
Anti-Obesity Effect of Pine Needle Extract on High-Fat Diet-Induced Obese Mice


 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
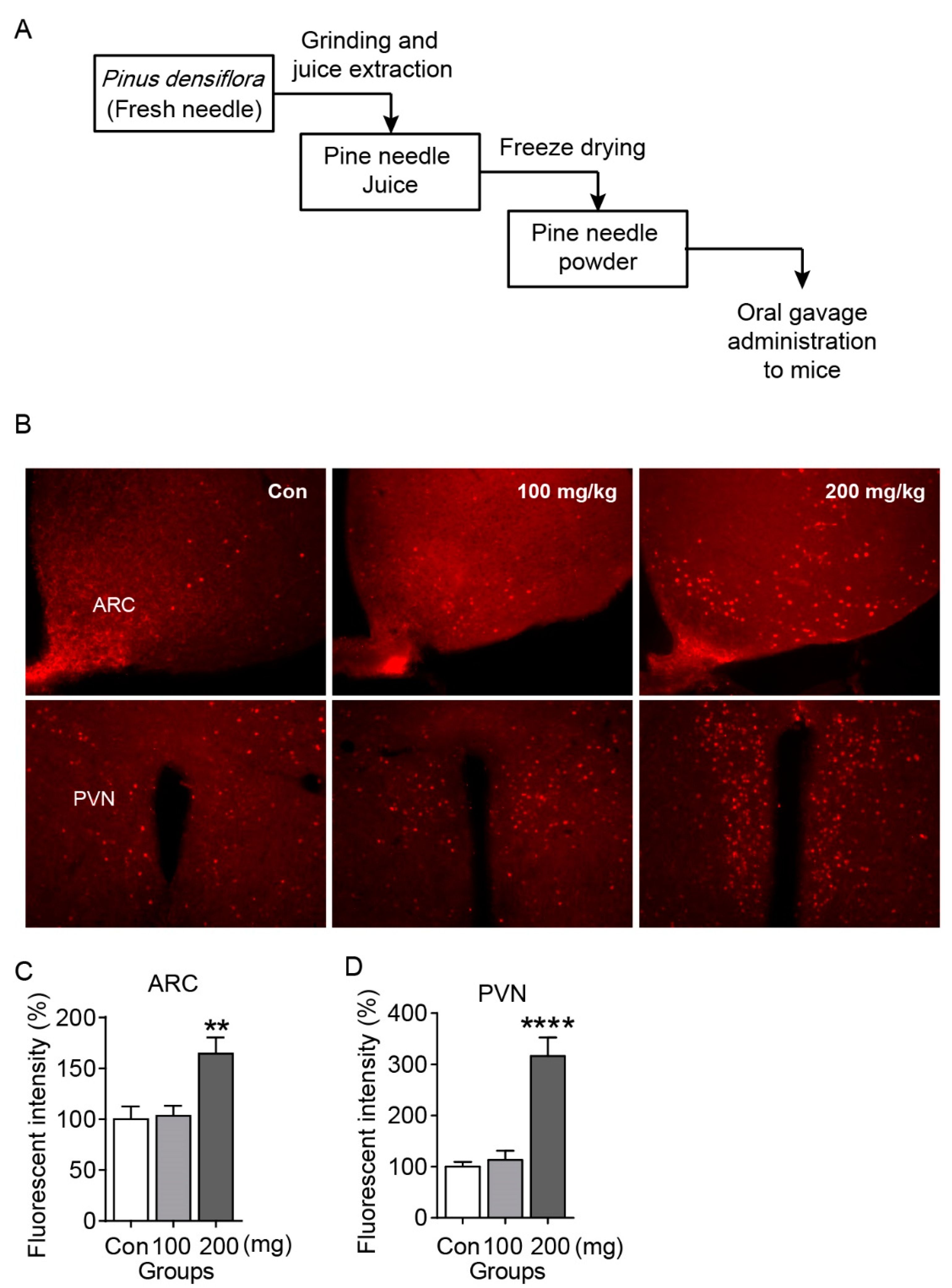
2.1. PNE Increases C-Fos Expression in Both ARC and PVN of the Hypothalamus
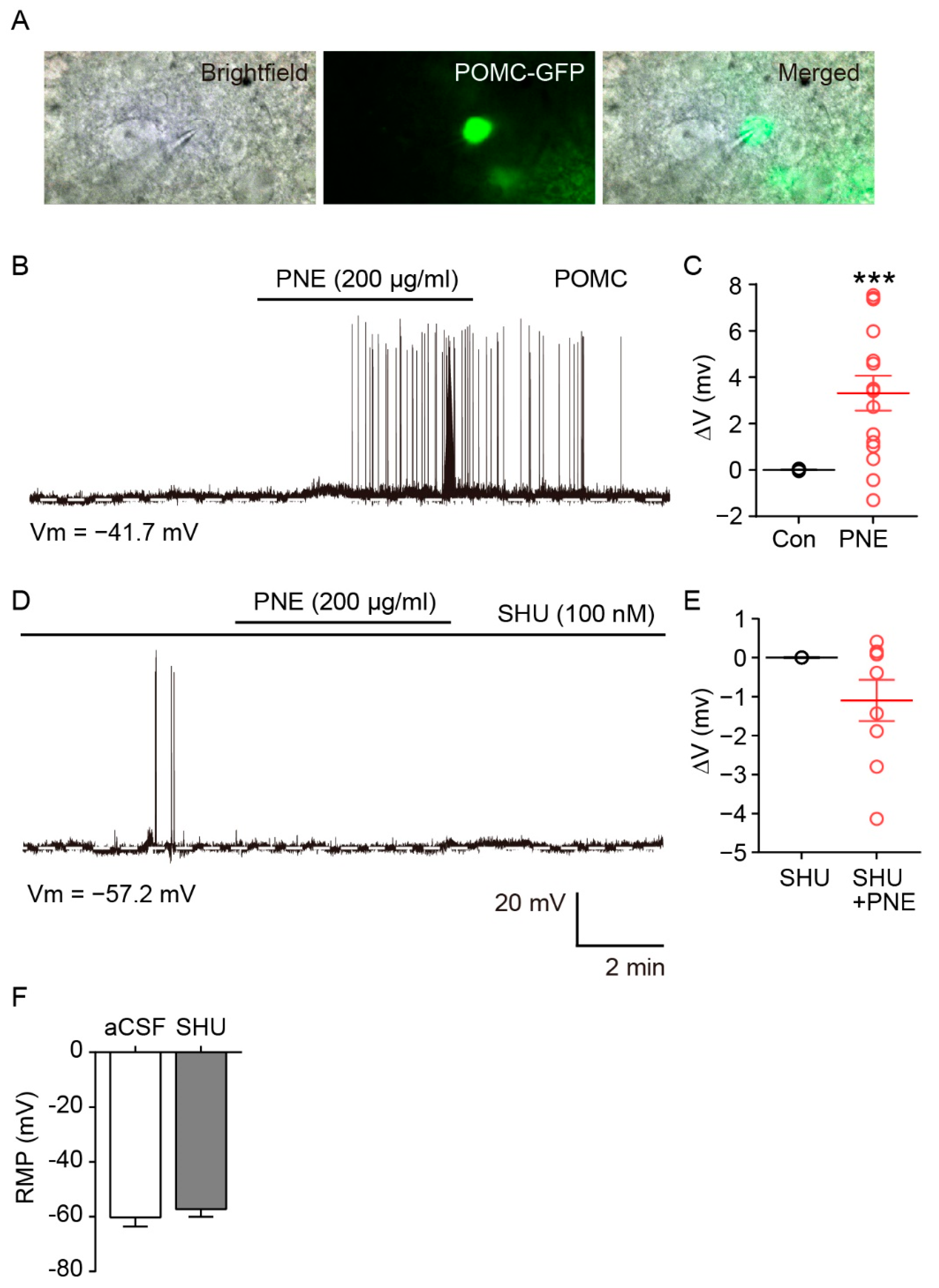
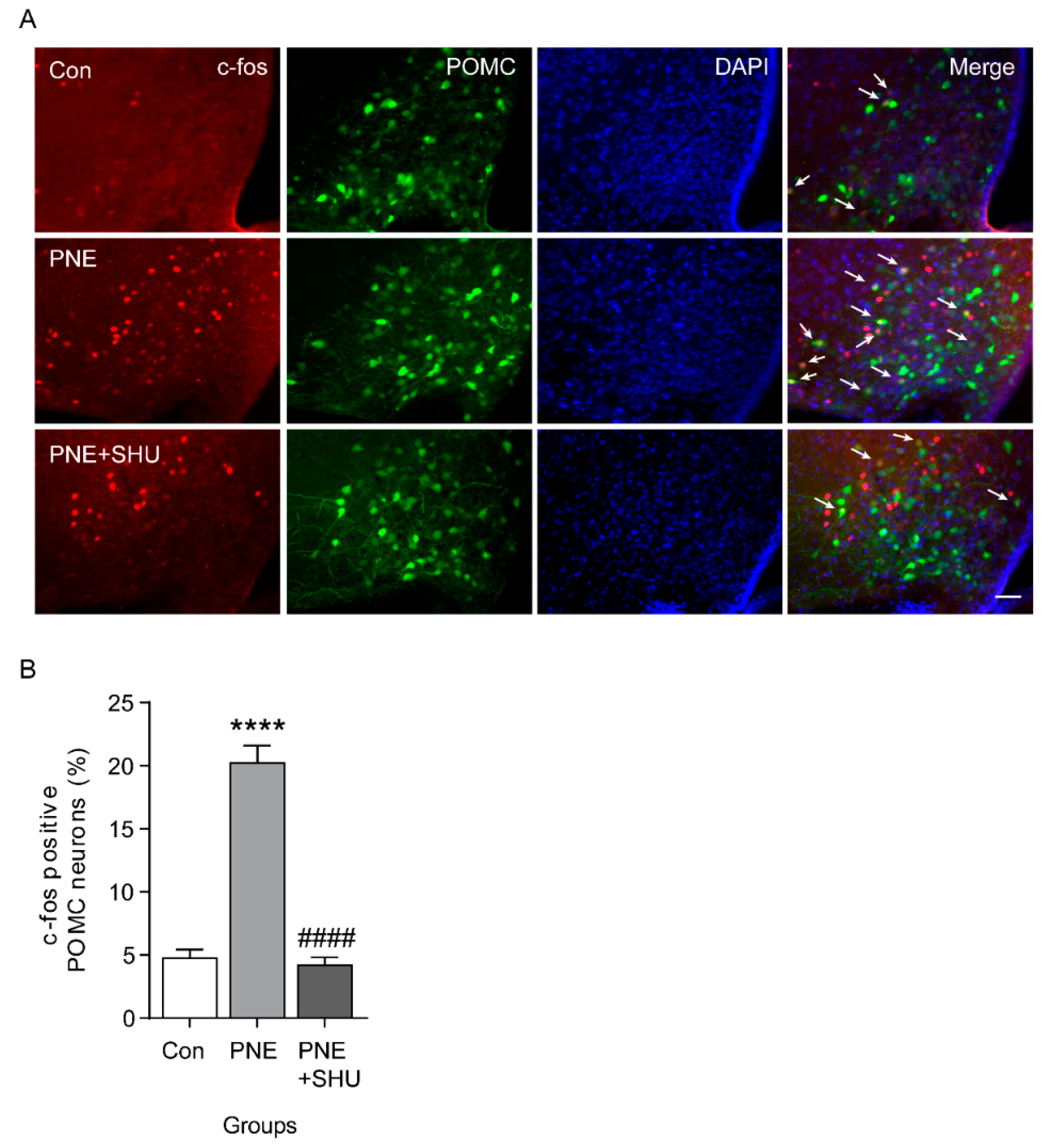
2.2. Blockade of PNE-Evoked Depolarization of ARC POMC Neurons by MC4Rs Inhibition
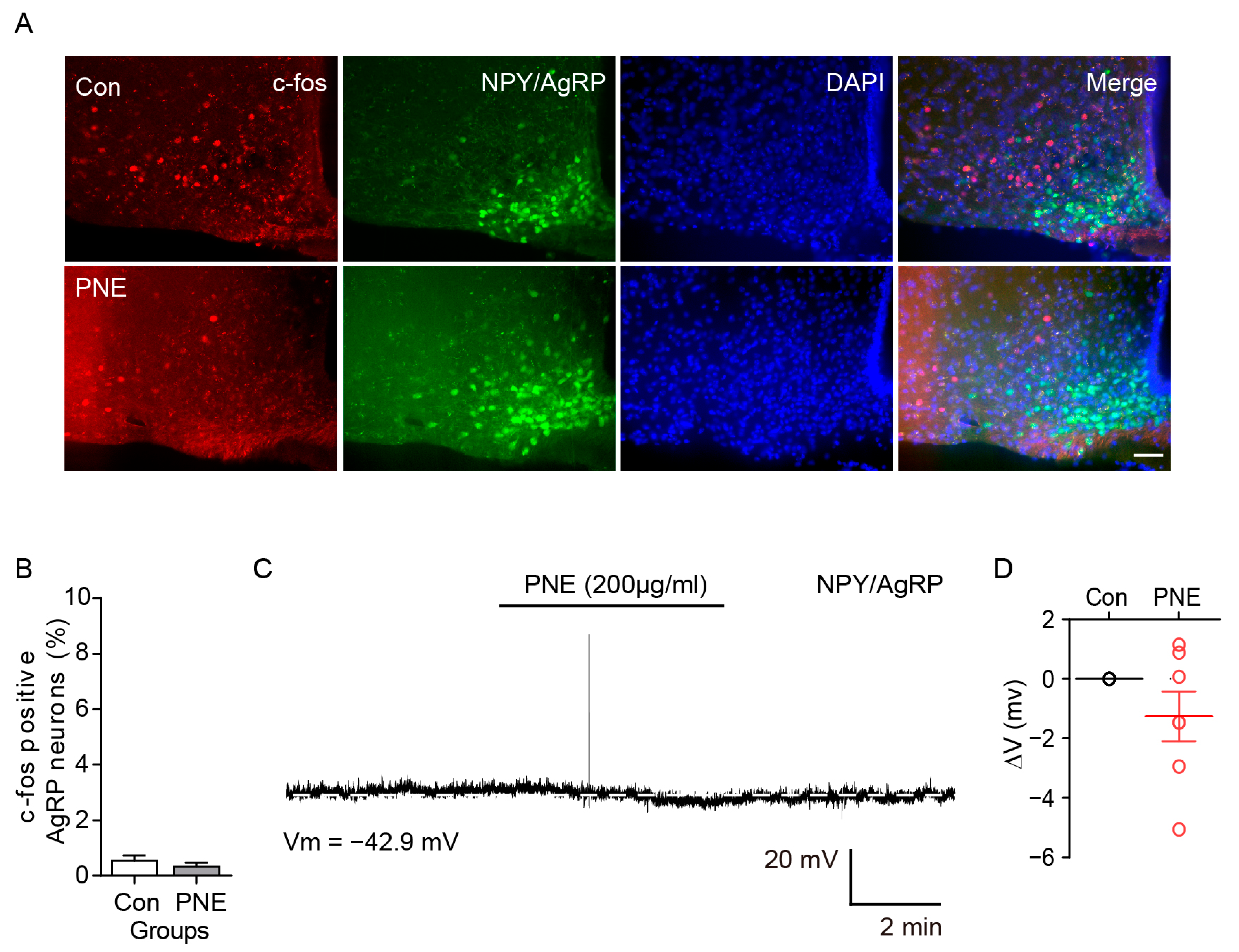
2.3. PNE Does Not Change the ARC NPY/AgRP Neuronal Activity in the Hypothalamus
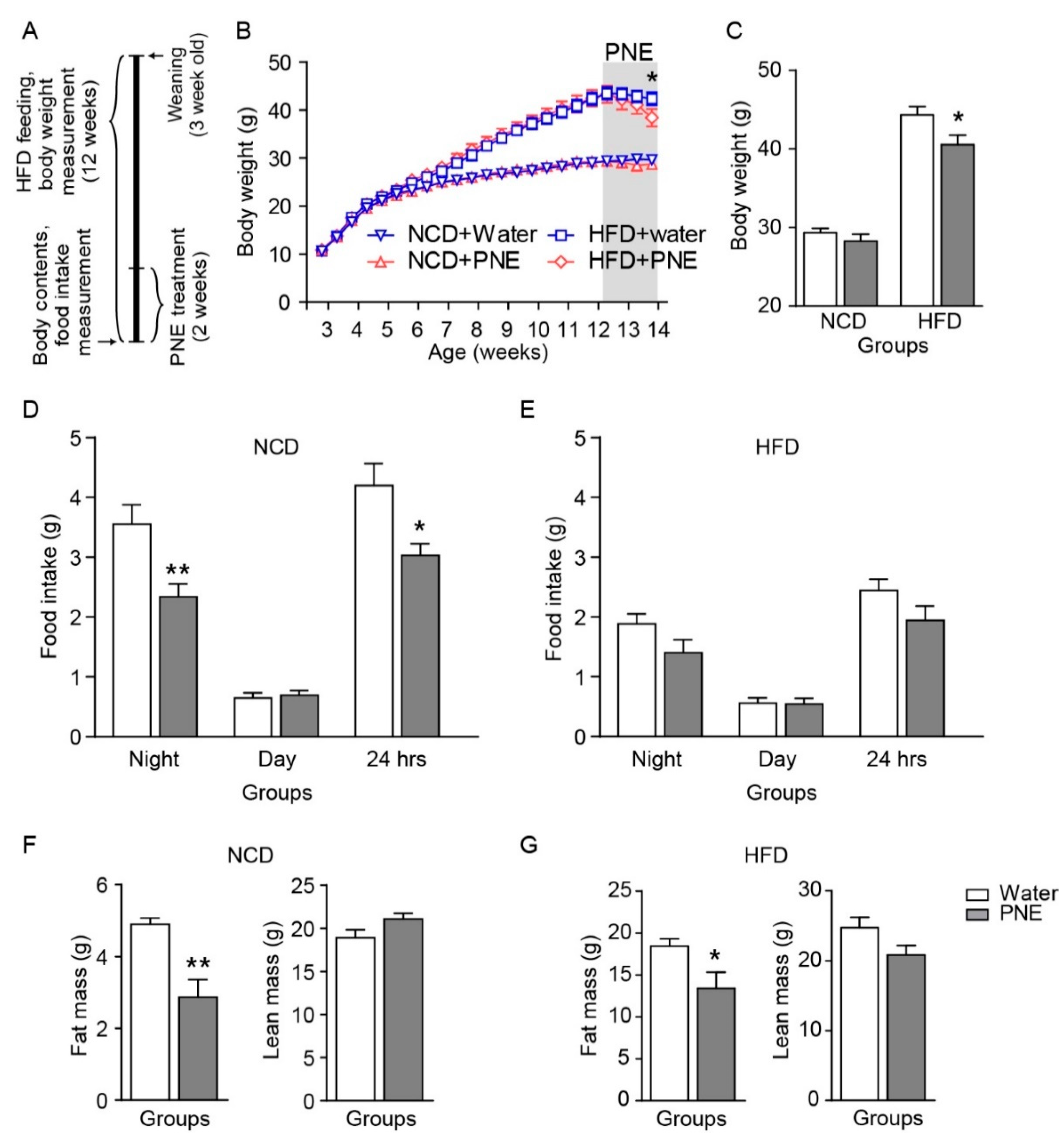
2.4. PNE Reduces Fat Mass but Not Feeding Behavior in HFD Fed Mice
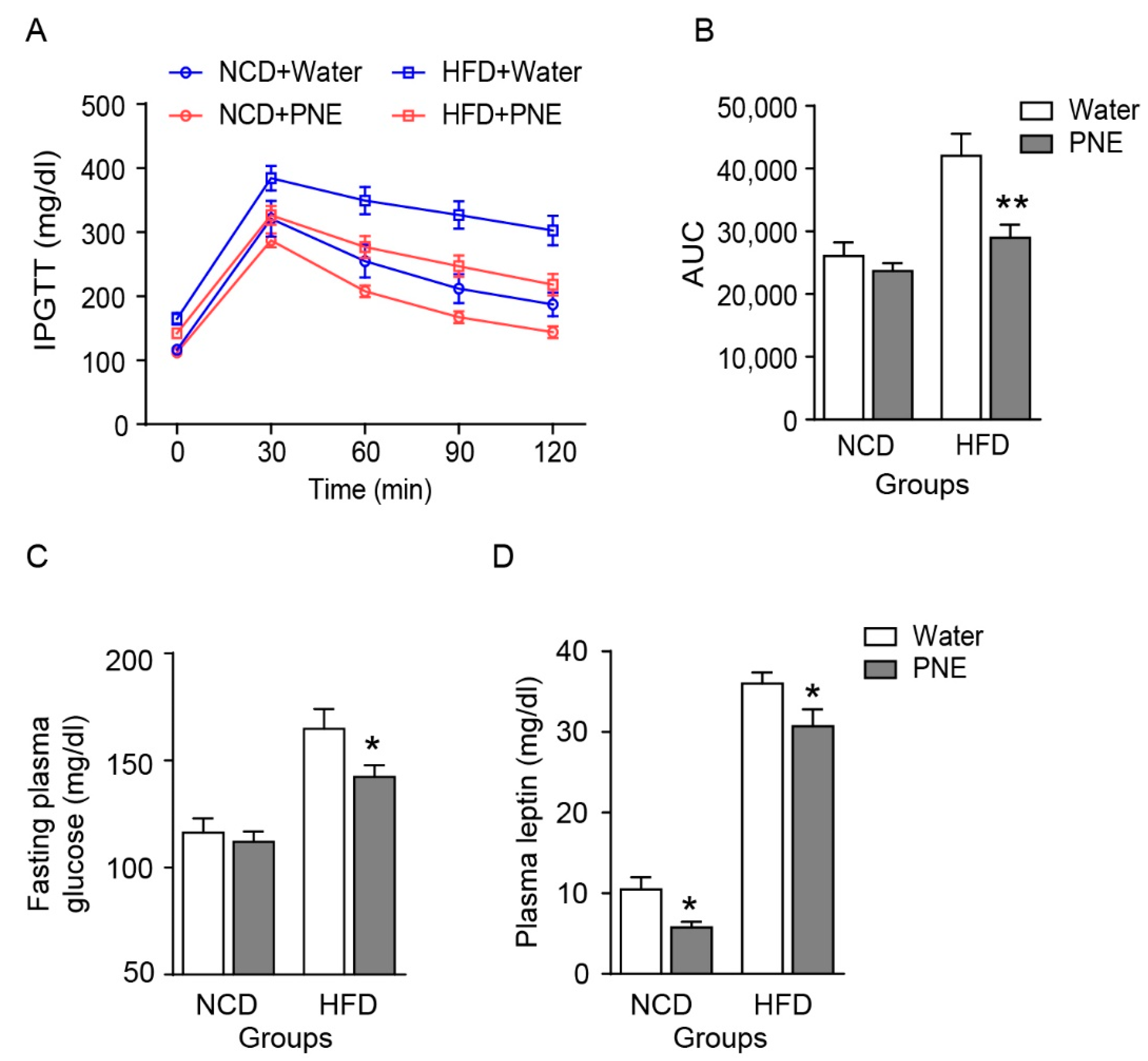
2.5. PNE Improves Glucose Tolerance by Regulating Leptin Levels in HFD-Induced Obese Mice
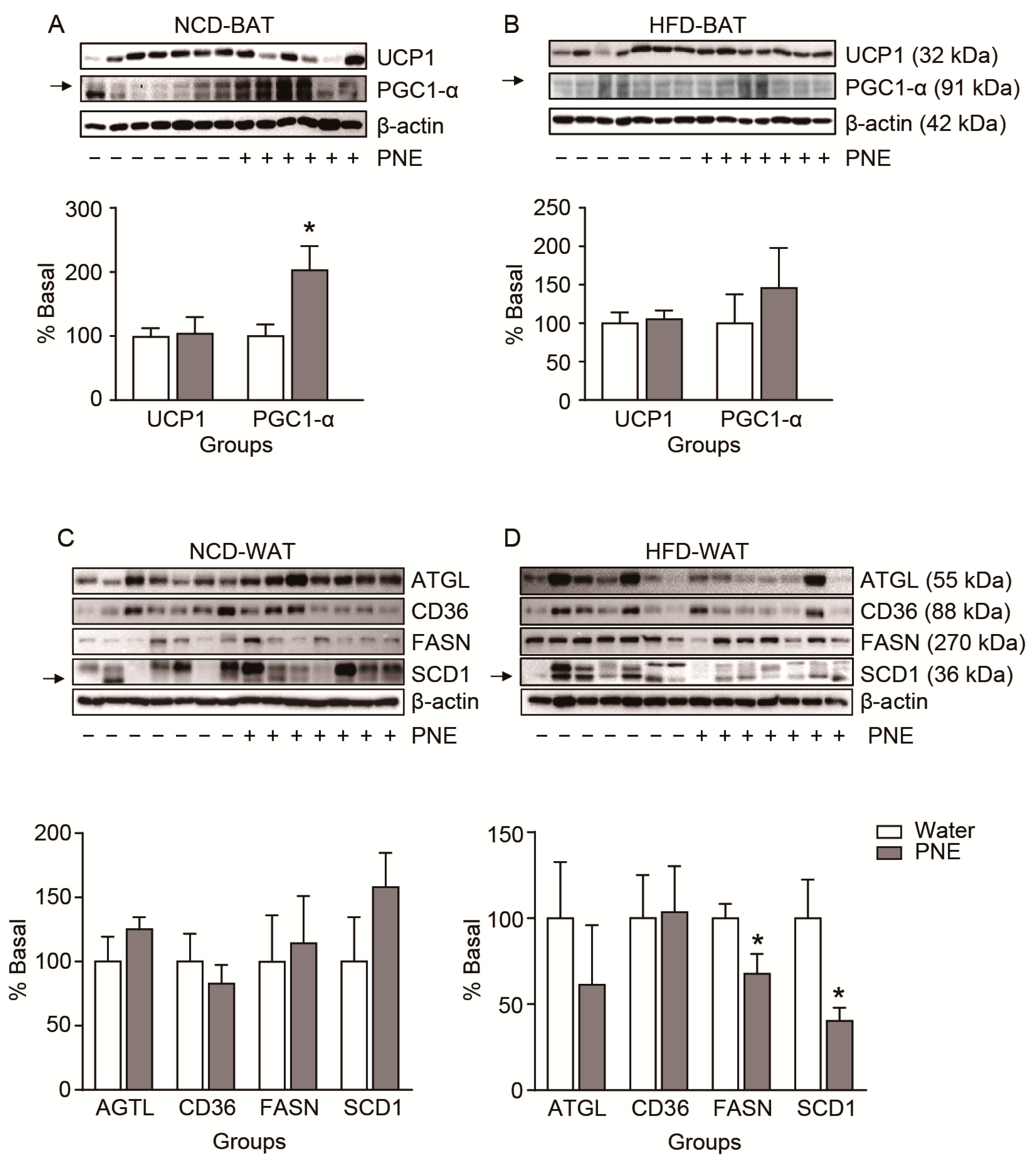
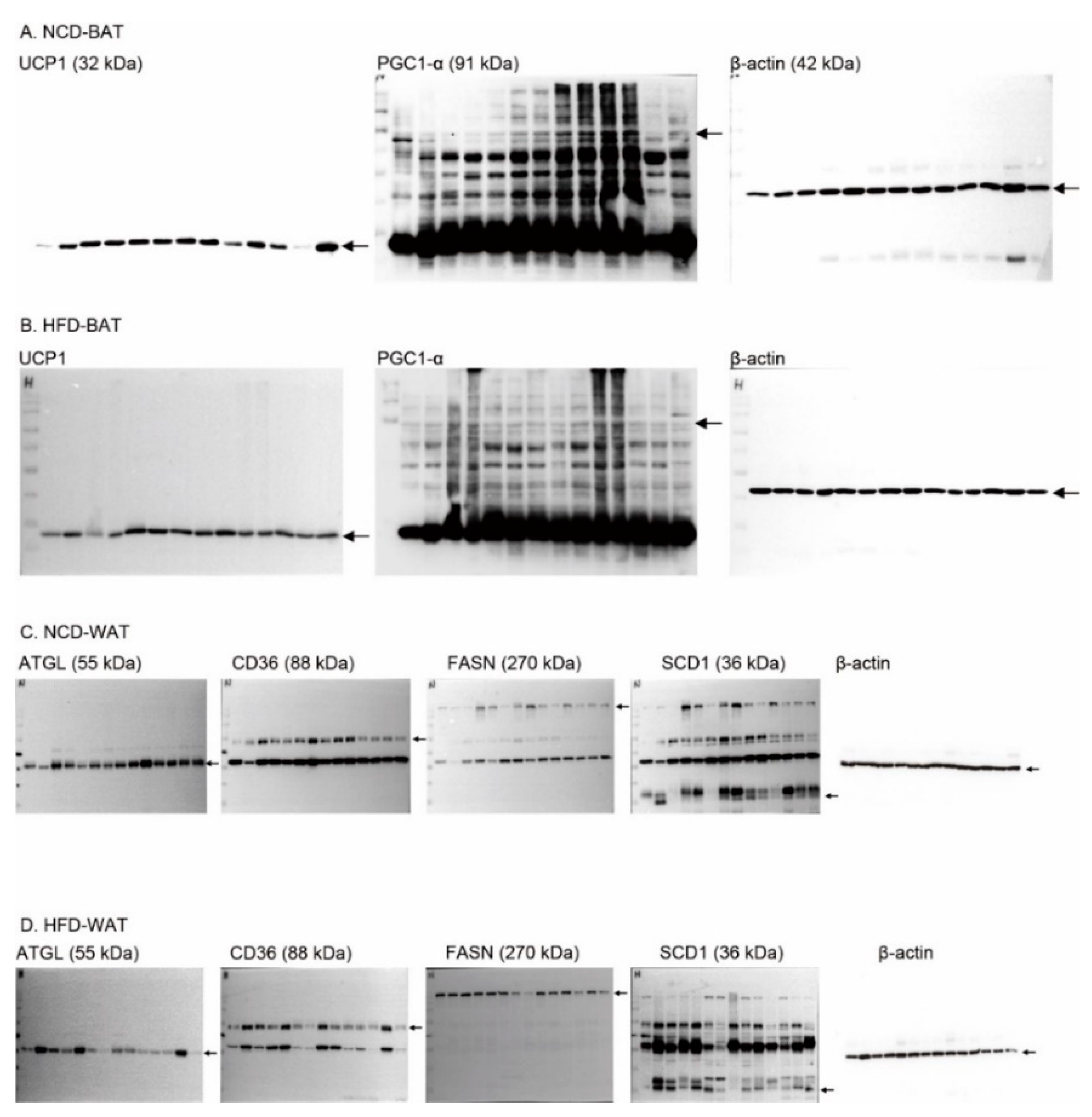
2.6. Body Condition-Dependent Regulation of Adipose Tissue Metabolism by PNE
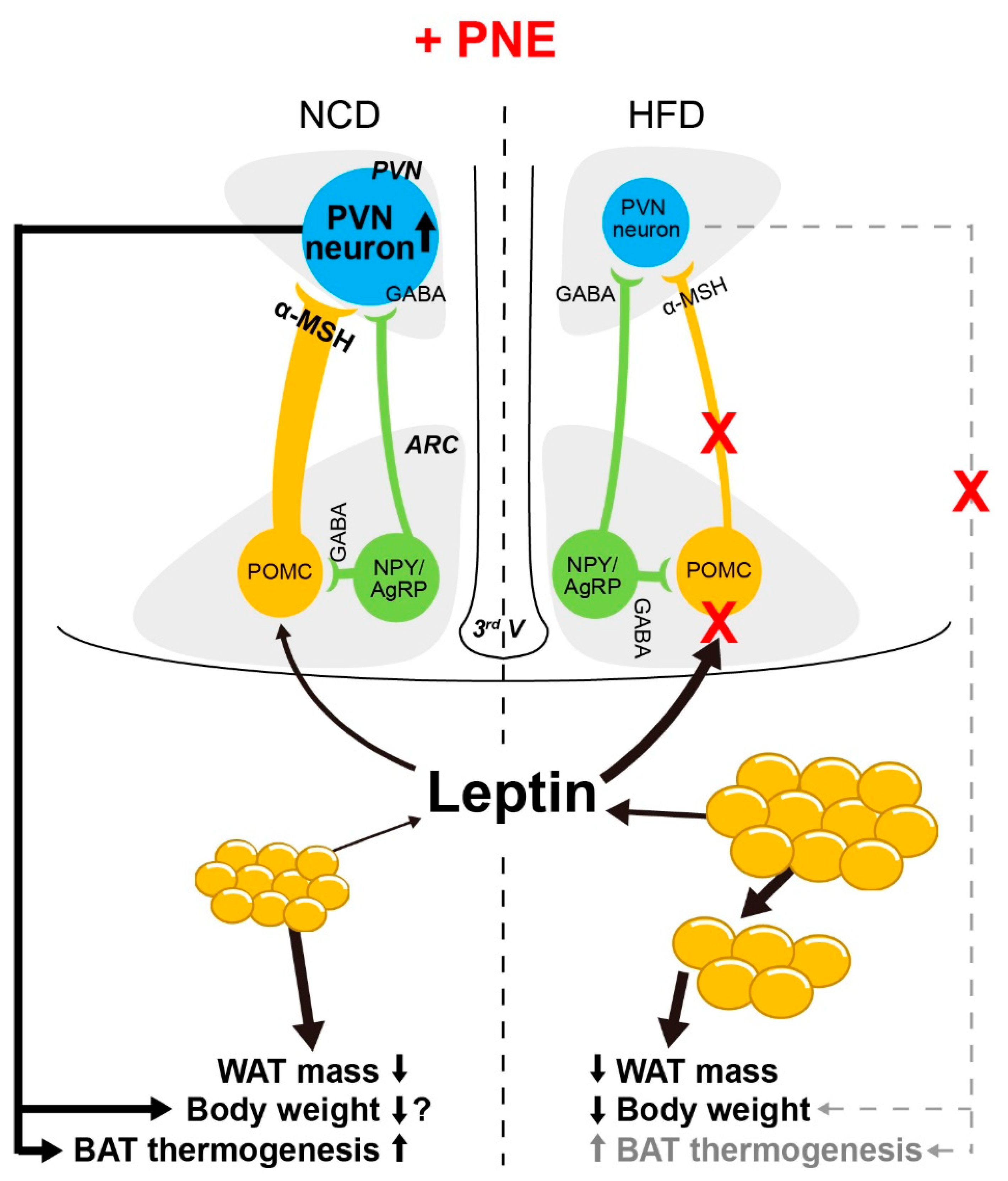
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Preparation of Pine Needle Extract
4.3. Measurement of Body Weight, Food Intake, and Body Mass Composition
4.4. Slice Preparation and Electrophysiological Recordings
4.5. Immunofluorescence Staining
4.6. IPGTT
4.7. Blood Plasma Leptin Assay
4.8. Western Blot
4.9. Statistics
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

References
- Romieu, I.; Dossus, L.; Barquera, S.; Blottière, H.M.; Franks, P.W.; Gunter, M.; Hwalla, N.; Hursting, S.D.; Leitzmann, M.; Margetts, B.; et al. Energy balance and obesity: What are the main drivers? Cancer Causes Control 2017, 28, 247–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lumeng, C.N.; Saltiel, A.R. Inflammatory links between obesity and metabolic disease. J. Clin. Investig. 