
Improved Anther Culture Media for Enhanced Callus Formation and Plant Regeneration in Rice (Oryza sativa L.)

 ,
, 

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Selection of Panicles in Booting Stage and Cold Pre-Treatment
2.3. Novel Media Compositions and Preparations
2.4. Anther Collection and Callus Induction Frequency
2.5. Plant Regeneration
2.6. Data Analyses
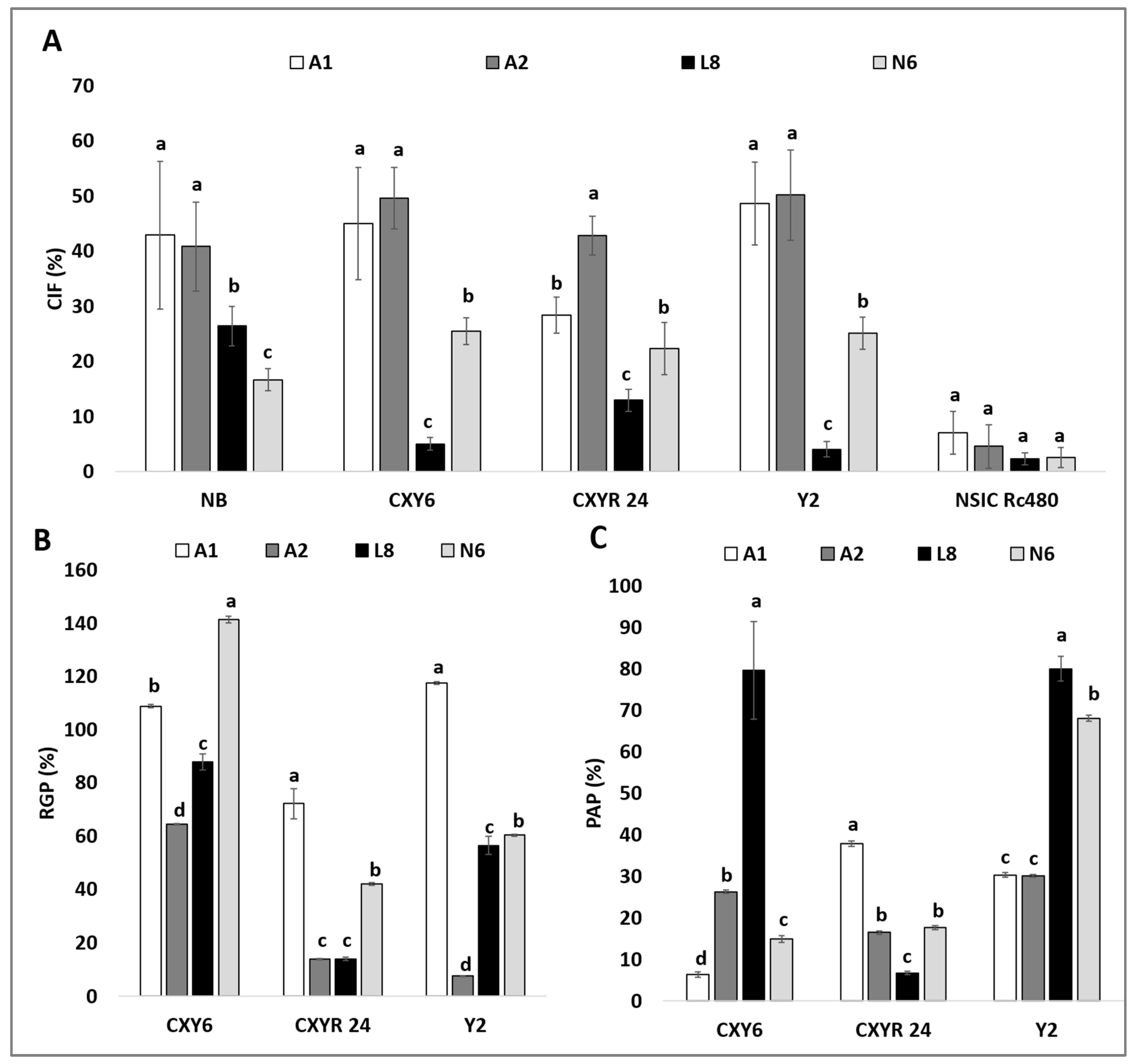
3. Results
3.1. Callus Induction Frequency in Improved Media
3.2. Effect on RGP by Using Improved Media
4. Discussion
4.1. Effect of Improved Media on Callus Induction
4.2. Effect of Improved Media on Plant Regeneration
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Rahman, Z.A.; Roowi, S.; WS, W.Z.; Subramaniam, S. Regeneration of Malaysian indica rice (Oryza sativa) variety MR232 via optimised somatic embryogenesis system. J. Phytol. 2010, 2, 30–38. [Google Scholar]
- Milovanovic, V.; Smutka, L. Asian Countries in the Global Rice Market. Acta Univ. Agric. Silvic. Mendel. Brun. 2017, 65, 679–688. [Google Scholar] [CrossRef] [Green Version]
- Conteh, A.M.H.; Yan, X.; Sankoh, F.P. The influence of price on rice production in Sierra Leone. Agric. Sci. 2012, 3, 462. [Google Scholar] [CrossRef] [Green Version]
- Brar, D.S.; Khush, G.S. Wild Relatives of Rice: A Valuable Genetic Resource for Genomics and Breeding Research. In The Wild Oryza Genomes; Springer: Cham, Switzerland, 2018; pp. 1–25. [Google Scholar] [CrossRef]
- Rao, G.J.N.; Reddy, J.N.; Variar, M.; Mahender, A. Molecular Breeding to Improve Plant Resistance to Abiotic Stresses; Springer: Berlin/Heidelberg, Germany, 2016; Volume 2, ISBN 9783319225180. [Google Scholar]
- Kumar, A.; Sandhu, N.; Yadav, S.; Pradhan, S.K.; Anandan, A.; Pandit, E.; Mahender, A.; Ram, T. Rice Varietal Development to Meet Future Challenges; Springer: Berlin/Heidelberg, Germany, 2017; ISBN 9780128139363. [Google Scholar]
- Mackill, D.J. Special Issue: Iconic Rice Varieties. Rice 2018, 11, 16. [Google Scholar] [CrossRef]
- Yorobe, J.; Pede, V.; Rejesus, R.; Velarde, O.; Wang, H.; Ali, J. Yield and Income Effects of the Green Super Rice (GSR) Varieties: Evidence from a Fixed-Effects Model in the Philippines. In Proceedings of the Agricultural & Applied Economics Association’s 2014 AAEA Annual Meeting, Minneapolis, MN, USA, 27–29 July 2014; p. 32. [Google Scholar]
- Rao, Y.; Li, Y.; Qian, Q. Recent progress on molecular breeding of rice in China. Plant Cell Rep. 2014, 33, 551–564. [Google Scholar] [CrossRef] [Green Version]
- Ashkani, S.; Rafii, M.Y.; Shabanimofrad, M.; Miah, G.; Sahebi, M.; Azizi, P.; Tanweer, F.A.; Akhtar, M.S.; Nasehi, A. Molecular breeding strategy and challenges towards improvement of blast disease resistance in rice crop. Front. Plant Sci. 2015, 6, 886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moukoumbi, Y.D.; El-Namaky, R.; Djaman, K.; Mbodj, D.; Manneh, B. Alternate phenotype–genotype selection for developing superior high-yielding irrigated rice lines. Crop J. 2018, 6, 191–201. [Google Scholar] [CrossRef]
