early Leaf Spot (Cercospora arachidicola Hori.) Resistant Peanut (Arachis hypogaea L.) Breeding Lines in Interspecific Derived Populations and Implications of Pathogen Variability on Stability of Breeding Programs. Ph.D. Thesis, Soil and Crop Science Department, Texas A&M University, College Station, TX, USA, 1998; 109p. [Google Scholar]
- Chiteka, Z.A.; Gorbet, D.W.; Knauft, D.A.; Shokes, F.M.; Kucharek, T.A. Components of resistance to late leafspot in peanut: I. Levels and variability: Implications for selection. Peanut Sci. 1988, 15, 25–30. [Google Scholar] [CrossRef]
- Chiteka, Z.A.; Gorbet, D.W.; Knauft, D.A.; Shokes, F.M.; Kucharek, T.A. Components of Resistance to Late Leafspot in Peanut. II. Correlations Among Components and Their Significance in Breeding for Resistance1. Peanut Sci. 1988, 15, 76–81. [Google Scholar] [CrossRef]
- Dwivedi, S.L.; Pande, S.; Rao, J.N.; Nigam, S.N. Components of resistance to late leafspot and rust among interspecific derivatives and their significance in foliar disease resistance breeding in groundnut (Arachis hypogaea L.). Euphytica 2002, 125, 81–88. [Google Scholar] [CrossRef]
- Xiao, J.; Li, J.; Grandillo, S.; Ahn, S.N.; Yuan, L.; Tanksley, S.D.; McCouch, S.R. Identification of trait-improving quantitative trait loci alleles from a wild rice relative. Oryza rufipogon. Genetics 1998, 150, 899–909. [Google Scholar]
- Wilson, J.N.; Chopra, R.; Baring, M.R.; Selvaraj, M.G.; Simpson, C.E.; Chagoya, J.; Burow, M.D. Advanced backcross quantitative trait loci (QTL) analysis of oil concentration and oil quality traits in peanut (Arachis hypogaea L.). Trop. Plant Biol. 2016, 10, 1–17. [Google Scholar] [CrossRef]
- Tengey, T.K.; Hillhouse, A.; Mendu, V.; Burow, M.D. Analysis of a BC3F6 interspecific peanut introgression population using genome-specific SNP markers. Proc. Am. Peanut Res. Edu. Soc. 2018, in press. [Google Scholar]
- Tengey, T.K. Genetic Mapping of Leafspot Resistant QTLs, and Introgression into West African Adapted and US-high Oleic peanuts. Ph.D. Thesis, Texas Tech University, Department of Plant and Soil Science, Lubbock, TX, USA, 2018. [Google Scholar]
- Kombiok, J.M.; Buah, S.S.J.; Dzomeku, I.K.; Abdulai, H. Sources of pod yield losses in groundnut in the Northern Savanna Zone of Ghana. W. Afr. J. Appl. Ecol. 2004, 20, 53–63. [Google Scholar]
- Clavel, D.; Sarr, B.; Marone, E.; Ortiz, R. Potential agronomic and physiological traits of Spanish groundnut varieties (Arachis hypogaea L.) as selection criteria under end-of-cycle drought conditions. Agronomie 2004, 24, 101–111. [Google Scholar] [CrossRef] [Green Version]
