
Molecular Mechanisms of Anti-Melanogenic Gedunin Derived from Neem Tree (Azadirachta indica) Using B16F10 Mouse Melanoma Cells and Early-Stage Zebrafish

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Chemicals and Reagents
2.2. Cell Culture
2.3. Cell Viability Assay
2.4. Determination of Melanin Contents
2.5. Intracellular Tyrosinase Activity Assay
2.6. RNA Isolation and qRT-PCR
2.7. Immunoblot Analysis
2.8. Zebrafish Embryo Test
2.9. Statistical Analysis
3. Results
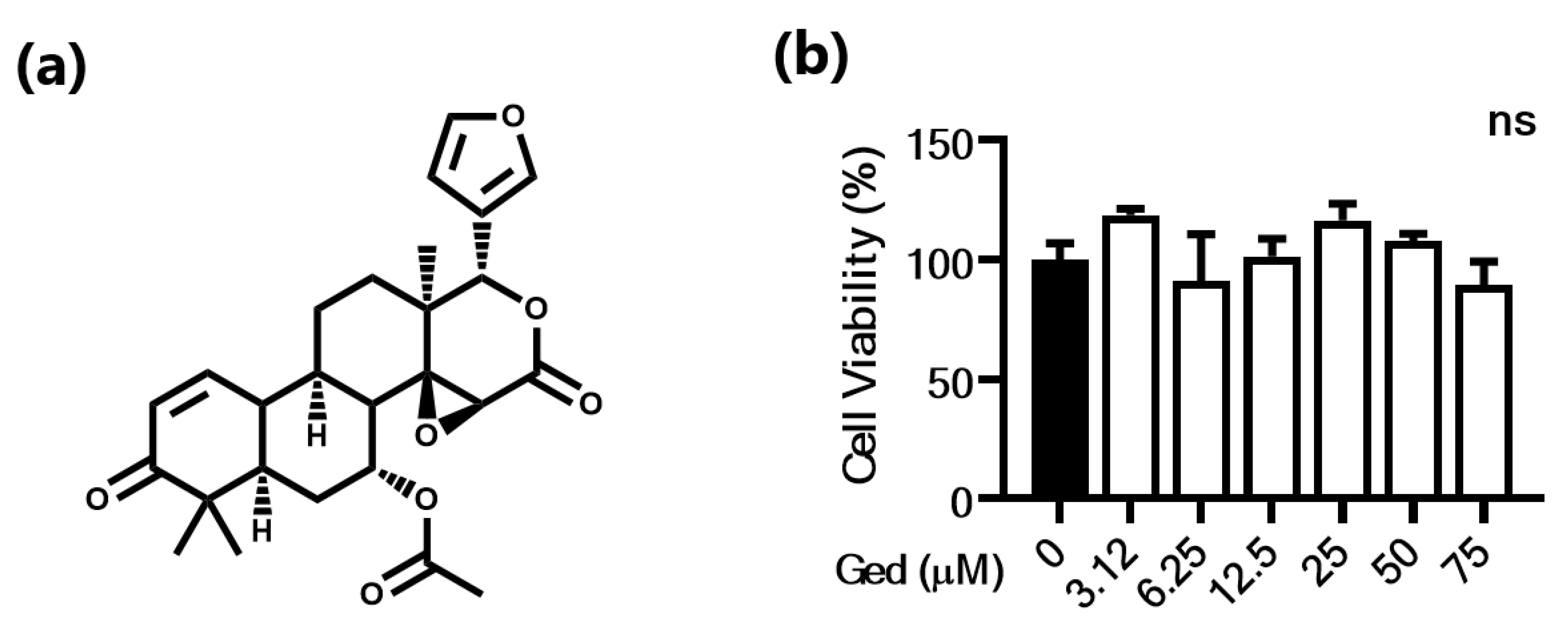
3.1. Cytotoxic Effect of Piperlongumine in B16F10 Cells
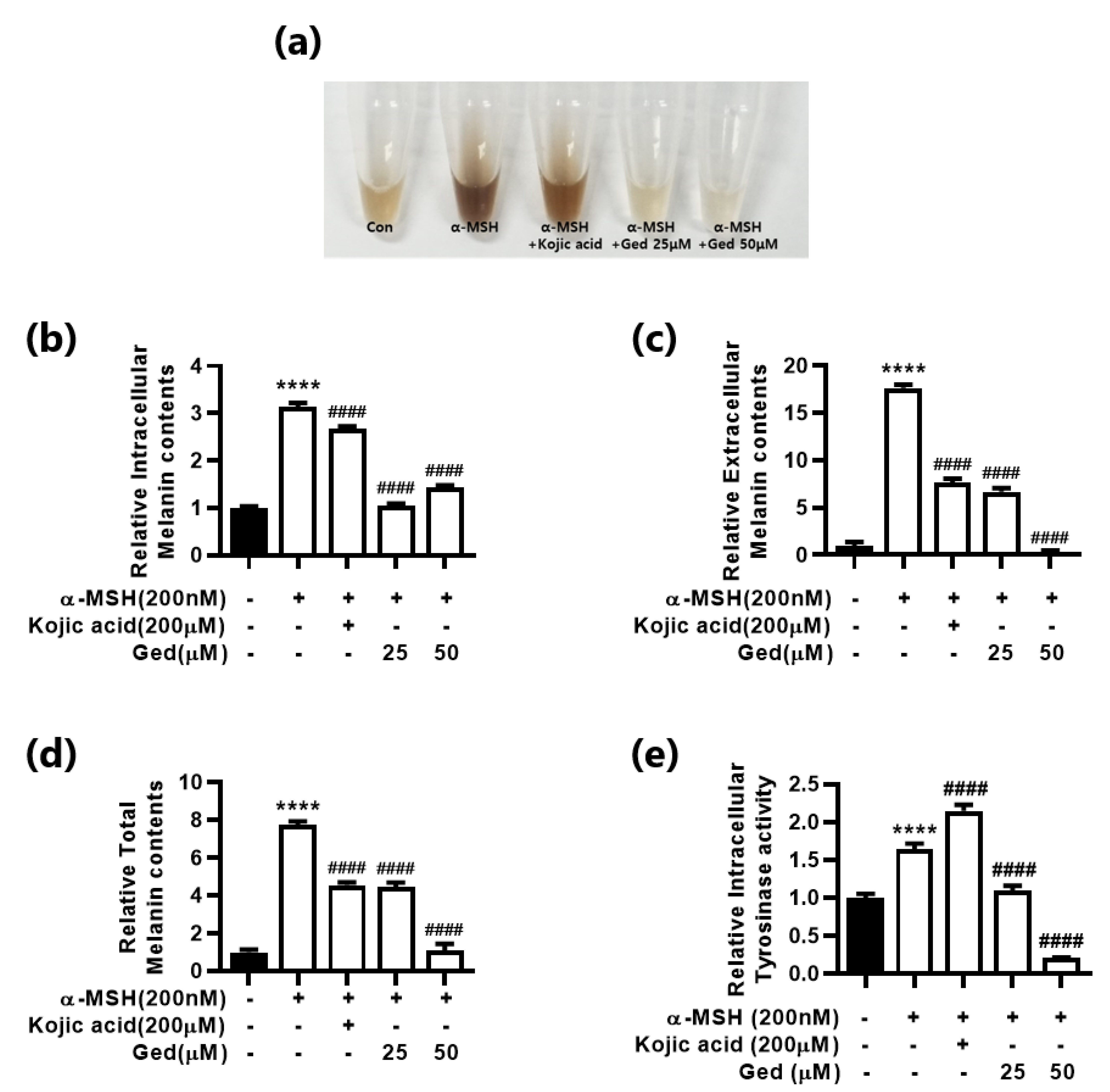
3.2. Gedunin Inhibits Melanin Production and Intracellular Tyrosinase Activity in B16F10 Mouse Melanoma Cells
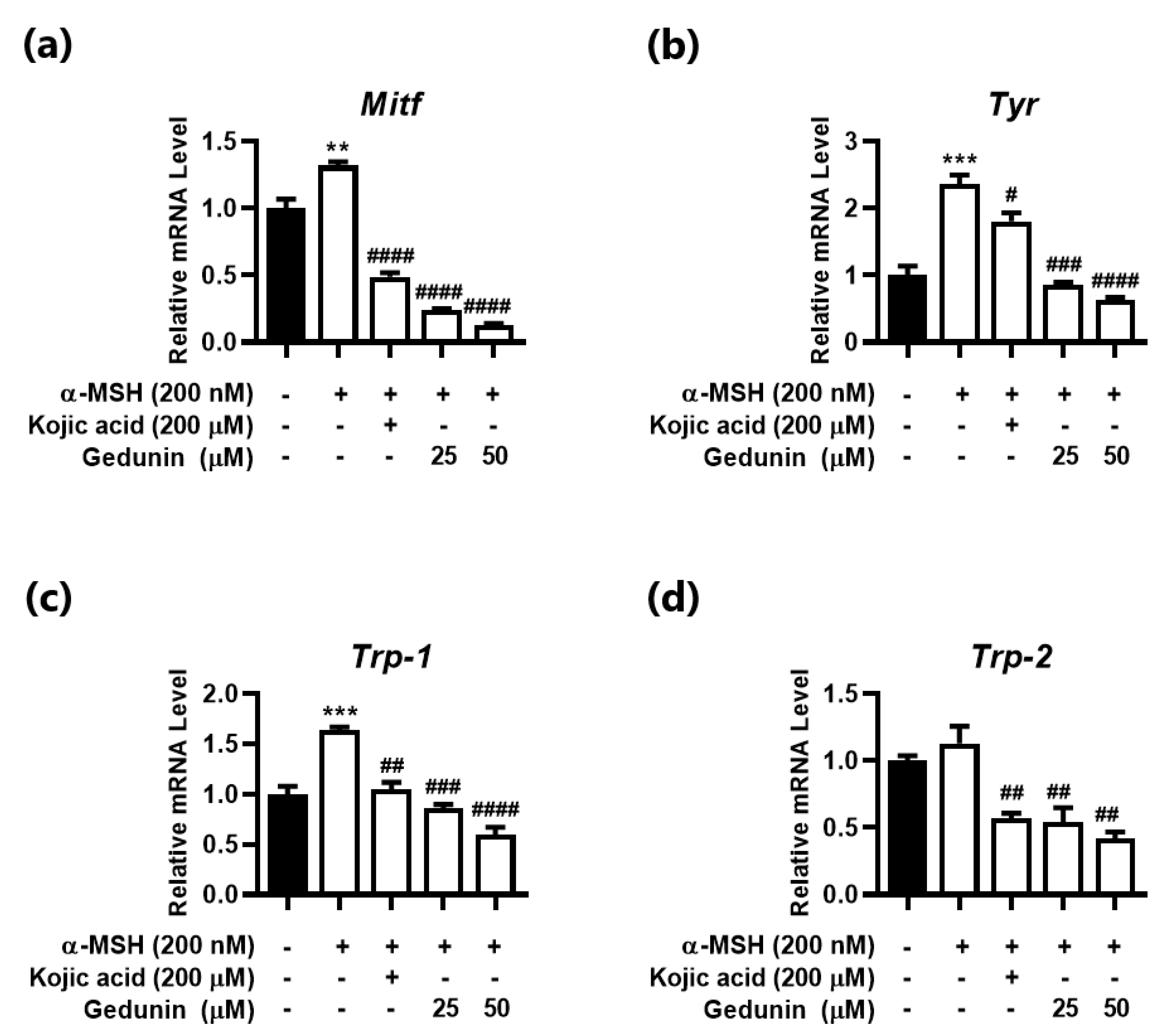
3.3. Gedunin Reduces α-MSH Induced Melanogenesis-Related Gene Expression