- Srivastava, D.; Shamim, M.; Kumar, M.; Mishra, A.; Pandey, P.; Kumar, D.; Yadav, P.; Siddiqui, M.H.; Singh, K.N. Current status of conventional and molecular interventions for blast resistance in rice. Rice Sci. 2017, 24, 299–321. [Google Scholar] [CrossRef]
- Khan, M.H.; Dar, Z.A.; Dar, S.A. Breeding strategies for improving rice yield—A review. Agric. Sci. 2015, 6, 467. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Vanga, S.; Saxena, R.; Orsat, V.; Raghavan, V. Effect of climate change on the yield of cereal crops: A review. Climate 2018, 6, 41. [Google Scholar] [CrossRef] [Green Version]
- Stuecker, M.F.; Tigchelaar, M.; Kantar, M.B. Climate variability impacts on rice production in the Philippines. PLoS ONE 2018, 13, 0201426. [Google Scholar] [CrossRef] [PubMed]
- Van Oort, P.A.J.; Zwart, S.J. Impacts of climate change on rice production in Africa and causes of simulated yield changes. Glob. Chang. Biol. 2018, 24, 1029–1045. [Google Scholar] [CrossRef] [Green Version]
- Mahender, A.; Swamy, B.; Anandan, A.; Ali, J.; Mahender, A.; Swamy, B.P.M.; Anandan, A.; Ali, J. Tolerance of Iron-Deficient and -Toxic Soil Conditions in Rice. Plants 2019, 8, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, J.; Jewel, Z.A.; Mahender, A.; Anandan, A.; Hernandez, J.; Li, Z. Molecular genetics and breeding for nutrient use efficiency in rice. Int. J. Mol. Sci. 2018, 19, 1762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hawkesford, M.J.; Araus, J.; Park, R.; Calderini, D.; Miralles, D.; Shen, T.; Zhang, J.; Parry, M.A.J. Prospects of doubling global wheat yields. Food Energy Secur. 2013, 2, 34–48. [Google Scholar] [CrossRef]
- Pang, Y.; Chen, K.; Wang, X.; Wang, W.; Xu, J.; Ali, J.; Li, Z. Simultaneous improvement and genetic dissection of salt tolerance of rice (Oryza sativa L.) by designed QTL pyramiding. Front. Plant Sci. 2017, 8, 1275. [Google Scholar] [CrossRef] [Green Version]
- Dramé, K.N.; Manneh, B.; Ismail, A.M. 11 Rice Genetic Improvement for Abiotic Stress Tolerance in Africa. In Realizing Africa’s Rice Promise; CABI: Boston, MA, USA, 2013; pp. 144–160. [Google Scholar]
- Ali, J.; Xu, J.-L.L.; Gao, Y.-M.M.; Ma, X.-F.F.; Meng, L.-J.J.; Wang, Y.; Pang, Y.-L.L.; Guan, Y.-S.S.; Xu, M.-R.R.; Revilleza, J.E.; et al. Harnessing the hidden genetic diversity for improving multiple abiotic stress tolerance in rice (Oryza sativa L.). PLoS ONE 2017, 12, 0172515. [Google Scholar] [CrossRef]
- Rashid, B.; Tariq, M.; Khalid, A.; Shams, F.; Ali, Q.; Ashraf, F.; Ghaffar, I.; Khan, M.I.; Rehman, R.; Husnain, T. Crop improvement: New approaches and modern techniques. Plant Gene Trait 2017, 8, 3. [Google Scholar] [CrossRef]
- Gosal, S.S.; Kang, M.S. Plant tissue culture and genetic transformation for crop improvement. Improv. Crop Resist. Abiotic Stress 2012, 357–397. [Google Scholar] [CrossRef]