- Shaner, G.; Finney, R.E. The effect of nitrogen fertilization on the expression of slow-mildewing resistance in Knox wheat and barley. Plant Dis. 1977, 71, 927–930. [Google Scholar] [CrossRef] [Green Version]
- Steele, R.G.D.; Torrie, J.H. Principles and Procedures of Statistics: A Biometric Approach, 2nd ed.; McGraw-Hill: New York, NY, USA, 1989. [Google Scholar]



{kind=link}
{kind=link}
| Accession | Mean 1 | 2006 Manga | 2006 Nyankpala | 2007 Manga | 2007 Nyankpala | 2005 Yoakum | N MFG 2 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Spanish | 287 | a 3 | 225 | a | 360 | a | 400 | ns 4 | 163 | a-d | n.d. 5 | 0 | |
| Chinese | n.d. | 225 | a | 320 | b | n.d. | n.d. | n.d. | 0 | ||||
| 55-437 | 267 | b | 216 | a–c | 315 | bc | 370 | 167 | ab | n.d. | 0 | ||
| 26-10-06-03 | 260 | bc | 215 | a–c | 285 | c–e | 375 | 165 | a–c | 7.33 | a–e | 0 | |
| 45-03-06-01 | 258 | b–d | 225 | a | 280 | de | 375 | 152 | a–h | 7.16 | a–e | 0 | |
| 31-04-10-03 | 257 | b–d | 225 | a | 295 | b–d | 360 | 147 | b–h | 6.00 | c–f | 0 | |
| 31-04-05-02 | 256 | b–d | 205 | b–d | 295 | b–d | 370 | 155 | a–f | 7.00 | a–e | 0 | |
| 63-04-02-03 | 256 | b–d | 210 | a–d | 295 | b–d | 375 | 142 | c–h | 7.66 | a–d | 0 | |
| 63-04-02-02 | 256 | b–d | 210 | a–d | 275 | d–f | 380 | 157 | a–f | 6.66 | a–f | 0 | |
| 50-04-03-03 | 253 | b–d | 215 | a–c | 255 | e–g | 380 | 163 | a–d | 8.33 | ab | 0 | |
| 45-04-02-01 | 252 | c–e | 225 | a | 280 | de | 375 | 128 | h | 4.83 | fg | 2 | |
| 44-10-04-02 | 252 | c–e | 200 | cd | 275 | d–f | 360 | 173 | a | 7.50 | a–d | 1 | |
| 70-03-07-01 | 252 | c–e | 225 | a | 265 | d–f | 380 | 137 | e–h | 5.66 | d–g | 2 | |
| 41-10-03-03 | 250 | c–f | 200 | cd | 280 | de | 355 | 167 | a–b | 7.33 | a–e | 1 | |
| 63-06-08-02 | 250 | c–f | 220 | ab | 270 | d–f | 350 | 160 | a–e | 8.33 | ab | 0 | |
| Florunner | 250 | c–f | 215 | a–c | 265 | d–f | 355 | 164 | a–c | 7.67 | a–d | 0 | |
| 46-06-04-01 | 250 | c-f | 210 | a–d | 260 | ef | 375 | 153 | a–g | 8.17 | ab | 1 | |
| 46-06-07-02 | 249 | c–g | 220 | ab | 265 | d–f | 370 | 142 | c–h | 8.17 | ab | 1 | |
| 41-10-01-03 | 249 | c–h | 215 | a–c | 265 | d–f | 360 | 155 | a–f | 8.00 | a–c | 0 | |
| 43-09-02-03 | 248 | c–h | 225 | a | 270 | d–f | 350 | 145 | b–h | 5.83 | d–f | 1 | |
| 63-04-05-02 | 247 | c–h | 220 | ab | 270 | d–f | 350 | 149 | a–h | 7.33 | a–e | 1 | |
| 43-09-03-01 | 246 | c–h | 220 | ab | 260 | ef | 375 | 128 | h | 5.83 | d–f | 1 | |
| 44-10-01-02 | 245 | d–h | 215 | a–c | 275 | d–f | 350 | 141 | c–h | 5.67 | d–g | 1 | |
| 19-05-06-03 | 245 | d–h | 204 | b–d | 255 | e–g | 375 | 146 | b–h | 8.00 | a–c | 3 | |
