
Potential Role of Rhizobacteria Isolated from Citrus Rhizosphere for Biological Control of Citrus Dry Root Rot


 , ,
, , 
Abstract
:1. Introduction
2. Results
2.1. Antagonistic Activity
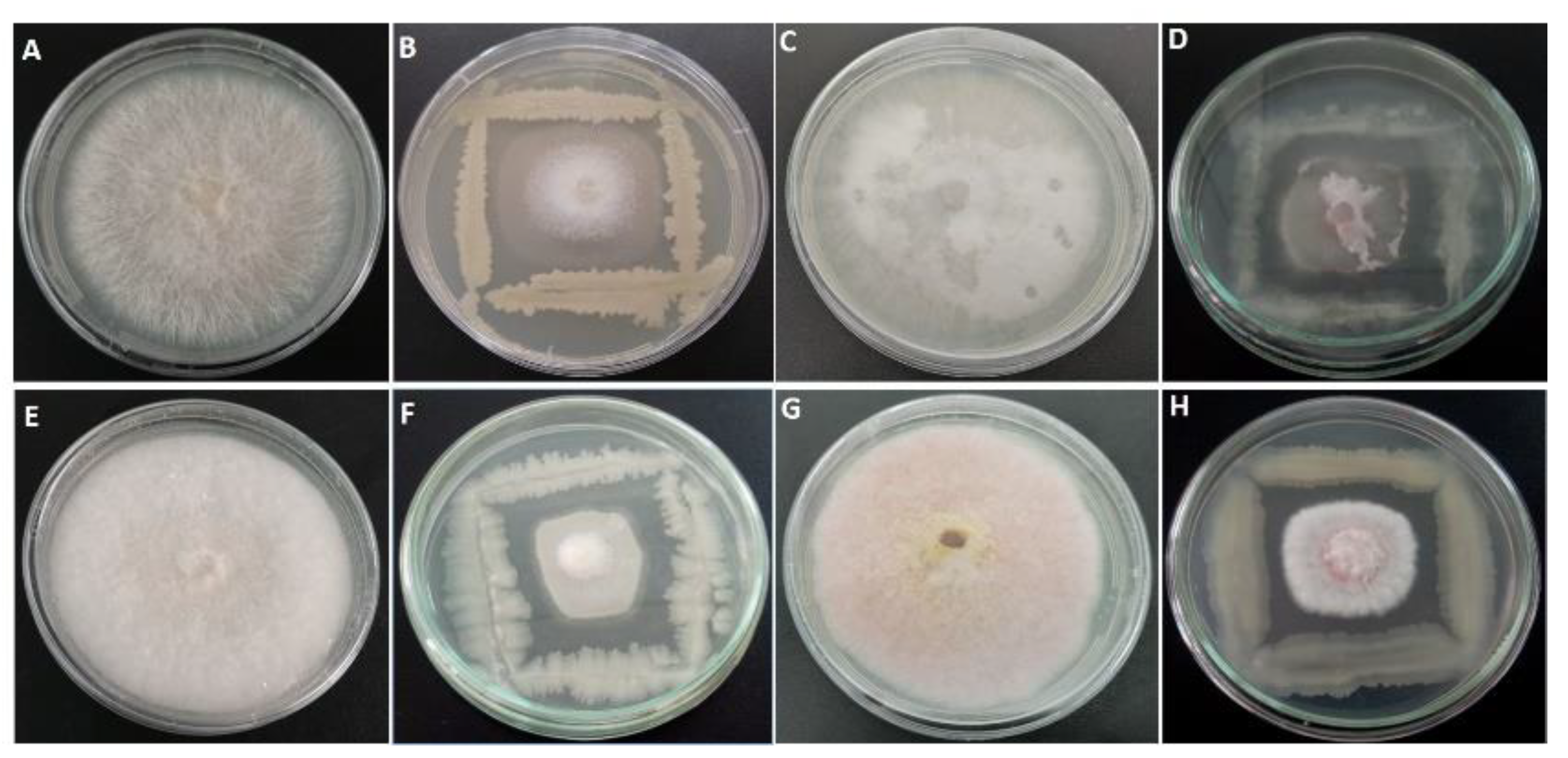
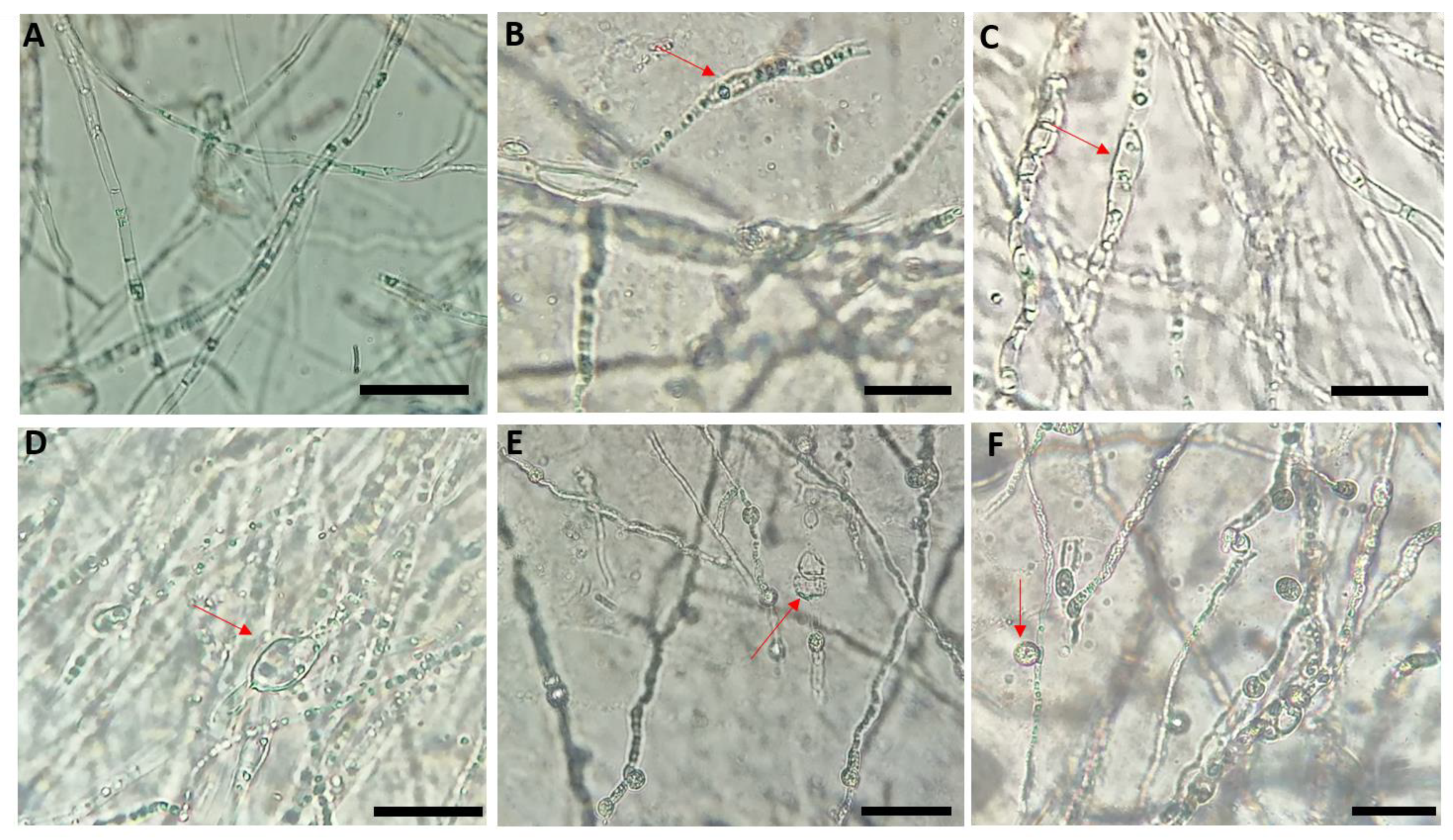
2.2. Effect of Bacterial Isolates on the Mycelial Structure of Neocosmospora Solani
2.3. Identification of Bacterial Isolates by 16S rDNA Amplicon Sequencing
2.4. Evaluation of Biocontrol Activities and Plant Growth Stimulating Traits Exercised by Selected Bacteria
2.4.1. Indirect Antagonism Activity of Selected Bacteria
2.4.2. Biochemical Traits
2.5. Hypersensitivity Test
2.6. Effect of Rhizobacteria on Plant Growth of Brassica Napus
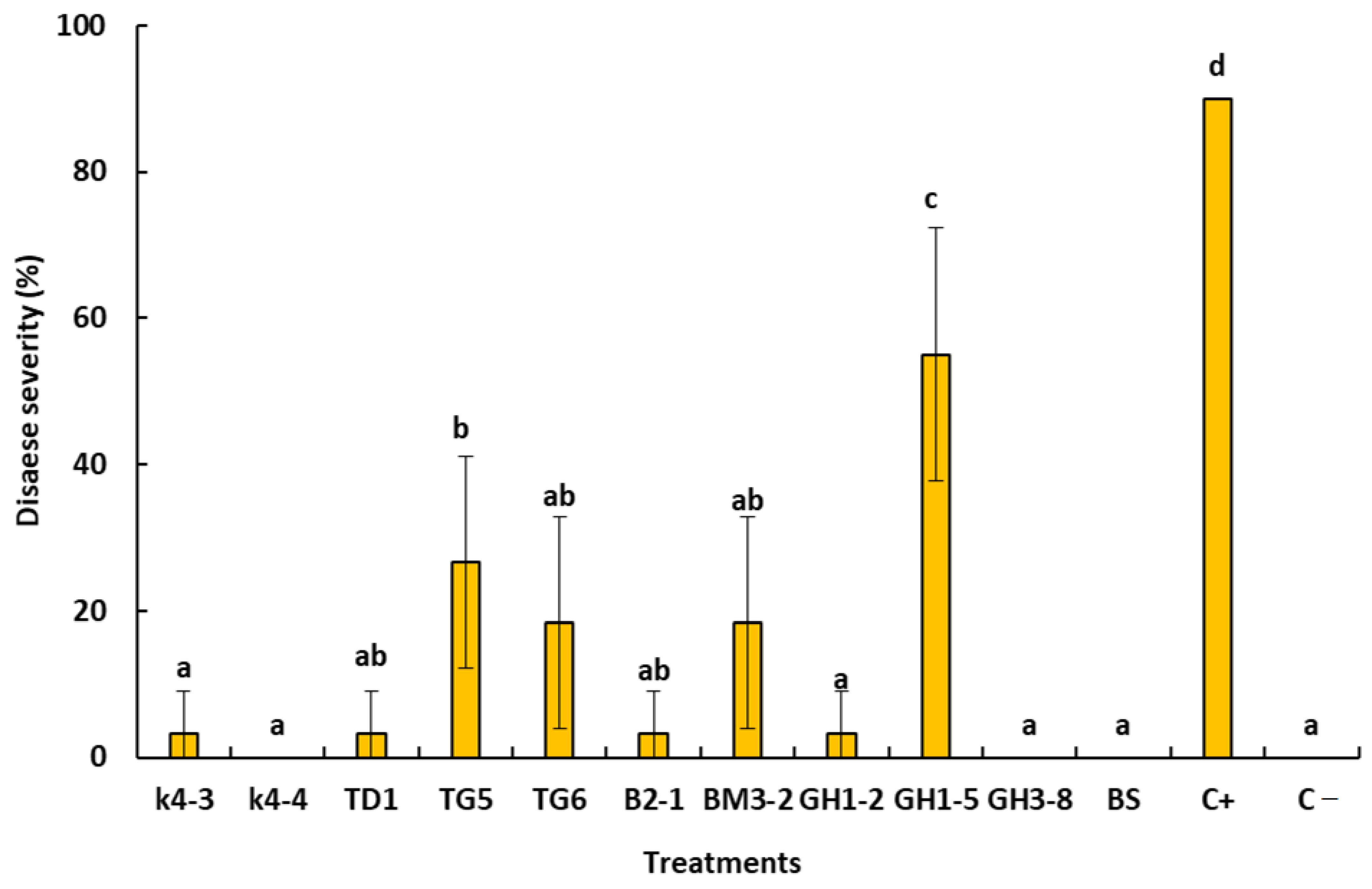
2.7. In Planta Bioassays
3. Discussion
4. Materials and Methods
4.1. Origin of Fungal Isolates
4.2. Isolation of Bacteria from Citrus Rhizosphere
4.3. Screening of Antagonist Bacteria
4.4. Effect of Bacterial Isolates on the Mycelial Structure of Neocosmospora Solani
4.5. Bacterial Identification
4.6. Evaluation of Biocontrol Activities and Plant Growth Stimulating Attributes
4.6.1. Indirect Antagonist Activity of Selected Bacteria
4.6.2. Microbial Traits
4.7. Greenhouse Experiment
4.7.1. Hypersensitivity Test
4.7.2. PGPR Test on Brassica Napus
4.7.3. In Planta Antagonism
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ezrari, S.; Lahlali, R.; Radouane, N.; Tahiri, A.; Asfers, A.; Boughalleb‑M’Hamdi, N.; Amiri, S.; Lazraq, A. Characterization of Fusarium species causing dry root rot disease of citrus trees in Morocco. J. Plant Dis. Prot. 2020. [Google Scholar] [CrossRef]
- Jaouad, M.; Moinina, A.; Ezrari, S.; Lahlali, R. Key pests and diseases of citrus trees with emphasis on root rot diseases: An overview. Mor. J. Agri. Sci. 2020, 1, 149–160. [Google Scholar]
- Sandoval-Denis, M.P.; Guarnaccia, V.; Polizzi, G.; Crous, P.W. Symptomatic citrus trees reveal a new pathogenic lineage in Fusarium and two new Neocosmospora species. Persoonia Mol. Phylogeny Evol. Fungi 2018, 40, 1–25. [Google Scholar] [CrossRef] [Green Version]
- Kurt, Ş.; Uysal, A.; Soylu, E.M.; Kara, M.; Soylu, S. Characterization and pathogenicity of Fusarium solani associated with dry root rot of citrus in the eastern Mediterranean region of Turkey. J. Gen. Plant Pathol. 2020, 20, 127. [Google Scholar] [CrossRef]
- Liu, H.F.; Zhou, J.; Liao, J.; Yi, J.P.; Ma, D.F.; Deng, J.X. Grafted twig rot on Citrus sinensis caused by a member of the Fusarium solani species complex. Can. J. Plant Pathol. 2019, 42, 133–139. [Google Scholar] [CrossRef]
- Spina, S.; Coco, V.; Gentile, A.; Catara, A.; Cirvilleri, G. Association of Fusarium solani with rolABC and wild type troyer citrange. J. Plant Pathol. 2008, 90, 479–486. [Google Scholar] [CrossRef]
- Lombard, L.; van der Merwe, N.A.; Groenewald, J.Z.; Crous, P.W. Generic concepts in Nectriaceae. Stud. Mycol. 2015, 80, 189–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adesemoye, A.; Eskalen, A.; Faber, B.; Bender, G.; Connell, N.O.; Kallsen, C.; Shea, T. Current knowledge on Fusarium dry rot of citrus. Citrograph 2011, 2(7), 29–33. [Google Scholar]
- Labuschagne, N.; Van Der Vegte, F.A.; Kotzé, J.M. Interaction between Fusarium solani and Tylenchulus semipenetrans on citrus roots. Phytophylactica 1989, 21, 29–33. [Google Scholar]
- Yaseen, T.; D’Onghia, A.M. Fusarium spp. associated to citrus dry root rot: An emerging issue for Mediterranean citriculture. Acta Hortic. 2012, 940, 647–656. [Google Scholar] [CrossRef]
- Guarnaccia, V.; Van Niekerk, J.; Crous, P.W.; Sandoval-Denis, M. Neocosmospora spp. associated with dry root rot of citrus in South Africa. Phytopathol. Mediterr. 2021, 60, 81–102. [Google Scholar]
- Hammam, M.M.A.; El-Mohamedy, R.S.R.; Abd-El-Kareem, F.; Abd-Elgawad, M.M.M. Evaluation of soil amended with bio-agents and compost alone or in combination for controlling citrus nematode Tylenchulus semipenetrans and Fusarium dry root rot on Volkamer lime under greenhouse conditions. Int. J. ChemTech Res. 2016, 9, 86–96. [Google Scholar]
- El-Mohamedy, R.S.R. Efficiency of different application methods of biocontrol agents and biocides in control of Fusarium root rot on some citrus rootstocks. Arch. Phytopathol. Plant Prot. 2009, 42, 819–828. [Google Scholar] [CrossRef]
- Parikh, L.; Adesemoye, A.O. Impact of delivery method on the efficacy of biological control agents and the virulence of Fusarium root rot pathogens in the greenhouse. Biocontrol. Sci. Technol. 2018, 28, 1191–1202. [Google Scholar] [CrossRef]
- Veldman, W.M.; Regnier, T.; Augustyn, W.A. Biocontrol of Fusarium mangiferae responsible for mango malformation using bacterial isolates. Sci. Hortic. 2018, 230, 186–195. [Google Scholar] [CrossRef]