- Li, W.; Gang, L.; Deming, Z.; Feng, W.; Jiabin, C. Tissue culture system for different hybrid of indica rice. J. Northeast Agric. Univ. 2011, 18, 13–17. (In English) [Google Scholar]
- Ren, J.; Wu, P.; Trampe, B.; Tian, X.; Lübberstedt, T.; Chen, S. Novel technologies in doubled haploid line development. Plant Biotechnol. J. 2017, 15, 1361–1370. [Google Scholar] [CrossRef] [Green Version]
- Forster, B.P.; Heberle-Bors, E.; Kasha, K.J.; Touraev, A. The resurgence of haploids in higher plants. Trends Plant Sci. 2007, 12, 368–375. [Google Scholar] [CrossRef]
- Bishnoi, U.; Jain, R.K.; Rohilla, J.S.; Chowdhury, V.K.; Gupta, K.R.; Chowdhury, J.B. Anther culture of recalcitrant indica× Basmati rice hybrids. Euphytica 2000, 114, 93–101. [Google Scholar] [CrossRef]
- Dwivedi, S.L.; Britt, A.B.; Tripathi, L.; Sharma, S.; Upadhyaya, H.D.; Ortiz, R. Haploids: Constraints and opportunities in plant breeding. Biotechnol. Adv. 2015, 33, 812–829. [Google Scholar] [CrossRef]
- Prigge, V.; Xu, X.; Li, L.; Babu, R.; Chen, S.; Atlin, G.N.; Melchinger, A.E. New insights into the genetics of in vivo induction of maternal haploids, the backbone of doubled haploid technology in maize. Genetics 2012, 190, 781–793. [Google Scholar] [CrossRef] [Green Version]
- Hooghvorst, I.; Ramos-Fuentes, E.; López-Cristofannini, C.; Ortega, M.; Vidal, R.; Serrat, X.; Nogués, S. Antimitotic and hormone effects on green double haploid plant production through anther culture of Mediterranean japonica rice. Plant Cell Tissue Organ Cult. 2018, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Rout, P.; Naik, N.; Ngangkham, U.; Verma, R.L.; Katara, J.L.; Singh, O.N.; Samantaray, S. Doubled Haploids generated through anther culture from an elite long duration rice hybrid, CRHR32: Method optimization and molecular characterization. Plant Biotechnol. 2016, 33, 177–186. [Google Scholar] [CrossRef] [Green Version]
- Naik, N.; Rout, P.; Umakanta, N.; Verma, R.L.; Katara, J.L.; Sahoo, K.K.; Singh, O.N.; Samantaray, S. Development of doubled haploids from an elite indica rice hybrid (BS6444G) using anther culture. Plant Cell Tissue Organ Cult. 2017, 128, 679–689. [Google Scholar] [CrossRef]
- Brar, D.S.; Khush, G.S. Cytogenetic manipulation and germplasm enhancement of rice (Oryza sativa L.). Genet. Resour. Chromosom. Eng. Crop Improv. 2006, 2, 115–158. [Google Scholar]
- Chen, Q.F.; Wang, C.L.; Lu, Y.M.; Shen, M.; Afza, R.; Duren, M.V.; Brunner, H. Anther culture in connection with induced mutations for rice improvement. Euphytica 2001, 120, 401–408. [Google Scholar] [CrossRef]
- Xa, T.T.; Lang, N.T. Rice breeding for high grain quality through anther culture. Omonrice 2011, 18, 68–72. [Google Scholar]
- Mishra, R.; Rao, G.J.N. In-vitro androgenesis in rice: Advantages, constraints and future prospects. Rice Sci. 2016, 23, 57–68. [Google Scholar] [CrossRef] [Green Version]