2011, 121, 2111–2117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- James, O.; Holly, R.; John, C. Energy Balance and Obesity. Circulation 2012, 126, 126–132. [Google Scholar]
- Dietrich, M.O.; Horvath, T.L. Hypothalamic control of energy balance: Insights into the role of synaptic plasticity. Trends Neurosci. 2013, 36, 65–73. [Google Scholar] [CrossRef]
- Waterson, M.J.; Horvath, T.L. Neuronal Regulation of Energy Homeostasis: Beyond the Hypothalamus and Feeding. Cell Metab. 2015, 22, 962–970. [Google Scholar] [CrossRef] [Green Version]
- Harrold, J.A. Hypothalamic control of energy balance. Curr. Drug Targets 2004, 5, 207–219. [Google Scholar] [CrossRef]
- Timper, K.; Brüning, J.C. Hypothalamic circuits regulating appetite and energy homeostasis: Pathways to obesity. Dis. Model Mech. 2017, 10, 679–689. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Lin, Y.C.; Kuo, T.W.; Knight, Z.A. Sensory detection of food rapidly modulates arcuate feeding circuits. Cell 2015, 160, 829–841. [Google Scholar] [CrossRef] [Green Version]
- Belgardt, B.F.; Okamura, T.; Brüning, J.C. Hormone and glucose signalling in POMC and AgRP neurons. J. Physiol. 2009, 587, 5305–5314. [Google Scholar] [CrossRef]
- Jobst, E.E.; Enriori, P.J.; Cowley, M.A. The electrophysiology of feeding circuits. Trends Endocrinol. Metab. 2004, 15, 488–499. [Google Scholar] [CrossRef]
- Kim, G.W.; Lin, J.E.; Valentino, M.A.; Colon-Gonzalez, F.; Waldman, S.A. Regulation of Appetite to Treat Obesity. Expert Rev. Clin. Pharmacol. 2011, 4, 243–259. [Google Scholar] [CrossRef] [Green Version]