- Vejan, P.; Abdullah, R.; Khadiran, T.; Ismail, S.; Nasrulhaq Boyce, A. Role of plant growth promoting rhizobacteria in agricultural sustainability-A review. Molecules 2016, 21, 573. [Google Scholar] [CrossRef]
- Dey, R.; Pal, K.K.; Tilak, K. Plant growth promoting rhizobacteria in crop protection and challenges. In Future Challenges in Crop Protection against Fungal Pathogens; Springer: New York, NY, USA, 2014; pp. 31–58. [Google Scholar]
- Kloepper, J.W.; Zablotowicz, R.M.; Tipping, E.M.; Lifshitz, R. Plant growth promotion mediated by bacterial rhizosphere colonizers. In The Rhizosphere and Plant Growth; Keister, D.L., Cregan, P.B., Eds.; Springer: Dordrecht, The Netherlands, 1991; pp. 315–326. [Google Scholar]
- Glick, B.R. Bacteria with ACC deaminase can promote plant growthand help to feed the world. Microbiol. Res. 2014, 169, 30–39. [Google Scholar] [CrossRef]
- Swarnalakshmi, K.; Yadav, V.; Tyagi, D.; Dhar, D.W.; Kannepalli, A.; Kumar, S. Significance of Plant Growth Promoting Rhizobacteria in Grain Legumes: Growth Promotion and Crop Production. Plants 2020, 9, 1596. [Google Scholar] [CrossRef]
- Prasad, M.; Srinivasan, R.; Chaudhary, M.; Choudhary, M.; Jat, L.K. Plant growth promoting rhizobacteria (PGPR) for sustainable agriculture: Perspectives and challenges. In PGPR Amelioration in Sustainable Agriculture; Elsevier: Cambridge, UK, 2019; pp. 129–157. [Google Scholar]
- Slama, H.B.; Cherif-Silini, H.; Bouket, A.C.; Qader, M.; Silini, A.; Yahiaoui, B.; Alenezi, F.N.; Luptakova, L.; Triki, M.A.; Vallat, A.; et al. Screening for Fusarium antagonistic bacteria from contrasting niches designated the endophyte bacillus halotoleransas plant warden against fusarium. Front. Microbiol. 2019, 10, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Suárez-Moreno, Z.R.; Vinchira-Villarraga, D.M.; Vergara-Morales, D.I.; Castellanos, L.; Ramos, F.A.; Guarnaccia, C.; Degrassi, G.; Venturi, V.; Moreno-Sarmiento, N. Plant-growth promotion and biocontrol properties of three Streptomyces spp. isolates to control bacterial rice pathogens. Front. Microbiol. 2019, 10, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Díaz Herrera, S.; Grossi, C.; Zawoznik, M.; Groppa, M.D. Wheat seeds harbour bacterial endophytes with potential as plant growth promoters and biocontrol agents of Fusarium graminearum. Microbiol. Res. 2016, 186–187, 37–43. [Google Scholar] [CrossRef]
- Paramanandham, P.; Rajkumari, J.; Pattnaik, S.; Busi, S. Biocontrol Potential against Fusarium oxysporum f. sp. lycopersici and Alternaria solani and Tomato Plant Growth Due to Plant Growth–Promoting Rhizobacteria. Int. J. Veg. Sci. 2017, 23, 294–303. [Google Scholar] [CrossRef]
- Singh, M.; Singh, D.; Gupta, A.; Pandey, K.D.; Singh, P.K.; Kumar, A. Plant growth promoting rhizobacteria: Application in biofertilizers and biocontrol of phytopathogens. In PGPR Amelioration in Sustainable Agriculture; Elsevier: Amsterdam, The Netherlands, 2019; pp. 41–66. [Google Scholar]
- Verma, M.; Mishra, J.; Arora, N.K. Plant Growth-Promoting Rhizobacteria: Diversity and Applications. Environ. Biotechnol. Sustain. Future 2019, 129–173. [Google Scholar] [CrossRef]
- Basu, A.; Prasad, P.; Das, S.N.; Kalam, S.; Sayyed, R.Z.; Reddy, M.S.; El Enshasy, H. Plant Growth Promoting Rhizobacteria (PGPR) as Green Bioinoculants: Recent Developments, Constraints, and Prospects. Sustainability 2021, 13, 1140. [Google Scholar] [CrossRef]
- Bubici, G.; Kaushal, M.; Prigigallo, M.I.; Cabanás, C.G.L.; Mercado-Blanco, J. Biological control agents against Fusarium wilt of banana. Front. Microbiol. 2019, 10, 616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lahlali, R.; Mchachti, O.; Radouane, N.; Ezrari, S.; Belabess, Z.; Khayi, S.; Mentag, R.; Tahiri, A.; Barka, E.A. The Potential of Novel Bacterial Isolates from Natural Soil for the Control of Brown Rot Disease (Monilinia fructigena) on Apple Fruits. Agronomy 2020, 10, 1814. [Google Scholar] [CrossRef]
- Knight, C.A.; Bowman, M.J.; Frederick, L.; Day, A.; Lee, C.; Dunlap, C.A. The first report of antifungal lipopeptide production by a Bacillus subtilis subsp. inaquosorum strain. Microbiol. Res. 2018, 216, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Tapi, A.; Chollet-Imbert, M.; Scherens, B.; Jacques, P. New approach for the detection of non-ribosomal peptide synthetase genes in Bacillus strains by polymerase chain reaction. Appl. Microbiol. Biotechnol. 2010, 85, 1521–1531. [Google Scholar] [CrossRef] [PubMed]
- Toral, L.; Rodríguez, M.; Béjar, V.; Sampedro, I. Antifungal activity of lipopeptides from Bacillus XT1 CECT 8661 against Botrytis cinerea. Front. Microbiol. 2018, 9, 1–12. [Google Scholar] [CrossRef]