- Mayakaduwa, D.; Silva, T.D. A cytological indicator allows rapid assessment of microspore maturity, leading to improved in vitro anther response in Indica rice (Oryza sativa L.). In Vitro Cell. Dev. Biol. 2017, 53, 591–597. [Google Scholar] [CrossRef]
- Maluszynski, M.; Kasha, K.J.; Szarejko, I. Published doubled haploid protocols in plant species. In Doubled Haploid Production in Crop Plants; Springer: Dordrecht, The Netherlands, 2003; pp. 309–335. [Google Scholar] [CrossRef]
- Dunwell, J.M. Haploids in flowering plants: Origins and exploitation. Plant Biotechnol. J. 2010, 8, 377–424. [Google Scholar] [CrossRef] [PubMed]
- López-Cristoffanini, C.; Serrat, X.; Ramos-Fuentes, E.; Hooghvorst, I.; Llaó, R.; López-Carbonell, M.; Nogués, S. An improved anther culture procedure for obtaining new commercial Mediterranean temperate japonica rice (Oryza sativa) genotypes. Plant Biotechnol. 2018, 32, 161–166. [Google Scholar] [CrossRef] [Green Version]
- Pauk, J.; Jancso, M.; Simon-Kiss, I. Rice doubled haploids and breeding. In Advances in Haploid Production in Higher Plants; Springer: Berlin/Heidelberg, Germany, 2009; pp. 189–197. [Google Scholar]
- Takamori, L.M.; Neto, N.B.M.; Vieira, L.G.E.; Ribas, A.F. Optimization of somatic embryogenesis and in vitro plant regeneration of Urochloa species using picloram. In Vitro Cell. Dev. Biol. 2015, 51, 554–563. [Google Scholar] [CrossRef]
- Chen, C.-C.; Tsay, H.-S.; Huang, C.-R. Factors affecting androgenesis in rice (Oryza sativa L.). In Rice; Springer: Berlin/Heidelberg, Germany, 1991; pp. 193–215. [Google Scholar]
- He, T.; Yang, Y.; Tu, S.B.; Yu, M.Q.; Li, X.F. Selection of interspecific hybrids for anther culture of indica rice. Plant Cell Tissue Organ Cult. 2006, 86, 271–277. [Google Scholar] [CrossRef] [Green Version]
- Herath, H.M.I.; Bandara, D.C.; Samarajeewa, P.K. Effect of culture media for anther culture of indica rice varieties and hybrids of Indica and Japonica. Trop. Agric. Res. Ext. 2007, 10, 17–22. [Google Scholar] [CrossRef] [Green Version]
- Kaushal, L.; Balachandran, S.M.; Ulaganathan, K.; Shenoy, V. Effect of culture media on improving anther culture response of rice (Oryza sativa L.). Int. J. Agric. Innov. Res. 2014, 3, 218–224. [Google Scholar]
- Javed, M.A.; Ishii, T.; Kamijima, O.; Misoo, S. The role of alternating culture temperatures and maltose in enhancing the anther culture efficiency of salt tolerant indica rice (Oryza sativa L.) cultivars, Pokkali and Nona Bokra. Plant Biotechnol. 2007, 24, 283–287. [Google Scholar] [CrossRef]
- Islam, S.M. The Effect of Colchicine Pretreatment on Isolated Microspore Culture of Wheat (‘Triticum aestivum’L.). Aust. J. Crop Sci. 2010, 4, 660. [Google Scholar]
- Trejo-Tapia, G.; Amaya, U.M.; Morales, G.S.; Sánchez, A.D.J.; Bonfil, B.M.; Rodríguez-Monroy, M.; Jiménez-Aparicio, A. The effects of cold-pretreatment, auxins and carbon source on anther culture of rice. Plant Cell Tissue Organ Cult. 2002, 71, 41–46. [Google Scholar] [CrossRef]