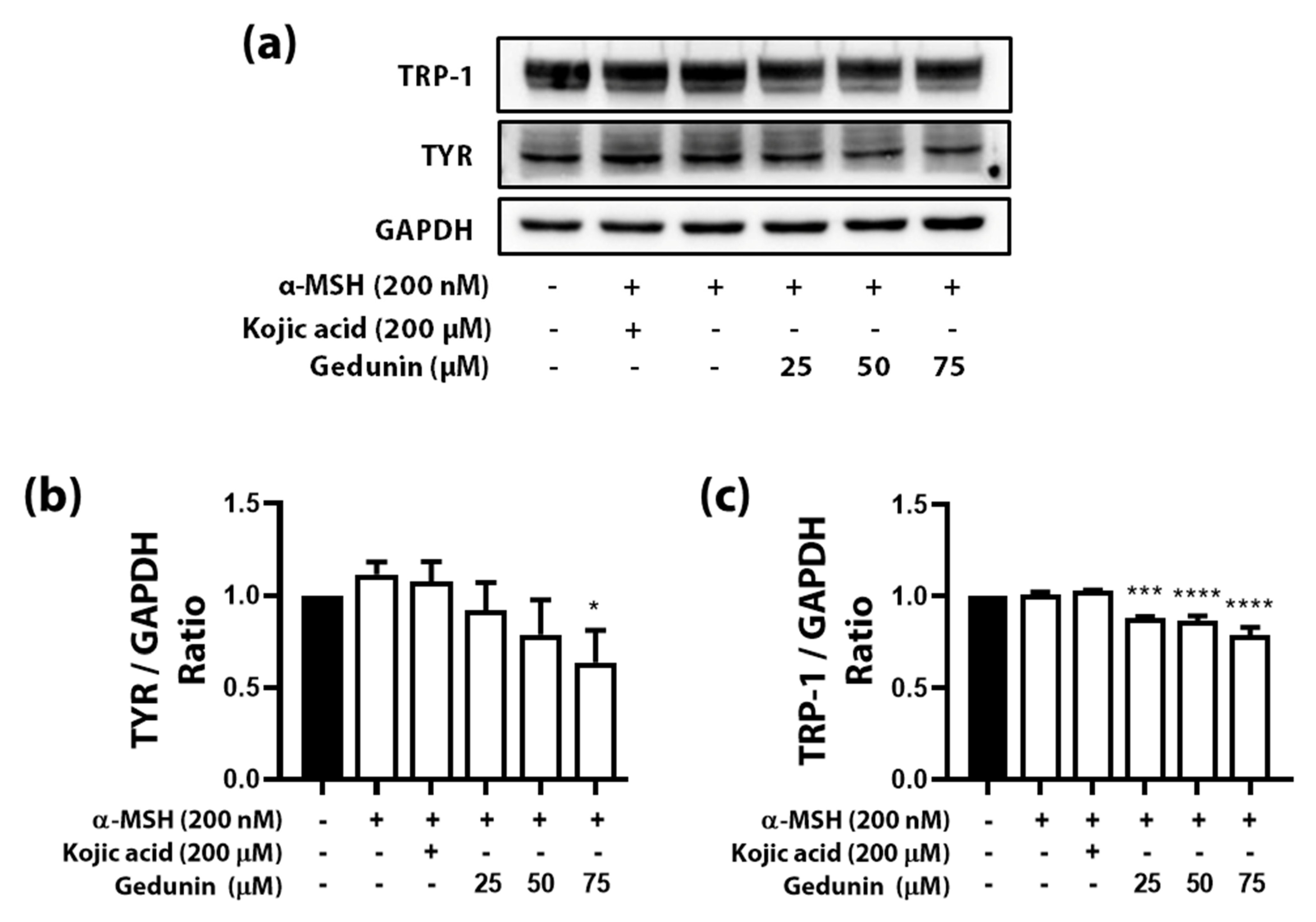
3.4. Gedunin Reduced TYR and TYR-1 Amount
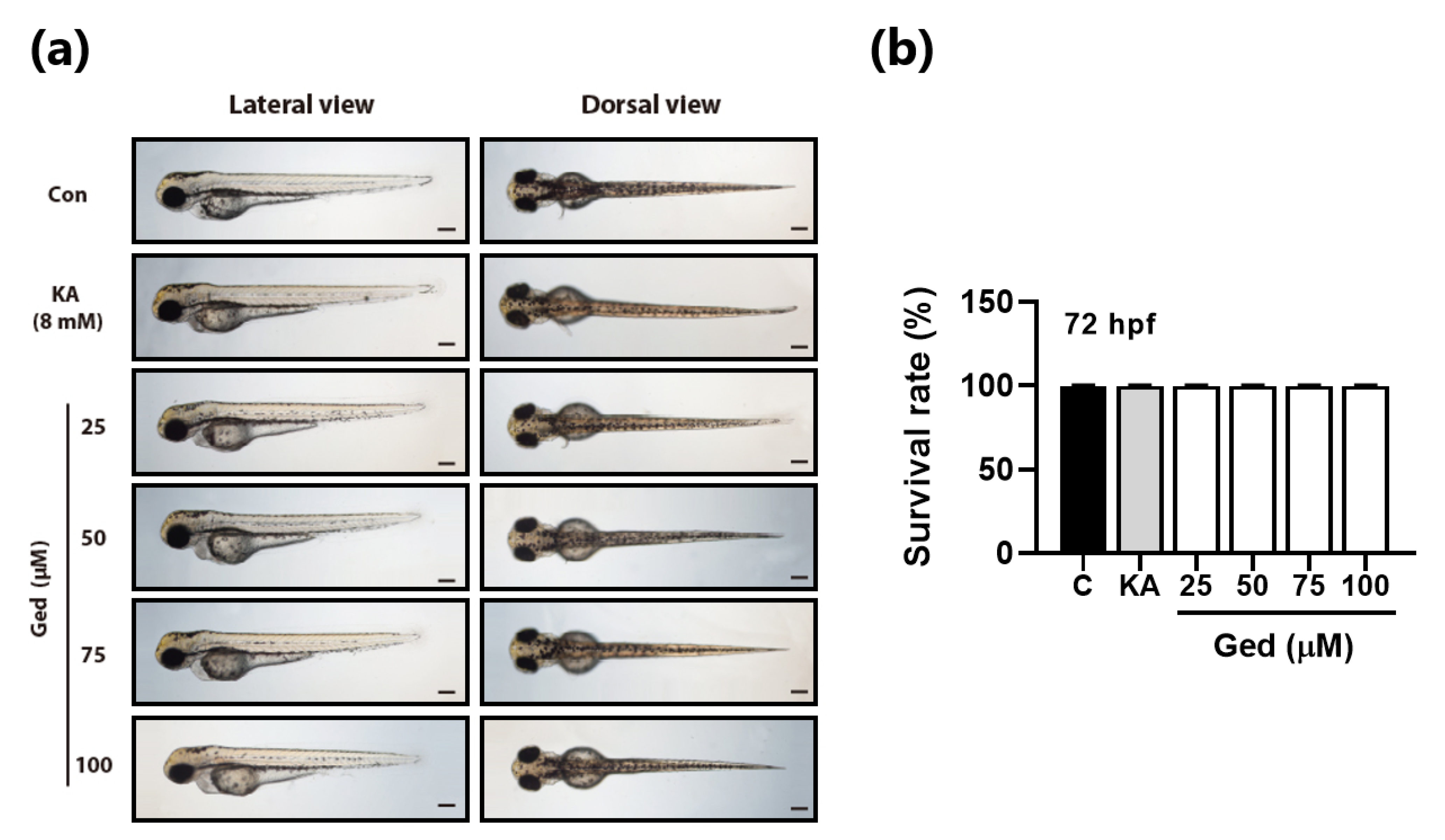
3.5. Toxicity of Gedunin against Zebrafish in Early-Stage Development
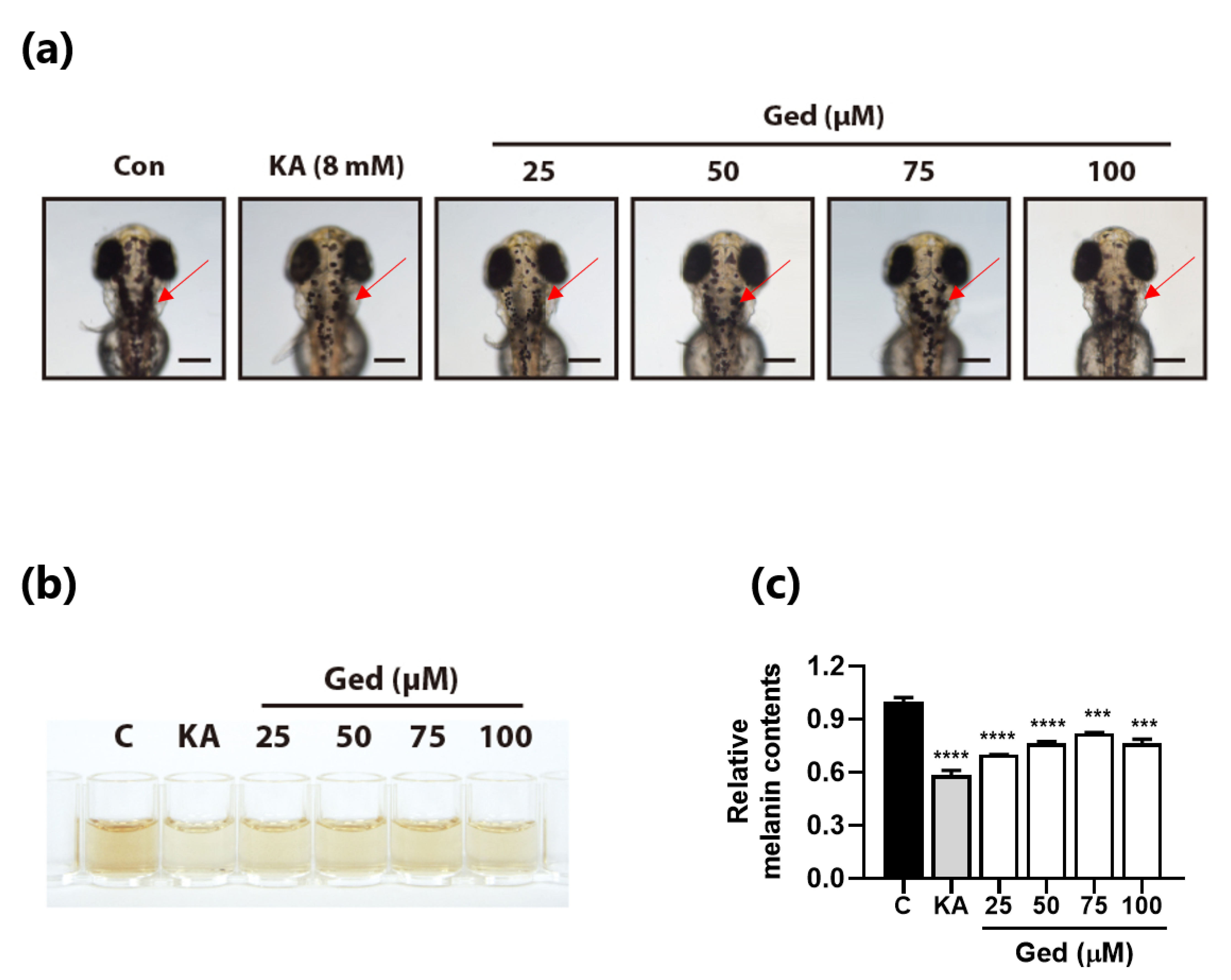
3.6. Gedunin Reduced Melanin Contents in Zebrafish Embryos
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pillaiyar, T.; Manickam, M.; Jung, S.H. Recent development of signaling pathways inhibitors of melanogenesis. Cell Signal. 2017, 40, 99–115. [Google Scholar] [CrossRef] [PubMed]
- D’Mello, S.A.; Finlay, G.J.; Baguley, B.C.; Askarian-Amiri, M.E. Signaling Pathways in Melanogenesis. Int. J. Mol. Sci. 2016, 17, 1144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolf Horrell, E.M.; Boulanger, M.C.; D’Orazio, J.A. Melanocortin 1 Receptor: Structure, Function, and Regulation. Front. Genet. 2016, 7, 95. [Google Scholar] [CrossRef] [Green Version]