- Stanković, S.; Mihajlović, S.; Draganić, V.; Dimkić, I.; Vukotić, G.; Berić, T.; Fira, D. Screening for the presence of biosynthetic genes for antimicrobial lipopeptides in natural isolates of Bacillus sp. Arch. Biol. Sci. 2012, 64, 1425–1432. [Google Scholar] [CrossRef]
- Compaoré, C.S.; Nielsen, D.S.; Sawadogo-Lingani, H.; Berner, T.S.; Nielsen, K.F.; Adimpong, D.B.; Diawara, B.; Ouédraogo, G.A.; Jakobsen, M.; Thorsen, L. Bacillus amyloliquefaciens ssp. plantarum strains as potential protective starter cultures for the production of Bikalga, an alkaline fermented food. J. Appl. Microbiol. 2013, 115, 133–146. [Google Scholar] [CrossRef]
- Sutyak, K.E.; Wirawan, R.E.; Aroutcheva, A.A.; Chikindas, M.L. Isolation of the Bacillus subtilis antimicrobial peptide subtilosin from the dairy product-derived Bacillus amyloliquefaciens. J. Appl. Microbiol. 2008, 104, 1067–1074. [Google Scholar] [CrossRef] [Green Version]
- Ryu, C.-M. Promoting plant protection by root-associated microbes. Plant Pathol. J. 2013, 29, 123. [Google Scholar]
- Li, Z.; Guo, B.; Wan, K.; Cong, M.; Huang, H.; Ge, Y. Effects of bacteria-free filtrate from Bacillus megaterium strain L2 on the mycelium growth and spore germination of Alternaria alternata. Biotechnol. Biotechnol. Equip. 2015, 29, 1062–1068. [Google Scholar] [CrossRef] [Green Version]
- Gong, A.D.; Li, H.P.; Yuan, Q.S.; Song, X.S.; Yao, W.; He, W.J.; Zhang, J.B.; Liao, Y.C. Antagonistic mechanism of iturin a and plipastatin a from Bacillus amyloliquefaciens S76-3 from wheat spikes against Fusarium graminearum. PLoS ONE 2015, 10, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Devi, A.R.; Sharma, G.D.; Majumdar, P.B.; Pandey, P. A multispecies consortium of bacteria having plant growth promotion and antifungal activities, for the management of Fusarium wilt complex disease in potato (Solanum tuberosum L.). Biocatal. Agric. Biotechnol. 2018, 16, 614–624. [Google Scholar] [CrossRef]
- Palmieri, D.; Vitullo, D.; De Curtis, F.; Lima, G. A microbial consortium in the rhizosphere as a new biocontrol approach against Fusarium decline of chickpea. Plant Soil 2017, 412, 425–439. [Google Scholar] [CrossRef]
- Przemieniecki, S.W.; Kurowski, T.P.; Kotlarz, K.; Krawczyk, K.; Damszel, M.; Pszczółkowska, A.; Kacprzak-Siuda, K.; Chareńska, A.; Mastalerz, J. Bacteria Isolated from Treated Wastewater for Biofertilization and Crop Protection Against Fusarium spp. Pathogens. J. Soil Sci. Plant Nutr. 2019, 19, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Khabbaz, S.E.; Wang, A.; Li, H.; Abbasi, P.A. Detection and characterization of broad-spectrum antipathogen activity of novel rhizobacterial isolates and suppression of Fusarium crown and root rot disease of tomato. J. Appl. Microbiol. 2015, 118, 685–703. [Google Scholar] [CrossRef]
- Chenniappan, C.; Narayanasamy, M.; Daniel, G.M.; Ramaraj, G.B.; Ponnusamy, P.; Sekar, J.; Vaiyapuri Ramalingam, P. Biocontrol efficiency of native plant growth promoting rhizobacteria against rhizome rot disease of turmeric. Biol. Control 2019, 129, 55–64. [Google Scholar] [CrossRef]
- Khan, N.; Martínez-Hidalgo, P.; Ice, T.A.; Maymon, M.; Humm, E.A.; Nejat, N.; Sanders, E.R.; Kaplan, D.; Hirsch, A.M. Antifungal activity of bacillus species against Fusarium and analysis of the potential mechanisms used in biocontrol. Front. Microbiol. 2018, 9, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Einloft, T.C.; Hartke, S.; de Oliveira, P.B.; Saraiva, P.S.; Dionello, R.G. Selection of rhizobacteria for biocontrol of Fusarium verticillioides on non-rhizospheric soil and maize seedlings roots. Eur. J. Plant Pathol. 2021, 1–16. [Google Scholar] [CrossRef]
- Ali, M.A.; Ren, H.; Ahmed, T.; Luo, J.; An, Q.; Qi, X.; Li, B. Antifungal Effects of Rhizospheric Bacillus Species Against Bayberry Twig Blight Pathogen Pestalotiopsis versicolor. Agronomy 2020, 10, 1811. [Google Scholar] [CrossRef]
- Amna; Xia, Y.; Farooq, M.A.; Javed, M.T.; Kamran, M.A.; Mukhtar, T.; Ali, J.; Tabassum, T.; Rehman, S.; Hussain Munis, M.F.; et al. Multi-stress tolerant PGPR Bacillus xiamenensis PM14 activating sugarcane (Saccharum officinarum L.) red rot disease resistance. Plant Physiol. Biochem. 2020, 151, 640–649. [Google Scholar] [CrossRef] [PubMed]
- Ji, Z.L.; Peng, S.; Chen, L.L.; Liu, Y.; Yan, C.; Zhu, F. Identification and characterization of a serine protease from Bacillus licheniformis W10: A potential antifungal agent. Int. J. Biol. Macromol. 2020, 145, 594–603. [Google Scholar] [CrossRef]
- Jeong, M.H.; Lee, Y.S.; Cho, J.Y.; Ahn, Y.S.; Moon, J.H.; Hyun, H.N.; Cha, G.S.; Kim, K.Y. Isolation and characterization of metabolites from Bacillus licheniformis MH48 with antifungal activity against plant pathogens. Microb. Pathog. 2017, 110, 645–653. [Google Scholar] [CrossRef] [PubMed]