- Zaidi, M.A.; Narayanan, M.; Sardana, R.; Taga, I.; Postel, S.; Johns, R.; McNulty, M.; Mottiar, Y.; Mao, J.; Loit, E. Optimizing tissue culture media for efficient transformation of different indica rice genotypes. Agron. Res. 2006, 4, 563–575. [Google Scholar]
- Raina, S.K.; Zapata, F.J. Enhanced anther culture efficiency of indica rice (Oryza sativa L.) through modification of the culture media. Plant Breed. 1997, 116, 305–315. [Google Scholar] [CrossRef]
- Faruq, G.; Shamsuddin, F.; Jenifer, A.; Jusoh, S.H.; Nezhadahmadi, A.; Khalid, N. Optimization of Media and Cold Pretreatment for Anther Culture Using Japonica/Indica and Indica/Indica Hybrids and their Callus Induction Comparisons in Different Rice Crosses. Indian J. Sci. Technol. 2014, 7, 1861–1870. [Google Scholar] [CrossRef]
- Gioi, T.D.; Tuan, V.D. Effect of different media and genotypes on anther culture efficiency of F1 plants derived from crosses between IR64 and new plant type rice cultivars. Omonrice 2002, 10, 107–109. [Google Scholar]
- Sah, S.K.; Kaur, A.; Singh, J. High frequency embryogenic callus induction and whole plant regeneration in japonica rice Cv. Kitaake. Rice Res. Open Access 2014. [Google Scholar] [CrossRef]
- Chu, C.-C.; Wang, C.-C.; Sun, C.-S.; Hsu, C.; Yin, K.-C.; Chu, C.-Y.; Bi, F.-Y. Establishment of an efficient medium for anther culture of rice through comparative experiments on the nitrogen sources. Sci. Sin. 1975, 18, 659–668. [Google Scholar]
- Roy, B. Anther culture response in indica rice and variations in major agronomic characters among the androclones of a scented cultivar, Karnal local. Afr. J. Biotechnol. 2005, 4, 235–240. [Google Scholar]
- Gioi, T.D.; Tuan, V.D. Anther Culture from Crosses between IR64 and New Plant Type Cultivar. Omonrice 2004, 12, 27–32. [Google Scholar]
- Vijava Laxmi, G.; Reddy, G.M. Anther culture of indica rice. Technical improvements in calluses induction and green plant regeneration [Oryza sativa L.]. J. Genet. Breed. 1997, 51, 295–301. [Google Scholar]
- Linsmaier, E.M.; Skoog, F. Organic growth factor requirements of tobacco tissue cultures. Physiol. Plant. 1965, 18, 100–127. [Google Scholar] [CrossRef]
- Lapitan, V.P.C.; Villegas, V.N. Anther culture response of some rice genotypes with varying amylose content to nitrogen fertilization. Philipp. J. Crop Sci. 1999, 24, 79–84. [Google Scholar]
- Silva, T.D.; Ratnayake, W.J. Another culture potential of indica rice varieties, KURULU THUDA and BG 250. Trop. Agric. Res. Ext. 2009, 12, 53–56. [Google Scholar] [CrossRef] [Green Version]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Yoshida, S.; Forno, D.A.; Cock, J.H. Laboratory Manual for Physiological Studies of Rice; THE INTERNATIONAL RICE RESEARCH INSTITUTE: LosBaños, Philippines, 1971. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; Version 4.0.3; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 20 March 2021).