- Fenselau, H.; Campbell, J.N.; Verstegen, A.M.; Madara, J.C.; Xu, J.; Shah, B.P.; Resch, J.M.; Yang, Z.; Mandelblat-Cerf, Y.; Livneh, Y.; et al. A rapidly acting glutamatergic ARC → PVH satiety circuit postsynaptically regulated by α-MSH. Nat. Neurosci. 2017, 20, 42–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baldini, G.; Phelan, K.D. The melanocortin pathway and control of appetite–progress and therapeutic implications. J. Endocrinol. 2019, 241, R1–R33. [Google Scholar] [CrossRef] [PubMed]
- Millington, G.W. The role of proopiomelanocortin (POMC) neurones in feeding behaviour. Nutr. Metab. (Lond.) 2007, 4, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pritchard, L.E.; Turnbull, A.V.; White, A. Pro–opiomelanocortin processing in the hypothalamus: Impact on melanocortin signaling and obesity. J. Endocrinol. 2002, 172, 411–421. [Google Scholar] [CrossRef] [PubMed]
- Caron, A.; Lee, S.; Elmquist, J.K.; Gautron, L. Leptin and brain-adipose crosstalks. Nat. Rev. Neurosci. 2018, 19, 153–165. [Google Scholar] [CrossRef]
- Zhang, W.; Bi, S. Hypothalamic Regulation of Brown Adipose Tissue Thermogenesis and Energy Homeostasis. Front. Endocrinol. (Lausanne) 2015, 6, 136. [Google Scholar] [CrossRef]
- Zhang, Y.; Rodrigues, E.; Li, G.; Gao, Y.; King, M.; Carter, C.S.; Tumer, N.; Cheng, K.Y.; Scarpace, P.J. Simultaneous POMC gene transfer to hypothalamus and brainstem increases physical activity, lipolysis and reduces adult-onset obesity. Eur. J. Neurosci. 2011, 33, 1541–1550. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, K.C.D.C.; Pereira, R.M.; de Campos, T.D.P.; de Moura, R.F.; da Silva, A.S.R.; Cintra, D.E.; Ropelle, E.R.; Pauli, J.R.; de Araújo, M.B.; de Moura, L.P. The Role of Physical Exercise to Improve the Browning of White Adipose Tissue via POMC Neurons. Front. Cell. Neurosci. 2018, 12, 88. [Google Scholar] [CrossRef]
- Mejido, D.C.P.; Peny, J.A.; Vieira, M.N.N.; Ferreira, S.T.; De Felice, F.G. Insulin and leptin as potential cognitive enhancers in metabolic disorders and Alzheimer’s disease. Neuropharmacology 2020, 171, 108115. [Google Scholar] [CrossRef]
- Picone, P.; Di Carlo, M.; Nuzzo, D. Obesity and Alzheimer disease: Molecular bases. Eur. J. Neurosci. 2020. online ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Seo, H.; Lee, N.H.; Ryu, S. Antioxidant and antiapoptotic effects of pine needle powder ingestion and endurance training in high cholesterol-fed rats. J. Exerc. Nutr. Biochem. 2014, 18, 301–309. [Google Scholar] [CrossRef] [Green Version]
- Park, G.Y.; Paudyal, D.; Hwang, I.D.; Tripathi, G.; Yang, Y.K.; Cheong, H.S. Production of fermented needle extracts from red pine and their Functional characterization. Biotechnol. Bioprocess. Eng. 2008, 13, 256–261. [Google Scholar] [CrossRef]
- Li, H.; Wang, Z.; Xu, Y.; Sun, G. Pine polyphenols from Pinus koraiensis prevent injuries induced by gamma radiation in mice. PeerJ 2016, 4, e1870. [Google Scholar] [CrossRef] [PubMed]
- Proshkina, E.; Plyusnin, S.; Babak, T.; Lashmanova, E.; Maganova, F.; Koval, L.; Platonova, E.; Shaposhnikov, M.; Moskalev, A. Terpenoids as Potential Geroprotectors. Antioxidants 2020, 9, 529. [Google Scholar] [CrossRef]