- Won, S.J.; Choub, V.; Kwon, J.H.; Kim, D.H.; Ahn, Y.S. The control of fusarium root rot and development of coastal pine (Pinus thunbergii Parl.) Seedlings in a container nursery by use of Bacillus licheniformis MH48. Forests 2018, 10, 6. [Google Scholar] [CrossRef] [Green Version]
- Liu, K.; Newman, M.; McInroy, J.A.; Hu, C.H.; Kloepper, J.W. Selection and assessment of plant growth-promoting rhizobacteria for biological control of multiple plant diseases. Phytopathology 2017, 107, 928–936. [Google Scholar] [CrossRef] [Green Version]
- Damasceno, C.L.; Duarte, E.A.A.; dos Santos, L.B.P.R.; de Oliveira, T.A.S.; de Jesus, F.N.; de Oliveira, L.M.; Góes-Neto, A.; Soares, A.C.F. Postharvest biocontrol of anthracnose in bananas by endophytic and soil rhizosphere bacteria associated with sisal (Agave sisalana) in Brazil. Biol. Control 2019, 137, 104016. [Google Scholar] [CrossRef]
- Rojas-Solís, D.; Zetter-Salmón, E.; Contreras-Pérez, M.; Rocha-Granados, M.D.C.; Macías-Rodríguez, L.; Santoyo, G. Pseudomonas stutzeri E25 and Stenotrophomonas maltophilia CR71 endophytes produce antifungal volatile organic compounds and exhibit additive plant growth-promoting effects. Biocatal. Agric. Biotechnol. 2018, 13, 46–52. [Google Scholar] [CrossRef]
- Alijani, Z.; Amini, J.; Ashengroph, M.; Bahramnejad, B. Volatile compounds mediated effects of Stenotrophomonas maltophilia strain UN1512 in plant growth promotion and its potential for the biocontrol of Colletotrichum nymphaeae. Physiol. Mol. Plant Pathol. 2020, 112, 101555. [Google Scholar] [CrossRef]
- Sahu, K.P.; Kumar, A.; Patel, A.; Kumar, M.; Gopalakrishnan, S.; Prakash, G.; Rathour, R.; Gogoi, R. Rice Blast Lesions: An Unexplored Phyllosphere Microhabitat for Novel Antagonistic Bacterial Species Against Magnaporthe oryzae. Microb. Ecol. 2020. [Google Scholar] [CrossRef] [PubMed]
- An, X.; Tian, C.; Xu, J.; Dong, F.; Liu, X.; Wu, X.; Zheng, Y. Characterization of hexaconazole-degrading strain Sphingobacterium multivorum and analysis of transcriptome for biodegradation mechanism. Sci. Total Environ. 2020, 722, 137171. [Google Scholar] [CrossRef] [PubMed]
- Murolo, S.; Concas, J.; Romanazzi, G. Use of biocontrol agents as potential tools in the management of chestnut blight. Biol. Control 2019, 132, 102–109. [Google Scholar] [CrossRef]
- Guevara-Avendaño, E.; Bejarano-Bolívar, A.A.; Kiel-Martínez, A.L.; Ramírez-Vázquez, M.; Méndez-Bravo, A.; von Wobeser, E.A.; Sánchez-Rangel, D.; Guerrero-Analco, J.A.; Eskalen, A.; Reverchon, F. Avocado rhizobacteria emit volatile organic compounds with antifungal activity against Fusarium solani, Fusarium sp. associated with Kuroshio shot hole borer, and Colletotrichum gloeosporioides. Microbiol. Res. 2019, 219, 74–83. [Google Scholar] [CrossRef]
- Yuttavanichakul, W.; Lawongsa, P.; Wongkaew, S.; Teaumroong, N.; Boonkerd, N.; Nomura, N.; Tittabutr, P. Improvement of peanut rhizobial inoculant by incorporation of plant growth promoting rhizobacteria (PGPR) as biocontrol against the seed borne fungus, Aspergillus niger. Biol. Control 2012, 63, 87–97. [Google Scholar] [CrossRef]
- Khunnamwong, P.; Lertwattanasakul, N.; Jindamorakot, S.; Suwannarach, N.; Matsui, K.; Limtong, S. Evaluation of antagonistic activity and mechanisms of endophytic yeasts against pathogenic fungi causing economic crop diseases. Folia Microbiol. 2020, 65, 573–590. [Google Scholar] [CrossRef]
- Schulz-Bohm, K.; Martín-Sánchez, L.; Garbeva, P. Microbial volatiles: Small molecules with an important role in intra- and inter-kingdom interactions. Front. Microbiol. 2017, 8, 1–10. [Google Scholar] [CrossRef]
- Ramette, A.; Frapolli, M.; Défago, G.; Moënne-Loccoz, Y. Phylogeny of HCN Synthase-Encoding hcnBC Genes in Biocontrol Fluorescent Pseudomonads and Its Relationship with Host Plant Species and HCN Synthesis Ability. Mol. Plant-Microbe Interact. 2003, 16, 525–535. [Google Scholar] [CrossRef] [Green Version]
- Blumer, C.; Haas, D. Mechanism, regulation, and ecological role of bacterial cyanide biosynthesis. Arch. Microbiol. 2000, 173, 170–177. [Google Scholar] [CrossRef]
- Aeron, A.; Dubey, R.C.; Maheshwari, D.K.; Pandey, P.; Bajpai, V.K.; Kang, S.C. Multifarious activity of bioformulated Pseudomonas fluorescens PS1 and biocontrol of Sclerotinia sclerotiorum in Indian rapeseed (Brassica campestris L.). Eur. J. Plant Pathol. 2011, 131, 81–93. [Google Scholar] [CrossRef]
- Kumar, P.; Thakur, S.; Dhingra, G.K.; Singh, A.; Pal, M.K.; Harshvardhan, K.; Dubey, R.C.; Maheshwari, D.K. Inoculation of siderophore producing rhizobacteria and their consortium for growth enhancement of wheat plant. Biocatal. Agric. Biotechnol. 2018, 15, 264–269. [Google Scholar] [CrossRef]
- Souza, D.R.; Ambrosini, A.; Passaglia, L.M.P. Plant growth-promoting bacteria as inoculants in agricultural soils. Genet. Mol. Biol. 2015, 38, 401–419. [Google Scholar] [CrossRef]