- Germana, M.A. Doubled haploid production in fruit crops. Plant Cell Tissue Organ Cult. 2006, 86, 131. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.-F.; Cui, L.; Malik, A.A.; Mbira, K.G. In vitro haploid and dihaploid production via unfertilized ovule culture. Plant Cell Tissue Organ Cult. 2011, 104, 311–319. [Google Scholar] [CrossRef]
- Murovec, J.; Bohanec, B. Haploids and doubled haploids in plant breeding. In Plant Breeding; InTechOpen: London, UK, 2012; Available online: https://www.intechopen.com/books/plant-breeding/haploids-and-doubled-haploids-in-plant-breeding (accessed on 15 April 2021). [CrossRef] [Green Version]
- Germana, M.A. Gametic embryogenesis and haploid technology as valuable support to plant breeding. Plant Cell Rep. 2011, 30, 839–857. [Google Scholar] [CrossRef]
- Żur, I.; Dubas, E.; Krzewska, M.; Janowiak, F. Current insights into hormonal regulation of microspore embryogenesis. Front. Plant Sci. 2015, 6, 424. [Google Scholar]
- Chong-Pérez, B.; Carrasco, B.; Silva, H.; Herrera, F.; Quiroz, K.; Garcia-Gonzales, R. Regeneration of highland papaya (Vasconcellea pubescens) from anther culture. Appl. Plant Sci. 2018, 6, 01182. [Google Scholar] [CrossRef] [Green Version]
- Zapata-Arias, F.J. Laboratory protocol for anther culture technique in rice. In Doubled Haploid Production in Crop Plants; Springer: Dordrecht, The Netherlands, 2003; pp. 109–116. [Google Scholar] [CrossRef]
- Lentini, Z.; Reyes, P.; Martínez, C.P.; Roca, W.M. Androgenesis of highly recalcitrant rice genotypes with maltose and silver nitrate. Plant Sci. 1995, 110, 127–138. [Google Scholar] [CrossRef]
- Herath, H.M.I.; Bandara, D.C.; Samarajeewa, P.K.; Wijesundara, D.S.A. The effect of plant growth regulators on anther culture response and plant regeneration in selected Sri Lankan Indica rice varieties, Japonica varieties and their inter-sub specific hybrids. Trop. Agric. Res. 2008, 20, 243–250. [Google Scholar]
- Medhabati, K.; Das, K.R.; Henary, C.; Singh, T.D.; Sunitibala, H. Androgenic callus induction of the indica rice hybrid of Chakhao Amubi and Basmati 370. Int. Res. J. Biol. Sci. 2014, 3, 73–79. [Google Scholar]
- Karim, N.H.; Shahjahan, A.K.M.; Nahar, M.A.; Miah, S.A.; Haque, M.Z. Improved media for callus induction from anthers of indica rice (Oryza sativa L.). Plant Tissue Cult. 1991, 1, 43–50. [Google Scholar]
- Talebi, R.; Rahemi, M.R.; Arefi, H.; Nourozi, M.; Bagheri, N. In vitro plant regeneration through anther culture of some Iranian local rice (Oryza sativa L.) cultivars. Pak. J. Biol. Sci. 2007, 10, 2056–2060. [Google Scholar]
- Sen, C.; Singh, R.P. Anther culture response in boro rice hybrids. Asian J. Biotechnol. 2011, 3, 470–477. [Google Scholar] [CrossRef] [Green Version]
- Cardoza, V.; Stewart, C.N. Brassica biotechnology: Progress in cellular and molecular biology. In Vitro Cell. Dev. Biol. 2004, 40, 542–551. [Google Scholar] [CrossRef]
- Kumar, G.P.; Sivakumar, S.; Siva, G.; Vigneswaran, M.; Kumar, T.S.; Jayabalan, N. Silver nitrate promotes high-frequency multiple shoot regeneration in cotton (Gossypium hirsutum L.) by inhibiting ethylene production and phenolic secretion. In Vitro Cell. Dev. Biol. 2016, 52, 408–418. [Google Scholar]
- Dias, J.S.; Martins, M.G. Effect of silver nitrate on anther culture embryo production of different Brassica oleracea morphotypes. Sci. Hortic. (Amst.) 1999, 82, 299–307. [Google Scholar] [CrossRef]
- Cristea, T.O.; Leonte, C.; Brezeanu, C.; Brezeanu, M.; Ambarus, S.; Calin, M.; Prisecaru, M. Effect of AgNO3 on androgenesis of Brassica oleracea L. anthers cultivated in vitro. Afr. J. Biotechnol. 2012, 11, 13788–13795. [Google Scholar]