- Toda, C.; Santoro, A.; Kim, J.D.; Diano, S. POMC Neurons: From Birth to Death. Annu. Rev. Physiol. 2017, 79, 209–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dodd, G.T.; Michael, N.J.; Lee-Young, R.S.; Mangiafico, S.P.; Pryor, J.T.; Munder, A.C.; Simonds, S.E.; Bringing, J.C.; Zhang, Z.Y.; Cowley, M.A.; et al. Insulin regulates POMC neuronal plasticity to control glucose metabolism. Elife 2018, 7, e38704. [Google Scholar] [CrossRef]
- Lee, D.K.; Jeong, J.H.; Chun, S.K.; Chua, S.J.; Jo, Y.H. Interplay between glucose and leptin signaling determines the strength of GABAergic synapses at POMC neurons. Nat. Commun. 2015, 6, 6618. [Google Scholar] [CrossRef] [Green Version]
- Parton, L.E.; Ye, C.P.; Coppari, R.; Enriori, P.J.; Choi, B.; Zhang, C.Y.; Xu, C.; Vianna, C.R.; Balthasar, N.; Lee, C.E.; et al. Glucose sensing by POMC neurons regulates glucose homeostasis and is impaired in obesity. Nature 2007, 449, 228–232. [Google Scholar] [CrossRef]
- Park, G.Y.; Paudyal, D.; Park, Y.M.; Lee, C.S.; Hwang, I.D.; Tripathi, G.R.; Cheong, H.S. Effects of Pine Needle Extracts on Plasma Cholesterol, Fibrinolysis and Gastrointestinal Motility. Biotechnol. Bioprocess. Eng. 2008, 13, 262. [Google Scholar] [CrossRef]
- Jeon, J.R.; Kim, J.Y. Effects of pine needle extract on differentiation of 3T3-L1 preadipocytes and obesity in high-fat diet fed rats. Biol. Pharm. Bull. 2006, 29, 2111–2115. [Google Scholar] [CrossRef] [Green Version]