- Lebrazi, S.; Niehaus, K.; Bednarz, H.; Fadil, M.; Chraibi, M.; Fikri-Benbrahim, K. Screening and optimization of indole-3-acetic acid production and phosphate solubilization by rhizobacterial strains isolated from Acacia cyanophylla root nodules and their effects on its plant growth. J. Genet. Eng. Biotechnol. 2020, 18. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.; Yao, T.; Feng, C.; Chen, L.; Li, J.; Wang, L. Identification and biocontrol potential of antagonistic bacteria strains against Sclerotinia sclerotiorum and their growth-promoting effects on Brassica napus. Biol. Control 2017, 104, 35–43. [Google Scholar] [CrossRef]
- Majeed, A.; Kaleem Abbasi, M.; Hameed, S.; Imran, A.; Rahim, N. Isolation and characterization of plant growth-promoting rhizobacteria from wheat rhizosphere and their effect on plant growth promotion. Front. Microbiol. 2015, 6, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, P.; Dubey, R.C.; Maheshwari, D.K. Bacillus strains isolated from rhizosphere showed plant growth promoting and antagonistic activity against phytopathogens. Microbiol. Res. 2012, 167, 493–499. [Google Scholar] [CrossRef] [PubMed]
- Prajakta, B.M.; Suvarna, P.P.; Raghvendra, S.P.; Alok, R.R. Potential biocontrol and superlative plant growth promoting activity of indigenous Bacillus mojavensis PB‑35 (R11) of soybean (Glycine max) rhizosphere. SN Appl. Sci. 2019, 1, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Lim, S.M.; Yoon, M.Y.; Choi, G.J.; Choi, Y.H.; Jang, K.S.; Shin, T.S.; Park, H.W.; Yu, N.H.; Kim, Y.H.; Kim, J.C. Diffusible and volatile antifungal compounds produced by an antagonistic Bacillus velezensis G341 against various phytopathogenic fungi. Plant Pathol. J. 2017, 33, 488–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-Raudales, I.; De La Cruz-Rodríguez, Y.; Vega-Arreguín, J.; Alvarado-Gutiérrez, A.; Fraire-Mayorga, A.; Alvarado-Rodríguez, M.; Balderas-Hernández, V.; Gómez-Soto, J.M.; Fraire-Velázquez, S. Draft Genome Sequence of Bacillus velezensis 3A-25B, a Strain with Biocontrol Activity against Fungal and Oomycete Root Plant Phytopathogens, Isolated from Grassland Soil. Genome Announc. 2017, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fira, D.; Dimkić, I.; Berić, T.; Lozo, J.; Stanković, S. Biological control of plant pathogens by Bacillus species. J. Biotechnol. 2018, 285, 44–55. [Google Scholar] [CrossRef]
- Dimkic, I.; Stankovic, S.; Nišavic, M.; Petkovic, M.; Ristivojevic, P.; Fira, D.; Beric, T. The profile and antimicrobial activity of Bacillus lipopeptide extracts of five potential biocontrol strains. Front. Microbiol. 2017, 8, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Velho, R.V.; Medina, L.F.C.; Segalin, J.; Brandelli, A. Production of lipopeptides among Bacillus strains showing growth inhibition of phytopathogenic fungi. Folia Microbiol. 2011, 56, 297–303. [Google Scholar] [CrossRef]
- Cao, Y.; Pi, H.; Chandrangsu, P.; Li, Y.; Wang, Y.; Zhou, H.; Xiong, H.; Helmann, J.D.; Cai, Y. Antagonism of Two Plant-Growth Promoting Bacillus velezensis Isolates Against Ralstonia solanacearum and Fusarium oxysporum. Sci. Rep. 2018, 8, 1–14. [Google Scholar] [CrossRef]
- Adeniji, A.A.; Aremu, O.S.; Babalola, O.O. Selecting lipopeptide-producing, Fusarium-suppressing Bacillus spp.: Metabolomic and genomic probing of Bacillus velezensis NWUMFkBS10.5. Microbiologyopen 2019, 8, e00742. [Google Scholar] [CrossRef] [Green Version]
- Palazzini, J.M.; Dunlap, C.A.; Bowman, M.J.; Chulze, S.N. Bacillus velezensis RC 218 as a biocontrol agent to reduce Fusarium head blight and deoxynivalenol accumulation: Genome sequencing and secondary metabolite cluster profiles. Microbiol. Res. 2016, 192, 30–36. [Google Scholar] [CrossRef]
- Zalila-Kolsi, I.; Ben Mahmoud, A.; Ali, H.; Sellami, S.; Nasfi, Z.; Tounsi, S.; Jamoussi, K. Antagonist effects of Bacillus spp. strains against Fusarium graminearum for protection of durum wheat (Triticum turgidum L. subsp. durum). Microbiol. Res. 2016, 192, 148–158. [Google Scholar] [CrossRef]
- Khan, N.; Maymon, M.; Hirsch, A. Combating Fusarium Infection Using Bacillus-Based Antimicrobials. Microorganisms 2017, 5, 75. [Google Scholar] [CrossRef] [Green Version]
- Syed-Ab-Rahman, S.F.; Carvalhais, L.C.; Chua, E.; Xiao, Y.; Wass, T.J.; Schenk, P.M. Identification of soil bacterial isolates suppressing different Phytophthora spp. And promoting plant growth. Front. Plant Sci. 2018, 871. [Google Scholar] [CrossRef]
- Win, T.T.; Bo, B.; Malec, P.; Fu, P. The effect of a consortium of Penicillium sp. and Bacillus sp. in suppressing banana fungal diseases caused by Fusarium sp. and Alternaria sp. J. Appl. Microbiol. 2021, jam.15067. [Google Scholar] [CrossRef]