- Sandhu, J.S.; Gill, M.S.; Gosal, S.S. Callus induction and plant regeneration from cultured anthers of indica rice varieties. Plant Tissue Cult. 1993, 3, 17–21. (In India) [Google Scholar]
- Yruela, I. Copper in plants: Acquisition, transport and interactions. Funct. Plant Biol. 2009, 36, 409–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thadavong, S.; Sripichitt, P.; Wongyai, W.; Jompuk, P. Callus induction and plant regeneration from mature embryos of glutinous rice (Oryza sativa L.) cultivar TDK1. Kasetsart J. (Nat. Sci.) 2002, 36, 334–344. [Google Scholar]
- Anitha, S.K.; Reddy, G.M. Anther culture studies and selection of salt tolerant cell lines from indica rice. Proc. Indian Natl. Sci. Acad. Part B 1997, 63, 99–106. [Google Scholar]
- Sengsai, S.; Peyachoknagul, S.; Sripichitt, P.; Thongpan, A.; Pongtongkam, P. Anther culture of BC1F1 (KDML105/IRBB5/KDML105) hybrid to produce bacterial blight resistance double haploid rice. Kasetsart J. 2007, 41, 251–261. [Google Scholar]
- Rukmini, M.; Rao, G.J.N.; Rao, R.N. Effect of cold pretreatment and phytohormones on anther culture efficiency of two Indica rice (Oryza sativa L.) Hybrids-Ajay and Rajalaxmi. J. Exp. Biol. Agric. Sci. 2013, 1, 69–76. [Google Scholar]
- Mendoza, M.G.; Kaeppler, H.F. Auxin and sugar effects on callus induction and plant regeneration frequencies from mature embryos of wheat (Triticum aestivum L.). In Vitro Cell. Dev. Biol. 2002, 38, 39–45. [Google Scholar] [CrossRef]
- Gamborg, O.L.; Miller, R.; Ojima, K. Nutrient requirements of suspension cultures of soybean root cells. Exp. Cell Res. 1968, 50, 151–158. [Google Scholar] [CrossRef]
- Rathore, S.; Singh, N.; Singh, S.K. Role of sucrose and season on rapid in vitro regeneration for two Stevia genotypes. GJBB 2013, 2, 150–153. [Google Scholar]
- Swedlund, B.; Locy, R.D. Sorbitol as the primary carbon source for the growth of embryogenic callus of maize. Plant Physiol. 1993, 103, 1339–1346. [Google Scholar] [CrossRef] [Green Version]
- Shahnewaz, S.; Bari, M.A. Effect of concentration of sucrose on the frequency of callus induction and plant regeneration in anther culture of rice (Oryza sativa L.). Plant Tissue Cult. 2004, 14, 37–43. [Google Scholar]
- Lal, D.; Shashidhar, H.E.; Godwa, P.H.R.; Ashok, T.H. Callus induction and regeneration from in vitro anther culture of rice (Oryza sativa L.). Int. J. Agric. Environ. Biotechnol. 2014, 7, 213. [Google Scholar] [CrossRef]
- Ikeuchi, M.; Sugimoto, K.; Iwase, A. Plant callus: Mechanisms of induction and repression. Plant Cell 2013, 25, 3159–3173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Islam, M.M.; Adhikary, S.K.; Gain, P.; Rahman, M.; Siddique, N.-A. Effect of plant growth regulators on callus induction and plant regeneration in anther culture of rice. Pakistan J. Biol. Sci. 2004, 7, 331–334. [Google Scholar] [CrossRef] [Green Version]
- Pande, H.; Bhojwani, S.S. Promotion of androgenesis in rice anther cultures by substitution of sucrose with maltose and mannitol. Biol. Plant. 1999, 42, 125–128. [Google Scholar] [CrossRef]
- Bagheri, N.; Babaeian-Jelodar, N.; Ghanbari, A. Evaluation of effective factors in anther culture of Iranian rice (Oryza sativa L.) cultivars. Biharean Biol. 2009, 3, 119–124. [Google Scholar]
- Silva, T.D. Indica rice anther culture: Can the impasse be surpassed? Plant Cell Tissue Organ Cult. 2010, 100, 1–11. [Google Scholar] [CrossRef]