- Sohn, J.W.; Elmquist, J.K.; Williams, K.W. Neuronal circuits that regulate feeding behavior and metabolism. Trends Neurosci. 2013, 36, 504–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarvestani, F.S.; Tamadon, A.; Hematzadeh, A.; Jahanara, M.; Shirazi, M.R.; Moghadam, A.; Niazi, A.; Moghiminasr, R. Expression of melanocortin-4 receptor and agouti-related peptide mRNAs in arcuate nucleus during long term malnutrition of female ovariectomized rats. Iran. J. Basic Med. Sci. 2015, 18, 104–107. [Google Scholar]
- Schwartz, M.W. Neuronal pathways regulating food intake and body adiposity. Ann. Endocrinol. (Paris) 2002, 63, 117–120. [Google Scholar] [PubMed]
- Lanfray, D.; Richard, D. Emerging Signaling Pathway in Arcuate Feeding-Related Neurons: Role of the Acbd7. Front. Neurosci. 2017, 11, 328. [Google Scholar] [CrossRef]
- Smith, M.A.; Hisadome, K.; Al-Qassab, H.; Heffron, H.; Withers, D.J.; Ashford, M.L. Melanocortins and agouti-related protein modulate the excitability of two arcuate nucleus neuron populations by alteration of resting potassium conductances. J. Physiol. 2007, 578, 425–438. [Google Scholar] [CrossRef] [Green Version]
- Anderson, E.J.; Çakir, I.; Carrington, S.J.; Cone, R.D.; Ghamari-Langroudi, M.; Gillyard, T.; Gimenez, L.E.; Litt, M.J. 60 YEARS OF POMC: Regulation of feeding and energy homeostasis by α-MSH. J. Mol. Endocrinol. 2016, 56, T157–T174. [Google Scholar] [CrossRef] [Green Version]
- Labbé, S.M.; Caron, A.; Lanfray, D.; Monge-Rofarello, B.; Bartness, T.J.; Richard, D. Hypothalamic control of brown adipose tissue thermogenesis. Front. Syst. Neurosci. 2015, 9, 150. [Google Scholar] [CrossRef]
- Contreras, C.; Gonzalez, F.; Fernø, J.; Diéguez, C.; Rahmouni, K.; Nogueiras, R.; López, M. The brain and brown fat. Ann. Med. 2015, 47, 150–168. [Google Scholar] [CrossRef] [Green Version]