- Freitas, M.A.; Medeiros, F.H.V.; Melo, I.S.; Pereira, P.F.; Peñaflor, M.F.G.V.; Bento, J.M.S.; Paré, P.W. Stem inoculation with bacterial strains Bacillus amyloliquefaciens (GB03) and Microbacterium imperiale (MAIIF2a) mitigates Fusarium root rot in cassava. Phytoparasitica 2019, 47, 135–142. [Google Scholar] [CrossRef]
- Parikh, L.; Eskelson, M.J.; Adesemoye, A.O. Relationship of in vitro and in planta screening: Improving the selection process for biological control agents against Fusarium root rot in row crops. Arch. Phytopathol. Plant Prot. 2018, 51, 156–169. [Google Scholar] [CrossRef]
- Abd-Elgawad, M.; El-Mougy, N.; El-Gamal, N.; Abdel-Kader, M.; Mohamed, M. Protective treatments against soilborne pathogens in citrus orchards. J. Plant Prot. Res. 2010, 50, 477–484. [Google Scholar] [CrossRef]
- Bahroun, A.; Jousset, A.; Mhamdi, R.; Mrabet, M.; Mhadhbi, H. Anti-fungal activity of bacterial endophytes associated with legumes against Fusarium solani: Assessment of fungi soil suppressiveness and plant protection induction. Appl. Soil Ecol. 2018, 124, 131–140. [Google Scholar] [CrossRef]
- Islam, S.; Akanda, A.M.; Prova, A.; Islam, M.T.; Hossain, M.M. Isolation and Identification of Plant Growth Promoting Rhizobacteria from Cucumber Rhizosphere and Their Effect on Plant Growth Promotion and Disease Suppression. Front. Microbiol. 2016, 6, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Llop, P.; Bonaterra, A.; Peñalver, J.; López, M.M. Development of a highly sensitive nested-PCR procedure using a single closed tube for detection of Erwinia amylovora in asymptomatic plant material. Appl. Environ. Microbiol. 2000, 66, 2071–2078. [Google Scholar] [CrossRef] [Green Version]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 1991, 173, 697–703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guevara-Avendaño, E.; Carrillo, J.D.; Ndinga-Muniania, C.; Moreno, K.; Méndez-Bravo, A.; Guerrero-Analco, J.A.; Eskalen, A.; Reverchon, F. Antifungal activity of avocado rhizobacteria against Fusarium euwallaceae and Graphium spp., associated with Euwallacea spp. nr. fornicatus, and Phytophthora cinnamomi. Antonie van Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2018, 111, 563–572. [Google Scholar] [CrossRef]
- Dinesh, R.; Anandaraj, M.; Kumar, A.; Bini, Y.K.; Subila, K.P.; Aravind, R. Isolation, characterization, and evaluation of multi-trait plant growth promoting rhizobacteria for their growth promoting and disease suppressing effects on ginger. Microbiol. Res. 2015, 173, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Etesami, H.; Alikhani, H.A.; Mirseyed Hosseini, H. Evaluation of halotolerant endophytic bacteria isolated from the halophyte suaeda for biological control of fungal rice pathogens. Arch. Phytopathol. Plant Prot. 2019, 52, 560–581. [Google Scholar] [CrossRef]
- Hsu, S.C.; Lockwood, J.L. Powdered Chitin Agar as a Selective Medium for Enumeration of Actinomycetes in Water and Soil1. Appl. Microbiol. 1975, 29, 422–426. [Google Scholar] [CrossRef] [PubMed]
- Velusamy, P.; Kim, K.Y. Chitinolytic activity of Enterobacter sp. KB3 antagonistic to Rhizoctonia solani and its role in the degradation of living fungal hyphae. Int. Res. J. Microbiol. 2011, 2, 206–214. [Google Scholar]
- Schwyn, B.; Neilands, J.B. Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef]
- Charest, M.H.; Beauchamp, C.J.; Antoun, H. Effects of the humic substances of de-inking paper sludge on the antagonism between two compost bacteria and Pythium ultimum. FEMS Microbiol. Ecol. 2005, 52, 219–227. [Google Scholar] [CrossRef]
- Jing, Y.X.; Yan, J.L.; He, H.D.; Yang, D.J.; Xiao, L.; Zhong, T.; Yuan, M.; de Cai, X.; Li, S. Bin Characterization of Bacteria in the Rhizosphere Soils of Polygonum Pubescens and Their Potential in Promoting Growth and Cd, Pb, Zn Uptake by Brassica napus. Int. J. Phytoremediation 2014, 16, 321–333. [Google Scholar] [CrossRef]
- Zhang, W.; Huang, Z.; He, L.; Sheng, X. Assessment of bacterial communities and characterization of lead-resistant bacteria in the rhizosphere soils of metal-tolerant Chenopodium ambrosioides grown on lead-zinc mine tailings. Chemosphere 2012, 87, 1171–1178. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Region | Bacteria | N. solani | F. oxysporum | F. brachygibbosum | F. equiseti |
|---|---|---|---|---|---|
| Taroudant | k3-7 | 64.48 ± 1.38 d–h | 54.75 ± 1.67 d,e,f | 59.87 ± 0.58 e,f,g | 9.25 ± 2.13 d,e,f |
| Taroudant | k4-3 | 66.39 ± 1.81 h,i | 53.02 ± 1.18 c,d | 53.65 ± 0.76 c,d | 59.44 ± 0.34 d,e,f |
| Taroudant | k4-4 | 72.22 ± 0.17 j | 54.08 ± 1.73 c,d,e | 56.75 ± 0.96 d,e | 68.73 ± 1.40 g |
| Sidi Kacem | Bel3-4 | 65.58 ± 0.27 e–i | 45.98 ± 0.51b | 48.60 ± 1.00 b | 50.54 ± 1.54 a,b |
| Taounate | TD1 | 71.50 ± 1.13 j | 51.53 ± 0.16 c,d | 57.72 ± 1.71 e,f | 59.12 ± 0.40 d,e,f |
| Taounate | TD7 | 63.96±1.25 b–h | 47.17 ± 1.44 b | 60.98 ± 1.00 e–h | 52.64 ± 0.72 b,c |