- Kumari, M.; Clarke, H.J.; Small, I.; Siddique, K.H.M. Albinism in plants: A major bottleneck in wide hybridization, androgenesis and doubled haploid culture. Crit. Rev. Plant Sci. 2009, 28, 393–409. [Google Scholar] [CrossRef]
- He, P.; Shen, L.; Lu, C.; Chen, Y.; Zhu, L. Analysis of quantitative trait loci which contribute to anther culturability in rice (Oryza sativa L.). Mol. Breed. 1998, 4, 165–172. [Google Scholar] [CrossRef]
- Chen, C.-C. Effects of Sucrose Concentration on Plant Production in Anther Culture of Rice 1. Crop Sci. 1978, 18, 905–906. [Google Scholar] [CrossRef]
- Bednarek, P.T.; Orłowska, R. Time of in vitro anther culture may moderate action of copper and silver ions that affect the relationship between DNA methylation change and the yield of barley green regenerants. Plants 2020, 9, 1064. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Chemicals | Concentration (mg L−1) | |||
|---|---|---|---|---|
| L8 | N6 | Ali-1 (A1) (Modified N6) | Ali-2 (A2) (Modified N6) | |
| Macronutrients | ||||
| MgSO4·7H2O | 185.00 | 185.00 | 185.00 | 200.00 * |
| KH2PO4 | 540.00 | 400.00 | 400.00 | 500.00 * |
| KNO3 | 3000.00 | 2830.00 | 2830.00 | 3000.00 * |
| CaCl2·2H2O | 150.00 | 166.00 | 166.00 | 150.00 * |
| (NH4)2SO4 | - | 463.00 | 463.00 | 300.00 * |
| Micronutrients | ||||
| H3BO3 | 6.00 | 1.60 | 1.60 | 2.00 * |
| MnSO4·4H2O | 17.98 | 4.40 | 4.40 | 5.00 * |
| ZnSO4·7H2O | 10.00 | 1.85 | 1.50 * | 3.00 * |
| Na2MoO4·2H2O | 0.250 | - | - | 0.250 # |
| CuSO4·5H2O | 0.025 | - | - | 0.025 # |
| CoCl2·6H2O | 0.025 | - | - | 0.025 # |
| KI | 0.83 | 0.80 | 0.80 | 0.83 * |
| FeSO4·7H2O | 27.85 | 27.80 | 27.80 | 43.00 * |
| Na2EDTA·2H2O | 37.25 | 37.30 | 37.30 | 56.00 * |
| AgNO3 | - | - | - | 10.00 # |
| Vitamins | ||||
| Nicotinic acid | 3.00 | 0.50 | 0.50 | 2.00 * |
| Thiamine HCl | 2.50 | 1.00 | 1.00 | 4.00 * |
| Pyridoxine HCl | 5.00 | 0.50 | 0.50 | 2.00 |
| Myo-inositol | 100.00 | - | 100.00 # | 100.00 # |
| Others | ||||
| Sucrose | 50,000.00 | 60,000.00 | 30,000.00 * | 10,000.00 * |
| Sorbitol | - | - | 10,000.00 # | |
| Maltose | - | - | 30,000.00 # | 40,000.00 # |
| Glycine | - | 10.00 | 10.00 | 10.00 |
| Phytagel | 8000.00 | 2000.00 | 2000.00 | 2000.00 |
| YE | - | - | - | 1000.00 # |
| Lactalbumin hydrolysate | 300.00 | - | - | - |
| Phytohormones | ||||
| 2,4-D | 0.50 | 2.00 | 1.00 * | 1.00 * |
| NAA | 3.50 | - | 1.00 # | 1.00 # |
| Kinetin | 2.00 | - | - | |
| Zeatin | - | - | 0.10 # | 0.10 # |
| pH | 5.80 | 5.80 | 5.80 | 5.80 |
| S. no | Sources of Variation | Degrees of Freedom | Sum of Squares | Mean Square |
|---|---|---|---|---|
| 1 | Genotype | 4 | 11,443 | 2861 *** |
| 2 | Media | 3 | 12,779 | 4260 *** |
| 3 | Genotype x media | 12 | 5395 | 450 *** |
| 4 | Residuals | 80 | 2497 | 31 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ali, J.; Nicolas, K.L.C.; Akther, S.; Torabi, A.; Ebadi, A.A.; Marfori-Nazarea, C.M.; Mahender, A. Improved Anther Culture Media for Enhanced Callus Formation and Plant Regeneration in Rice (Oryza sativa L.). Plants 2021, 10, 839. https://doi.org/10.3390/plants10050839
Ali J, Nicolas KLC, Akther S, Torabi A, Ebadi AA, Marfori-Nazarea CM, Mahender A. Improved Anther Culture Media for Enhanced Callus Formation and Plant Regeneration in Rice (Oryza sativa L.). Plants. 2021; 10(5):839. https://doi.org/10.3390/plants10050839
Chicago/Turabian StyleAli, Jauhar, Katrina Leslie C. Nicolas, Shahana Akther, Azerkhsh Torabi, Ali Akbar Ebadi, Corinne M. Marfori-Nazarea, and Anumalla Mahender. 2021. "Improved Anther Culture Media for Enhanced Callus Formation and Plant Regeneration in Rice (Oryza sativa L.)" Plants 10, no. 5: 839. https://doi.org/10.3390/plants10050839