- Butler, A.A.; Cone, R.D. The melanocortin receptors: Lessons from knockout models. Neuropeptides 2002, 36, 77–84. [Google Scholar] [CrossRef]
- Won, S.B.; Jung, G.Y.; Kim, J.H.; Chung, Y.S.; Hong, E.K.; Kwon, Y.H. Protective Effect of Pinus koraiensis Needle Water Extract Against Oxidative Stress in HepG2 Cells and Obese Mice. J. Med. Food 2013, 16, 569–576. [Google Scholar] [CrossRef] [Green Version]
- Boccellino, M.; D’Angelo, S. Anti-Obesity Effects of Polyphenol Intake: Current Status and Future Possibilities. Int. J. Mol. Sci. 2020, 21, 5642. [Google Scholar] [CrossRef]
- Samodien, E.; Johnson, R.; Pheiffer, C.; Mabasa, L.; Erasmus, M.; Louw, J.; Chellan, N. Diet-induced hypothalamic dysfunction and metabolic disease, and the therapeutic potential of polyphenols. Mol. Metab. 2019, 27, 1–10. [Google Scholar] [CrossRef]
- Ibars, M.; Ardid-Ruiz, A.; Suárez, M.; Muguerza, B.; Bladé, C.; Aragonès, G. Proanthocyanidins potentiate hypothalamic leptin / STAT3 signaling and Pomc gene expression in rats with diet-induced obesity. Int. J. Obes. (Lond.) 2017, 41, 129–136. [Google Scholar] [CrossRef] [Green Version]
- Siegrist-Kaiser, C.A.; Pauli, V.; Juge-Aubry, C.E.; Boss, O.; Pernin, A.; Chin, W.W.; Cusin, I.; Rohner-Jeanrenaud, F.; Burger, A.G.; Zapf, J.; et al. Direct effects of leptin on brown and white adipose tissue. J. Clin. Investig. 1997, 100, 2858–2864. [Google Scholar] [CrossRef] [Green Version]
- Wagoner, B.; Hausman, D.B.; Harris, R.B. Direct and indirect effects of leptin on preadipocyte proliferation and differentiation. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2006, 290, R1557–R1564. [Google Scholar] [CrossRef] [Green Version]