- Videira, I.F.; Moura, D.F.; Magina, S. Mechanisms regulating melanogenesis. An. Bras. Dermatol. 2013, 88, 76–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vachtenheim, J.; Borovansky, J. “Transcription physiology” of pigment formation in melanocytes: Central role of MITF. Exp. Dermatol. 2010, 19, 617–627. [Google Scholar] [CrossRef] [PubMed]
- Gillbro, J.M.; Olsson, M.J. The melanogenesis and mechanisms of skin-lightening agent-existing and new approaches. Int. J. Cosmet. Sci. 2011, 33, 210–221. [Google Scholar] [CrossRef] [PubMed]
- Cortes, R.; Agulleiro, M.J.; Navarro, S.; Guillot, R.; Sanchez, E.; Cerda-Reverter, J.M. Melanocortin receptor accessory protein 2 (MRAP2) interplays with the zebrafish melanocortin 1 receptor (MC1R) but has no effect on its pharmacological profile. Gen. Comp. Endocrinol. 2014, 201, 30–36. [Google Scholar] [CrossRef]
- Cheng, M.C.; Lee, T.H.; Chu, Y.T.; Syu, L.L.; Hsu, S.J.; Cheng, C.H.; Wu, J.; Lee, C.K. Melanogenesis Inhibitors from the Rhizoma of Ligusticum Sinense in B16-F10 Melanoma Cells In Vitro and Zebrafish In Vivo. Int. J. Mol. Sci. 2018, 19, 3994. [Google Scholar] [CrossRef] [Green Version]
- Lajis, A.F.B. A Zebrafish Embryo as an Animal Model for the Treatment of Hyperpigmentation in Cosmetic Dermatology Medicine. Medicina 2018, 54, 35. [Google Scholar] [CrossRef] [Green Version]
- Agalou, A.; Thrapsianiotis, M.; Angelis, A.; Papakyriakou, A.; Skaltsounis, A.L.; Aligiannis, N.; Beis, D. Identification of Novel Melanin Synthesis Inhibitors from Crataegus pycnoloba Using an in Vivo Zebrafish Phenotypic Assay. Front. Pharmacol. 2018, 9, 265. [Google Scholar] [CrossRef]