| Taounate | TG5 | 65.96 ± 0.29 g,h,i | 46.65 ± 0.59 b | 59.31 ± 0.87 e,f,g | 60.25 ± 0.03 e,f |
| Taounate | TG6 | 64.29 ± 1.25 c–h | 46.10 ± 0.49 b | 62.47 ± 1.51 g,h | 55.99 ± 0.60 c,d |
| Taounate | TM10 | 64.07 ± 1.70 b–h | 43.83 ± 0.37 b | 51.93 ± 1.90 b,c | 48.31 ± 0.77 a |
| Berkane | B2-1 | 62.63 ± 1.80 a–g | 38.01 ± 1.54 a | 34.48 ± 1.56 a | 56.51 ± 1.49 d,e |
| Bni Mellal | BM1-3 | 59.16 ± 0.66 a | 57.10 ± 1.46 e,f,g | 67.89 ± 0.38 j,k | 62.66 ± 0.64 f |
| Bni Mellal | BM3-2 | 65.87 ± 0.44 f–i | 57.95 ±1.26 f,g | 70.59 ± 1.18 k,l | 60.65 ±0.13 f |
| Bni Mellal | BM3-4 | 63.24 ± 0.29 b–g | 59.75 ± 1.62 g,h | 66.87 ± 1.93 i,j,k | 69.29 ± 0.70 g,h |
| Bni Mellal | BM3-5 | 60.84 ± 0.10 a–d | 67.62 ± 0.57 j | 67.01 ± 1.69 i,j,k | 66.35 ± 1.10 g |
| Bni Mellal | BM4-1 | 64.36 ± 0.28 d–h | 75.42 ± 0.75 k | 64.41 ± 1.28 h,i,j | 68.92 ± 1.68 g |
| Bni Mellal | BM4-3 | 60.63 ± 1.54 a,b,c | 68.21 ± 0.29 j | 63.11 ± 1.16 g,h,i | 69.76 ± 1.77 g,h |
| Sidi Kacem | GH1-1 | 62.22 ± 1.14 a–f | 63.21± 0.93 i | 72.12 ± 0.72 l | 72.81 ± 1.00 i |
| Sidi Kacem | GH1-2 | 61.89 ± 1.67 a–e | 51.18 ± 1.60 c | 59.94 ± 1.66 e,f,g | 59.97 ± 1.54 e,f |
| Sidi Kacem | GH1-5 | 60.49 ± 0.90 a,b | 57.52 ± 0.36 e,f,g | 57.74 ± 1.80 e,f | 59.19 ± 1.50 d,e,f |
| Sidi Kacem | GH3-8 | 68.93 ± 0.53 i,j | 62.13 ± 1.17 h,i | 61.51 ± 0.97 f,g,h | 61.04 ± 1.22 f |
| Bacterial Isolate Code | Species | Accession Numbers | Cell–Free Filtrates | VOCs |
|---|---|---|---|---|
| k3-7 | B. xiamenensis | MW843010 | 29.93 ± 0.65 a,b | 43.70 ± 0.86 i |
| k4-3 | B. licheniformis | MW843011 | 31.83 ± 1.16 b,c | 35.03 ±1.02 e |
| k4-4 | B. subtilis | MW843012 | 35.17 ± 1.55 d,e | 37.31 ± 0.97 e,f,g |
| Bel3-4 | S. multivorum | MW856827 | 30.89 ± 0.02 b | 43.82 ± 0.77 i |
| TD1 | S. maltophilia | MW856828 | 27.92 ± 0.23 a | 36.61 ± 0.93 e,f |
| TD7 | B. amyloliquefaciens | MW847947 | 32.02 ± 1.21 b,c | 20.79 ± 1.00 a,b |
| TG5 | B. subtilis | MW847627 | 32.36 ± 0.46 b,c | 22.85 ± 0.63 b,c |
| TG6 | B. subtilis | MW847628 | 37.63 ± 0.50 e,f,g | 17.97 ± 0.70 a |
| TM10 | B. velezensis | MW847948 | 34.13 ± 1.12 c,d | 39.85 ± 1.15 g,h |
| B2-1 | Stenotrophomona sp. | MW849323 | 40.70 ± 0.36 h,i,j | 24.79 ± 1.15 c,d |
| BM1-3 | B. halotolerans | MW847629 | 39.39 ± 1.31 g,h,i | 38.92 ± 0.58 f,g,h |
| BM3-2 | B. amyloliquefaciens | MW847949 | 36.63 ± 0.31 d,e,f | 25.40 ± 0.57 c,d |
| BM3-4 | B. amyloliquefaciens | MW847950 | 43.65 ± 0.35 k | 27.29 ± 0.56 d |
| BM3-5 | B. halotolerans | MW847951 | 40.69 ± 0.20 h,i,j | 19.10 ± 1.88 a |
| BM4-1 | B. halotolerans | MW847952 | 41.82 ± 0.91i,j,k | 20.25 ± 0.65 a,b |
| BM4-3 | B. halotolerans | MW847953 | 42.35 ± 0.34 j,k | 36.70 ± 1.44 e,f |
| CH1-1 | B. amyloliquefaciens | MW847630 | 39.30 ±0.15 g,h,i | 25.12 ±1.07 c,d |
| GH1-2 | B. tequilensis | MW848377 | 38.71 ±0.41 f,g,h | 27.70 ±0.55 d |
| GH1-5 | S. maltophilia | MW848819 | 41.91 ± 0.22 i,j,k | 41.10 ± 0.42 h,i |
| GH3-8 | B. subtilis | MW847631 | 36.05 ± 1.65 d,e | 48.63 ± 0.56 j |
| Isolates | PI | PrI | AI | CI | ChI | PSI | HCN | IAA | SD |
|---|---|---|---|---|---|---|---|---|---|
| k3-7 | 2.70 ± 0.05 l | 2.13 ± 0.02 d | 1.24 ± 0.09 c | 2.16 ± 0.05 d,e | 1.00 ± 0.00 a | 2.11 ± 0.09 a | − | − | − |
| k4-3 | 1.89 ± 0.02 g | 2.37 ± 0.29 e | 1.64 ± 0.05 f,g | 2.22 ± 0.04 d,e | 1.00 ± 0.00 a | 1.00 ± 0.00 f,g,h | + | − | − |
| k4-4 | 1.67 ± 0.05 f | 1.42 ± 0.09 b,c | 1.86 ± 0.01 I,j | 2.08 ± 0.04 d | 1.00 ± 0.00 a | 2.05 ± 0.03 f | + | − | + |
| Bel3-4 | 1.46 ± 0.03 c,d,e | 1.49 ± 0.14 c | 1.45 ± 0.01 d | 2.16 ± 0.03 d,e | 1.00 ± 0.00 a | 2.14 ± 0.08 g,h | − | − | + |
| TD1 | 1.42 ± 0.08 c | 2.07 ± 0.19 d | 1.55 ± 0.02 e | 2.11 ± 0.03 d | 1.85 ± 0.35 e | 2.15 ± 0.09 g,h | − | − | − |
| TD7 | 1.42 ± 0.02 c | 1.33 ± 0.14 a,b,c | 2.10 ±0.08 l | 2.62 ± 0.28 f | 1.00 ± 0.00 a | 1.00 ± 0.00 a | − | + | − |
| TG5 | 1.43 ± 0.04 c,d | 1.28 ± 0.15 a,b,c | 1.69 ± 0.03 g,h | 2.43 ± 0.21 e,f | 2.01 ± 0.32 e | 1.00 ± 0.00 a | − | + | − |
| TG6 | 1.00 ± 0.00 a | 1.15 ± 0.04 a | 1.71 ± 0.08 g,h | 1.00 ± 0.00 a | 1.00 ± 0.00 a | 1.28 ± 0.06 d | − | + | − |
| TM10 | 2.04 ± 0.01 h | 1.31 ± 0.06 a,b,c | 1.66 ± 0.03 g,h | 1.00 ± 0.00 a | 1.00 ± 0.00 a | 1.19 ± 0.04 c | − | + | − |
| B2-1 | 1.29 ± 0.03 b | 1.39 ± 0.07 b,c | 1.81 ± 0.04 i | 1.00 ± 0.00 a | 1.08 ± 0.03 a,b | 1.44 ± 0.05 e | + | − | − |
| BM1-3 | 1.00 ± 0.00 a | 1.31 ± 0.13 a,b,c | 1.92 ± 0.08 j,k | 1.47 ± 0.02 b | 1.00 ± 0.00 a | 1.14 ± 0.03 b,c | − | − | − |
| BM3-2 | 1.24 ± 0.03 b | 1.34 ± 0.08 a,b,c | 1.73 ± 0.03 h | 1.55 ± 0.00 b,c | 1.00 ± 0.00 a | 2.14 ± 0.03 g,h | + | + | − |
| BM3-4 | 1.50 ± 0.05 d,e | 1.31 ± 0.09 a,b,c | 1.58 ± 0.00 e,f | 2.23 ± 0.09 d,e | 1.41 ± 0.15 c | 1.15 ± 0.06 c | + | − | − |