| 43-08-06-02 | 245 | d–h | 210 | a–d | 265 | d–f | 365 | 139 | d–h | 8.50 | a | 1 | |
| 41-05-05-03 | 244 | d–h | 195 | d | 255 | e–g | 375 | 150 | a–h | 4.91 | fg | 4 | |
| 43-09-03-02 | 239 | e–i | 210 | a–d | 255 | e–g | 360 | 130 | gh | 5.33 | e–g | 4 | |
| 60-02-02-03 | 236 | f–i | 205 | b–d | 255 | e–g | 335 | 150 | a–h | 4.67 | fg | 4 | |
| 43-08-07-02 | 235 | g–i | 220 | ab | 245 | fg | 335 | 139 | d–h | 3.67 | g | 3 | |
| 50-04-01-03 | 235 | hi | 220 | ab | 225 | g | 360 | 134 | f–h | 8.67 | a | 2 | |
| 50-02-04-01 | 228 | i | 210 | a–d | 225 | g | 330 | 147 | b–h | 6.33 | b–f | 3 | |
| p | 0.0044 | 0.0070 | 0.0001 | 0.2056 | 0.0207 | 0.0001 | |||||||
| Mean | 250 | 215 | 273 | 364 | 150 | 6.82 | |||||||
| LSD | 15 | 17 | 32 | 24 | 2.02 | ||||||||
| CV | 7.6% | 4.7% | 7.2% | 6.5% | 9.2% | 15.1% | |||||||
| Accession | Mean | 2006 Manga | 2006 Nyankpala | 2007 Manga | 2007 Nyankpala | N MFG 1 | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Spanish | 238 | ns 2 | 195 | a 3 | 380 | a | 260 | ns | 118 | a–d | 0 |
| Chinese | n.d. 4 | 195 | a | 340 | b | n.d. | n.d. | 0 | |||
| 55-437 | 227 | 185 | a–c | 345 | b | 250 | 129 | ab | 0 | ||
| 45-04-02-01 | 213 | 195 | a | 290 | c–e | 275 | 92 | hi | 0 | ||
| 26-10-06-03 | 210 | 185 | a–c | 280 | c–g | 255 | 121 | a–d | 0 | ||
| 31-04-10-03 | 210 | 190 | ab | 280 | c–g | 255 | 114 | a–f | 0 | ||
| 43-09-02-03 | 209 | 195 | a | 290 | c–e | 240 | 110 | b–h | 0 | ||
| 45-03-06-01 | 207 | 185 | a–c | 275 | c–h | 260 | 109 | b–i | 2 | ||
| 41-10-01-03 | 207 | 175 | b–e | 290 | c–e | 250 | 113 | a–g | 1 | ||
| 70-03-07-01 | 207 | 185 | a–c | 260 | e–h | 270 | 113 | a–g | 1 | ||
| 63-06-08-02 | 206 | 185 | a–c | 285 | c–f | 235 | 120 | a–d | 1 | ||
| 31-04-05-02 | 206 | 170 | c–e | 305 | c | 235 | 115 | a–f | 0 | ||
| 41-10-03-03 | 205 | 165 | de | 270 | d–h | 255 | 131 | a | 2 | ||
| 44-10-01-02 | 204 | 180 | a–d | 295 | cd | 235 | 107 | c–i | 1 | ||
| 63-04-02-02 | 204 | 170 | c–e | 270 | d–h | 260 | 114 | a–f | 2 | ||
| 63-04-02-03 | 203 | 175 | b–e | 290 | c–e | 240 | 109 | c–i | 2 | ||
| 46-06-07-02 | 203 | 185 | a–c | 260 | e–h | 255 | 114 | a–f | 1 | ||
| 63-04-05-02 | 202 | 180 | a–d | 280 | c–g | 240 | 107 | c–i | 1 | ||
| 46-06-04-01 | 202 | 165 | de | 265 | d–h | 255 | 122 | a–d | 2 | ||
| 44-10-04-02 | 201 | 165 | de | 280 | c–g | 235 | 125 | a–c | 1 | ||
| 43-08-06-02 | 201 | 180 | a–d | 275 | c–h | 245 | 103 | d–i | 2 | ||
| 50-02-04-01 | 200 | 180 | a–d | 260 | e–h | 255 | 104 | d–i | 2 | ||
| 50-04-03-03 | 199 | 175 | b–e | 250 | gh | 260 | 112 | a–h | 2 | ||