- Choi, T.Y.; Kim, J.H.; Ko, D.H.; Kim, C.H.; Hwang, J.S.; Ahn, S.; Kim, S.Y.; Kim, C.D.; Lee, J.H.; Yoon, T.J. Zebrafish as a new model for phenotype-based screening of melanogenic regulatory compounds. Pigment. Cell Res. 2007, 20, 120–127. [Google Scholar] [CrossRef]
- Roh, E.; Yun, C.Y.; Lee, J.W.; Lee, C.; Hwang, B.Y.; Oh, S.T.; Jung, S.H.; Han, S.B.; Kim, Y. Hypopigmenting activity of bisabolangelone isolated from Angelica koreana Maxim. in alpha-melanocyte stimulating hormone-activated B16 or melan-a cells. Planta Med. 2011, 77, 248–251. [Google Scholar] [CrossRef]
- Roh, E.; Jeong, I.Y.; Shin, H.; Song, S.; Doo Kim, N.; Jung, S.H.; Tae Hong, J.; Ho Lee, S.; Han, S.B.; Kim, Y. Downregulation of melanocyte-specific facultative melanogenesis by 4-hydroxy-3-methoxycinnamaldehyde acting as a cAMP antagonist. J. Investig. Dermatol. 2014, 134, 551–553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, H.; Hong, S.D.; Roh, E.; Jung, S.H.; Cho, W.J.; Park, S.H.; Yoon, D.Y.; Ko, S.M.; Hwang, B.Y.; Hong, J.T.; et al. cAMP-dependent activation of protein kinase A as a therapeutic target of skin hyperpigmentation by diphenylmethylene hydrazinecarbothioamide. Br. J. Pharmacol. 2015, 172, 3434–3445. [Google Scholar] [CrossRef] [Green Version]
- Tharmarajah, L.; Samarakoon, S.R.; Ediriweera, M.K.; Piyathilaka, P.; Tennekoon, K.H.; Senathilake, K.S.; Rajagopalan, U.; Galhena, P.B.; Thabrew, I. In Vitro Anticancer Effect of Gedunin on Human Teratocarcinomal (NTERA-2) Cancer Stem-Like Cells. Biomed. Res. Int. 2017, 2017, 2413197. [Google Scholar] [CrossRef] [Green Version]