| BM3-5 | 2.57 ± 0.03 k | 1.25 ± 0.11 a,b | 2.07 ± 0.05 l | 2.17 ± 0.24 d,e | 1.65 ± 0.27 d | 1.14 ± 0.04 b,c | − | − | − |
| BM4-1 | 1.52 ± 0.04 e | 1.22 ± 0.07 a,b | 1.16 ± 0.02 b | 2.29 ± 0.32 d,e | 1.27 ± 0.11 b,c | 1.00 ± 0.00 a | − | − | − |
| BM4-3 | 2.22 ± 0.02 j | 1.19 ± 0.03 a,b | 1.25 ± 0.04 c | 2.62 ± 0.23 f | 1.00 ± 0.00 a | 1.07 ± 0.03 a,b | − | + | − |
| GH1-1 | 1.46 ± 0.03 c,d,e | 1.30 ± 0.11 a,b,c | 1.03 ± 0.01 a | 1.77 ± 0.28 c | 1.00 ± 0.00 a | 1.00 ± 0.00 a | − | − | − |
| GH1-2 | 2.12 ± 0.01 i | 1.28 ± 0.03 a,b,c | 1.53 ± 0.01 e | 2.09 ± 0.01 d | 1.00 ± 0.00 a | 2.09 ± 0.01 f,g | + | + | + |
| GH1-5 | 1.60 ± 0.05 f | 1.42 ± 0.14 b,c | 1.98 ± 0.02 k | 1.00 ± 0.00 a | 1.19 ± 0.05 a,b | 1.00 ± 0.00 a | − | + | − |
| GH3-8 | 1.24 ± 0.03 b | 1.26 ± 0.07 a,b,c | 1.71 ± 0.06 g,h | 2.20 ±0.03 d,e | 1.97 ± 0.28 e | 2.19 ± 0.06 h | − | + | − |
| Bacterial Isolate | LN | TL | RL | TFW | RFW | RDW | TDW |
|---|---|---|---|---|---|---|---|
| k3-7 | 4 ± 1.00 a | 22.67 ± 2.517 c,d | 15.83 ± 1.16 f,g,h | 1.51 ± 0.07 b,c,d | 0.81 ± 0.11 d,e,f | 0.007 ± 0.001 b,c | 0.086 ± 0.007 a,b |
| k4-3 | 4 ± 0.00 a | 15.17 ± 0.76 a | 13.83 ± 1.16 d,e,f | 1.28 ± 0.55 a,b,c | 0.44 ± 0.04 a | 0.006 ± 0.002 a,b | 0.068 ± 0.013 a,b |
| k4-4 | 4 ± 0.00 a | 21.84 ± 2.57 c,d | 16.00 ± 1.80 f,g,h | 1.69 ± 0.09 d,e,f,g | 0.94 ± 0.13 f,g | 0.013 ± 0.001 e,f | 0.085 ± 0.030 a,b |
| Bel3-4 | 4 ± 0.00 a | 20.40 ± 1.44 b,c,d | 17.67 ± 1.53 h,i | 2.08 ± 0.14 g,h | 0.69 ± 0.14 b,c,d,e | 0.010 ± 0.001 d,e,f | 0.221 ± 0.059 d |
| TD1 | 3.68 ± 0.58 a | 20.17 ± 2.47 b,c,d | 13.67 ± 1.16 c,d,e,f | 1.70 ± 0.15 d,e,f,g | 0.69 ± 0.08 b–e | 0.010 ± 0.001 d,e,f | 0.102 ± 0.015 b,c |
| TD7 | 4 ± 0.00 a | 20.33 2.52 b,c,d | 18.33 ± 0.76 h,i | 2.56 ± 0.18 i | 1.10 ± 0.17 g | 0.012 ± 0.002 d,e,f | 0.19 ± 0.021 d |
| TG5 | 4 ± 0.00 a | 22.50 ± 2.29 c,d | 15.83 ± 0.29 f,g,h | 1.94 ± 0.13 e,f,g,h | 0.52 ± 0.06 a,b | 0.009 ± 0.001 c,d,e | 0.089 ± 0.017 a,b |
| TG6 | 4 ±1.00 a | 15.33 ± 1.89 a | 14.17 ± 1.04 d,e,f | 1.36 ± 0.04 a,b,c,d | 0.47 ± 0.05 a | 0.008 ± 0.001 b,c,d | 0.088 ± 0.016 a,b |
| TM10 | 4 ± 0.00 a | 19.17 ± 2.57 a–d | 14.84 ± 1.89 e,f,g | 1.92 ± 0.14 e,f,g,h | 0.85 ± 0.13 e,f | 0.010 ± 0.001 c,d,e | 0.266 ± 0.067 e |
| B2-1 | 4.33 ± 0.58 a | 22.50 ± 3.91 c,d | 19.33 ± 2.47 i | 2.12 ± 0.52 h | 0.75 ± 0.15 c,d,e,f | 0.011 ± 0.001 d,e,f | 0.269 ± 0.044 e |
| BM1-3 | 3.68 ± 0.58 a | 15.50 ± 2.29 a | 12.17 ± 2.08 b,c,d | 1.48 ± 0.06 b,c,d | 0.56 ± 0.11 a,b,c | 0.011 ± 0.002 d,e,f | 0.090 ± 0.011 a,b |
| BM3-2 | 4 ± 0.00 a | 23.10 ± 1.91 d | 13.17 ±1.06 c,d,e | 2.13 ± 0.09 h | 0.90 ± 0.15 e,f | 0.011 ± 0.002 d,e,f | 0.146 ± 0.025 c |
| BM3-4 | 4.33 ± 0.58 a | 17.60 ± 1.40 a,b | 18.17 ± 1.76 h,i | 1.64 ± 0.14 c,d,e,f | 0.64 ± 0.08 a,b,c,d | 0.005 ± 0.002 b,c | 0.070 ± 0.008 a,b |
| BM3-5 | 3.33 ± 0.58 a | 18.67 ± 2.31 a,b,c | 12.03 ± 1.62 a,b,c,d | 1.51 ± 0.05 b,c,d | 0.580 ± 0.09 a,b,c | 0.006 ± 0.003 b,c | 0.087 ± 0.014 a,b |
| BM4-1 | 4.33 ± 0.58 a | 16.67 ± 1.53 a,b | 16.83 ± 1.04 g,h,i | 1.94 ± 0.06 e,f,g,h | 1.11 ± 0.13 g | 0.013 ± 0.001 f | 0.048 ± 0.010 a |
| BM4-3 | 4 ± 0.00 a | 15.17 ± 0.76 a | 14.33 ± 1.04 d,e,f,g | 1.19 ± 0.06 a,b | 0.56 ± 0.03 a,b,c | 0.007 ± 0.005 b,c | 0.090 ± 0.014 a,b |
| GH1-1 | 4.33 ±0.58 a | 16.93 ± 2.53 a,b | 11.90 ± 0.96 a,b,c,d | 1.49 ± 0.33 b,c,d | 0.76 ± 0.07 c,d,e,f | 0.010 ± 0.002 c,d,e | 0.142 ± 0.013 c |
| GH1-2 | 4 ± 1.00 a | 17.50 ± 1.32 a,b | 11.17 ± 0.76 a,b,c | 1.57 ± 0.19 b,c,d,e | 0.95 ± 0.07 f,g | 0.012 ± 0.001 e,f | 0.103 ± 0.010 b,c |
| GH1-5 | 4 ± 0.00 a | 17.00 ± 1.32 a,b | 12.17 ± 1.26 b,c,d | 1.98 ± 0.10 f,g,h | 0.86 ± 0.18 e,f | 0.010 ± 0.002 c,d,e | 0.093 ± 0.023 a,b |
| GH3-8 | 4 ± 0.00 a | 16.00 ±2.65 a | 9.57 ±0.95 a | 1.44 ± 0.131 a,b,c,d | 0.70 ± 0.03 b,c,d,e | 0.010 ± 0.002 d,e,f | 0.107 ± 0.008 b,c |
| Untreated control | 3 ± 0.00 a | 15.60 ± 0.40 a | 10.50 ± 1.00 a,b | 1.08 ± 0.06 a | 0.73 ± 0.15 b,c,d,e | 0.003 ± 0.001 a | 0.080 ± 0.004 a,b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ezrari, S.; Mhidra, O.; Radouane, N.; Tahiri, A.; Polizzi, G.; Lazraq, A.; Lahlali, R. Potential Role of Rhizobacteria Isolated from Citrus Rhizosphere for Biological Control of Citrus Dry Root Rot. Plants 2021, 10, 872. https://doi.org/10.3390/plants10050872
Ezrari S, Mhidra O, Radouane N, Tahiri A, Polizzi G, Lazraq A, Lahlali R. Potential Role of Rhizobacteria Isolated from Citrus Rhizosphere for Biological Control of Citrus Dry Root Rot. Plants. 2021; 10(5):872. https://doi.org/10.3390/plants10050872
Chicago/Turabian StyleEzrari, Said, Oumayma Mhidra, Nabil Radouane, Abdessalem Tahiri, Giancarlo Polizzi, Abderrahim Lazraq, and Rachid Lahlali. 2021. "Potential Role of Rhizobacteria Isolated from Citrus Rhizosphere for Biological Control of Citrus Dry Root Rot" Plants 10, no. 5: 872. https://doi.org/10.3390/plants10050872