| 41-05-05-03 | 198 | 160 | e | 265 | d–h | 250 | 119 | a–d | 2 | ||
| 19-05-06-03 | 197 | 167 | de | 265 | d–h | 250 | 107 | c–i | 3 | ||
| 43-09-03-01 | 196 | 185 | a–c | 255 | f–h | 250 | 96 | f–i | 2 | ||
| 50-04-01-03 | 195 | 190 | ab | 245 | h | 250 | 93 | g–i | 2 | ||
| Florunner | 193 | 170 | c–e | 260 | e–h | 225 | 118 | a–d | 2 | ||
| 43-08-07-02 | 193 | 175 | b–e | 255 | f–h | 245 | 97 | e–i | 3 | ||
| 43-09-03-02 | 191 | 175 | b–e | 260 | e–h | 240 | 89 | i | 3 | ||
| 60-02-02-03 | 190 | 170 | c–e | 250 | gh | 230 | 110 | b–h | 2 | ||
| p | 0.1024 | 0.0003 | 0.0001 | 0.7600 | 0.0106 | ||||||
| Mean | 204 | 179 | 280 | 249 | 111 | ||||||
| LSD | 17 | 33 | 21 | ||||||||
| CV | 12.1% | 5.7% | 7.2% | 9.1% | 10.4% | ||||||
| Accession | Mean | Nyankapala 2006 | Manga 2007 | Nyankpala 2007 | N MFG 1 | ||||
|---|---|---|---|---|---|---|---|---|---|
| 55-437 | 36.30 | a 2 | 34.42 | a | 47.17 | ab | 27.33 | a-c | 3 |
| Spanish | 36.07 | ab | 31.09 | bc | 47.37 | ab | 29.76 | a | 2 |
| Chinese | n.d.3 | 30.29 | b–g | n.d. | n.d. | 1 | |||
| 50-04-01-03 | 34.79 | a–d | 30.43 | b–g | 44.10 | bc | 29.85 | a | 1 |
| 45-04-02-01 | 34.06 | a–e | 30.11 | b–h | 43.10 | cd | 28.96 | a | 1 |
| 50-04-03-03 | 33.87 | b–f | 30.91 | b–e | 42.63 | c–e | 28.07 | a–c | 1 |
| 43-09-03-02 | 33.71 | c–f | 27.64 | l | 44.03 | bc | 29.46 | a | 1 |
| 43-08-06-02 | 33.68 | c–g | 29.30 | d–l | 43.13 | cd | 28.62 | a | 1 |
| 43-08-07-02 | 33.60 | c–g | 31.17 | b | 41.37 | c–f | 28.27 | a–c | 1 |
| 26-10-06-03 | 33.50 | c–g | 29.08 | f–l | 44.10 | bc | 27.31 | a–d | 1 |
| 41-05-05-03 | 33.42 | d–g | 30.02 | b–i | 40.83 | c–f | 29.40 | a | 1 |
| 45-03-06-01 | 33.24 | d–g | 29.69 | b–j | 41.90 | c–e | 28.13 | a–c | 1 |
| 63-06-08-02 | 33.20 | d–g | 28.71 | g–l | 43.70 | bc | 27.20 | a–e | 1 |
| 46-06-04-01 | 33.18 | d–g | 31.04 | b–d | 39.83 | d–g | 28.65 | a | 1 |
| Florunner | 33.15 | d–g | 29.33 | c–l | 42.07 | c–e | 28.07 | a–c | 1 |
| 31-04-05-02 | 32.98 | d–g | 29.36 | c–l | 40.90 | c–f | 28.69 | a | 1 |
| 63-04-05-02 | 32.98 | d–g | 30.11 | b–h | 40.43 | c–f | 28.40 | ab | 1 |
| 31-04-10-03 | 32.94 | d–g | 29.27 | d–l | 41.60 | c–f | 27.94 | a–c | 1 |
| 60-02-02-03 | 32.92 | d–g | 29.44 | b–k | 40.77 | c–f | 28.57 | ab | 1 |
| 46-06-07-02 | 32.92 | d–g | 29.38 | c–l | 40.93 | c–f | 28.44 | ab | 1 |
| 44-10-04-02 | 32.92 | d–g | 30.01 | b–i | 41.37 | c–f | 27.38 | a–c | 1 |
| 63-04-02-03 | 32.91 | d–g | 29.80 | b–i | 40.67 | c–f | 28.25 | a–c | 1 |
| 50-02-04-01 | 32.79 | d–g | 28.30 | i–l | 42.10 | c–e | 27.97 | a–c | 1 |
| 41-10-03-03 | 32.78 | d–g | 28.38 | h–l | 41.83 | c–e | 28.13 | a–c | 1 |