- Atawia, R.T.; Bunch, K.L.; Toque, H.A.; Caldwell, R.B.; Caldwell, R.W. Mechanisms of obesity-induced metabolic and vascular dysfunctions. Front. Biosci. (Landmark Ed.) 2019, 24, 890–934. [Google Scholar]
- Ahn, S.M.; Choe, E.S. Activation of group I metabotropic glutamate receptors increases serine phosphorylation of GluR1 alpha-amino-3-hydroxy-5-methylisoxazole-4-propionic acid receptors in the rat dorsal striatum. J. Pharmacol. Exp. Ther. 2009, 329, 1117–1126. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.M.; Ahn, S.M.; Go, B.S.; Wang, J.Q.; Choe, E.S. Alterations in AMPA receptor phosphorylation in the rat striatum following acute and repeated cocaine administration. Neuroscience 2009, 163, 618–626. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, E.A.; Yang, J.-H.; Byeon, E.-H.; Kim, W.; Kang, D.; Han, J.; Hong, S.-G.; Kim, D.-R.; Park, S.-J.; Huh, J.-W.; et al. Anti-Obesity Effect of Pine Needle Extract on High-Fat Diet-Induced Obese Mice. Plants 2021, 10, 837. https://doi.org/10.3390/plants10050837
Kim EA, Yang J-H, Byeon E-H, Kim W, Kang D, Han J, Hong S-G, Kim D-R, Park S-J, Huh J-W, et al. Anti-Obesity Effect of Pine Needle Extract on High-Fat Diet-Induced Obese Mice. Plants. 2021; 10(5):837. https://doi.org/10.3390/plants10050837
Chicago/Turabian StyleKim, Eun A., Ju-Hwan Yang, Eun-Hye Byeon, Woong Kim, Dawon Kang, Jaehee Han, Seong-Geun Hong, Deok-Ryong Kim, Sang-Je Park, Jae-Won Huh, and et al. 2021. "Anti-Obesity Effect of Pine Needle Extract on High-Fat Diet-Induced Obese Mice" Plants 10, no. 5: 837. https://doi.org/10.3390/plants10050837