| 43-09-02-03 | 32.72 | d–g | 28.45 | h–l | 41.60 | c–f | 28.10 | a–c | 1 |
| 43-09-03-01 | 32.39 | e–g | 27.69 | kl | 41.13 | c–f | 28.35 | ab | 1 |
| 63-04-02-02 | 32.31 | e–g | 30.80 | b–f | 41.63 | c–f | 24.50 | ef | 0 |
| 41-10-01-03 | 31.63 | f–h | 27.96 | j–l | 41.37 | c–f | 25.57 | c–f | 0 |
| 44-10-01-02 | 31.33 | gh | 29.20 | e–l | 38.93 | e–g | 25.85 | b–f | 0 |
| 70-03-07-01 | 29.78 | h | 27.87 | kl | 37.93 | fg | 23.53 | f | 0 |
| 19-05-06-03 | 29.65 | h | 27.81 | kl | 36.60 | g | 24.55 | d–f | 0 |
| p | 0.0003 | 0.0001 | 0.0001 | 0.0008 | |||||
| Mean | 33.15 | 29.58 | 42.06 | 27.79 | |||||
| LSD | 2.36 | 1.79 | 3.76 | 2.76 | |||||
| CV | 7.5% | 3.7% | 5.5% | 5.8% | |||||
| Accession | Mean | Manga 2006 | Nyankpala 2006 | N MFG 1 | |||
|---|---|---|---|---|---|---|---|
| 63-04-05-02 | 52.12 | a 2 | 51.33 | a | 52.91 | a | 2 |
| 63-04-02-02 | 51.67 | ab | 50.78 | ab | 52.55 | ab | 2 |
| 43-08-07-02 | 50.77 | bc | 49.71 | a–c | 51.83 | ab | 2 |
| 70-03-07-01 | 50.71 | bc | 49.38 | a–d | 52.03 | ab | 2 |
| 41-10-03-03 | 50.25 | c | 49.09 | a–e | 51.42 | a–c | 2 |
| 50-04-03-03 | 50.35 | c | 49.06 | a–e | 51.63 | a–c | 2 |
| 46-06-04-01 | 50.23 | c | 49.06 | a–e | 51.40 | a–d | 2 |
| 44-10-01-02 | 48.69 | d | 48.68 | a–f | 48.70 | c–g | 1 |
| 63-04-02-03 | 48.23 | d | 48.13 | a–g | 48.33 | d–h | 1 |
| 44-10-04-02 | 48.58 | d | 47.68 | a–h | 49.48 | b–f | 1 |
| 41-05-05-03 | 46.75 | fg | 46.88 | b–h | 46.61 | e–j | 0 |
| 46-06-07-02 | 48.09 | de | 46.50 | b–h | 49.69 | b–e | 0 |
| 31-04-05-02 | 46.77 | fg | 46.43 | b–h | 47.11 | e–i | 0 |
| 31-04-10-03 | 46.61 | fg | 46.35 | c–i | 46.86 | e–j | 0 |
| 43-09-03-01 | 46.90 | ef | 45.82 | c–i | 47.99 | e–i | 0 |
| 43-09-02-03 | 46.14 | f–i | 45.57 | c–i | 46.71 | e–j | 0 |
| Florunner | 45.89 | f–j | 45.07 | d–i | 46.71 | e–j | 0 |
| 41-10-01-03 | 46.41 | f–h | 45.05 | d–i | 47.77 | e–i | 0 |
| 26-10-06-03 | 46.21 | f–i | 45.02 | d–i | 47.40 | e–i | 0 |
| 45-03-06-01 | 45.65 | f–k | 44.96 | e–i | 46.33 | g–j | 0 |
| 50-04-01-03 | 45.58 | g–k | 44.82 | e–j | 46.33 | g–j | 0 |
| 60-02-02-03 | 45.12 | i–k | 44.66 | f–j | 45.58 | h–j | 0 |
| 45-04-02-01 | 45.32 | h–k | 44.64 | f–j | 46.00 | g–j | 0 |
| 63-06-08-02 | 45.54 | g–k | 44.63 | f–j | 46.44 | f–j | 0 |
| 43-08-06-02 | 45.87 | f–j | 44.63 | f–j | 47.11 | e–i | 0 |
| 55-437 | 44.85 | jk | 44.36 | f–j | 45.33 | h–j | 0 |
| 19-05-06-03 | 45.03 | i–k | 44.07 | g–j | 45.99 | g–j | 0 |
| 50-02-04-01 | 44.50 | k | 43.78 | g–j | 45.21 | ij | 0 |
| 43-09-03-02 | 44.66 | jk | 43.65 | h–j | 45.67 | g–j | 0 |
| Chinese | 42.99 | l | 41.99 | ij | 44.00 | j | 0 |
| Spanish | 42.23 | l | 40.45 | j | 44.00 | j | 0 |
| p | 0.0001 | 0.0029 | 0.0001 | ||||
| Mean | 46.07 | 46.20 | 47.91 | ||||
| LSD | 1.27 | 4.39 | 3.08 | ||||
| CV | 2.1% | 4.7% | 3.9% | ||||
| Accession | Mean | Manga 2006 | Nyankpala 2006 | Manga 2007 | Nyankpala 2007 | N MFG 1 | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 43-09-03-01 | 2027 | a 2 | 2400 | a | 336 | a–c | 3407 | a | 1966 | a–c | 4 |
| 44-10-01-02 | 1787 | ab | 2067 | ab | 342 | a–c | 2487 | a–c | 2251 | ab | 4 |
| 63-06-08-02 | 1546 | a–c | 933 | c–g | 314 | a–c | 2687 | ab | 2249 | ab | 3 |
| 43-09-02-03 | 1529 | a–d | 1733 | a–c | 294 | b–e | 2293 | a–d | 1793 | b–d | 2 |
| 43-08-06-02 | 1518 | b–d | 867 | d–g | 398 | ab | 2013 | b–i | 2795 | a | 2 |
| 43-09-03-02 | 1476 | b–d | 1170 | c–f | 270 | c–h | 2347 | a–d | 2103 | a–c | 2 |
| 46-06-07-02 | 1394 | b–e | 1600 | a–d | 261 | c–i | 2260 | a–e | 1453 | b–f | 2 |
| 19-05-06-03 | 1358 | b–f | 1533 | b–e | 286 | b–f | 2233 | a–f | 1627 | b–e | 1 |
| 41-10-01-03 | 1276 | c–g | 933 | c–g | 342 | a–c | 2033 | b–h | 1795 | b–d | 1 |
| 26-10-06-03 | 1262 | c–g | 833 | d–g | 227 | c–i | 1833 | b–i | 2153 | a–c | 1 |
| 45-04-02-01 | 1245 | c–g | 933 | c–g | 284 | b–g | 2173 | a–g | 1590 | b–e | 1 |
| 50-02-04-01 | 1224 | c–h | 1067 | c–g | 305 | a–d | 1553 | b–i | 1969 | a–c | 2 |
| 63-04-05-02 | 1219 | c–h | 933 | c–g | 171 | e–j | 1700 | b–i | 2073 | a–c | 1 |
| 44-10-04-02 | 1196 | c–i | 933 | c–g | 271 | c–h | 2280 | a–e | 1301 | c–f | 0 |
| Florunner | 1134 | c–i | 500 | fg | 228 | c–i | 2147 | b–g | 1661 | b-e | 0 |
| 41-10-03-03 | 1125 | c–i | 667 | fg | 259 | c–i | 1740 | b–i | 1835 | b–d | 0 |
| 43-08-07-02 | 1097 | c–i | 1267 | b–f | 285 | b–f | 1420 | c–i | 1417 | b–f | 0 |
| Spanish | 1026 | d–j | 1000 | c–g | 142 | ij | 1540 | b–i | 1421 | b–f | 0 |
| 31-04-05-02 | 954 | e–k | 933 | c–g | 252 | c–i | 1373 | c–i | 1257 | c–f | 0 |
| 45-03-06-01 | 939 | e–k | 867 | d–g | 138 | ij | 1760 | b–i | 991 | d–f | 0 |
| 50-04-03-03 | 913 | e–k | 733 | e–g | 217 | c–i | 1753 | b–i | 949 | d–f | 0 |
| 41-05-05-03 | 878 | f–k | 833 | d–g | 312 | a–c | 1113 | d–i | 1255 | c–f | 1 |
| 60-02-02-03 | 841 | g–k | 733 | e–g | 160 | f–j | 1000 | f–i | 1470 | b–f | 0 |
| 50-04-01-03 | 831 | g–k | 1333 | b–f | 168 | f–j | 1200 | d–i | 622 | f | 0 |
| 70-03-07-01 | 779 | g–k | 733 | e–g | 181 | d–j | 1213 | d–i | 989 | d–f | 0 |
| 46-06-04-01 | 721 | h–k | 667 | fg | 240 | c–i | 967 | g–i | 1009 | d–f | 0 |
| 63-04-02-03 | 712 | i–k | 567 | fg | 155 | h–j | 1047 | e–i | 861 | ef | 0 |
| 31-04-10-03 | 565 | jk | 667 | fg | 229 | c–i | 793 | i | 570 | f | 0 |
| 63-04-02-02 | 501 | k | 267 | g | 158 | g–j | 887 | hi | 625 | f | 0 |
| Chinese | n.d. | 733 | e–g | 425 | a | n.d. | n.d. | 0 | |||
| 55-437 | n.d. | n.d. | 55 | j | 1600 | b–i | 799 | ef | 0 | ||
| p | <0.0001 | 0.0003 | <0.0001 | 0.0161 | 0.0002 | ||||||
| Mean | 1140 | 1006 | 249 | 1762 | 1495 | ||||||
| LSD | 506 | 895 | 127 | 1239 | 916 | ||||||
| CV | 54.0% | 48.1% | 31.2% | 43.0% | 35.4% | ||||||
| 2006 Manga | AUDPCELS | AUDPCLLS | T50ELS | T50LLS | PODYIELD | |||||
| AUDPCELS | 1.0000 | |||||||||
| AUDPCLLS | 0.8423 | *** 2 | 1.0000 | |||||||
| T50ELS | n.d. | n.d. | n.d. | |||||||
| T50LLS | −0.1654 | −0.3492 | ** | n.d. | 1.0000 | |||||
| PODYIELD | 0.2675 | * | 0.2768 | ** | n.d. | 0.0913 | 1.0000 | |||
| 2006 Nyankpala | AUDPCELS | AUDPCLLS | T50ELS | T50LLS | PODYIELD | |||||
| AUDPCELS | 1.0000 | |||||||||
| AUDPCLLS | 0.7735 | *** | 1.0000 | |||||||
| T50ELS | 0.2565 | * | 0.2740 | ** | 1.0000 | |||||
| T50LLS | −0.1009 | −0.3126 | ** | 0.1037 | 1.0000 | |||||
| PODYIELD | −0.1271 | −0.0050 | −0.2816 | ** | −0.0379 | 1.0000 | ||||
| 2007 Manga | AUDPCELS | AUDPCLLS | T50ELS | T50LLS | PODYIELD | |||||
| AUDPCELS | 1.0000 | |||||||||
| AUDPCLLS | 0.6608 | *** | 1.0000 | |||||||
| T50ELS | −0.0160 | 0.0409 | 1.0000 | |||||||
| T50LLS | n.d. | n.d. | n.d. | n.d. | ||||||
| PODYIELD | −0.0613 | −0.1501 | 0.1178 | n.d. | 1.0000 | |||||
| 2007 Nyankpala | AUDPCELS | AUDPCLLS | T50ELS | T50LLS | PODYIELD | |||||
| AUDPCELS | 1.0000 | |||||||||
| AUDPCLLS | 0.8104 | *** | 1.0000 | |||||||
| T50ELS | 0.0483 | −0.0492 | 1.0000 | |||||||
| T50LLS | n.d. | n.d. | n.d. | n.d. | ||||||
| PODYIELD | −0.0634 | −0.1161 | 0.0551 | n.d. | 1.0000 | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Denwar, N.N.; Simpson, C.E.; Starr, J.L.; Wheeler, T.A.; Burow, M.D. Evaluation and Selection of Interspecific Lines of Groundnut (Arachis hypogaea L.) for Resistance to Leaf Spot Disease and for Yield Improvement. Plants 2021, 10, 873. https://doi.org/10.3390/plants10050873
Denwar NN, Simpson CE, Starr JL, Wheeler TA, Burow MD. Evaluation and Selection of Interspecific Lines of Groundnut (Arachis hypogaea L.) for Resistance to Leaf Spot Disease and for Yield Improvement. Plants. 2021; 10(5):873. https://doi.org/10.3390/plants10050873
Chicago/Turabian StyleDenwar, Nicholas N., Charles E. Simpson, James L. Starr, Terry A. Wheeler, and Mark D. Burow. 2021. "Evaluation and Selection of Interspecific Lines of Groundnut (Arachis hypogaea L.) for Resistance to Leaf Spot Disease and for Yield Improvement" Plants 10, no. 5: 873. https://doi.org/10.3390/plants10050873