- Borges, P.V.; Moret, K.H.; Raghavendra, N.M.; Costa, T.E.; Monteiro, A.P.; Carneiro, A.B.; Pacheco, P.; Temerozo, J.R.; Bou-Habib, D.C.; das Gracas Henriques, M.; et al. Protective effect of gedunin on TLR-mediated inflammation by modulation of inflammasome activation and cytokine production: Evidence of a multitarget compound. Pharmacol. Res. 2017, 115, 65–77. [Google Scholar] [CrossRef] [PubMed]
- Borges, P.V.; Moret, K.H.; Maya-Monteiro, C.M.; Souza-Silva, F.; Alves, C.R.; Batista, P.R.; Caffarena, E.R.; Pacheco, P.; Henriques, M.; Penido, C. Gedunin Binds to Myeloid Differentiation Protein 2 and Impairs Lipopolysaccharide-Induced Toll-Like Receptor 4 Signaling in Macrophages. Mol. Pharmacol. 2015, 88, 949–961. [Google Scholar] [CrossRef] [Green Version]
- Braga, T.M.; Rocha, L.; Chung, T.Y.; Oliveira, R.F.; Pinho, C.; Oliveira, A.I.; Morgado, J.; Cruz, A. Biological Activities of Gedunin-A Limonoid from the Meliaceae Family. Molecules 2020, 25, 493. [Google Scholar] [CrossRef] [Green Version]
- Thomford, N.E.; Senthebane, D.A.; Rowe, A.; Munro, D.; Seele, P.; Maroyi, A.; Dzobo, K. Natural Products for Drug Discovery in the 21st Century: Innovations for Novel Drug Discovery. Int. J. Mol. Sci. 2018, 19, 1578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Endo, H.; Owada, S.; Inagaki, Y.; Shida, Y.; Tatemichi, M. Glucose starvation induces LKB1-AMPK-mediated MMP-9 expression in cancer cells. Sci. Rep. 2018, 8, 10122. [Google Scholar] [CrossRef]
- Oh, T.I.; Jung, H.J.; Lee, Y.M.; Lee, S.; Kim, G.H.; Kan, S.Y.; Kang, H.; Oh, T.; Ko, H.M.; Kwak, K.C.; et al. Zerumbone, a Tropical Ginger Sesquiterpene of Zingiber officinale Roscoe, Attenuates alpha-MSH-Induced Melanogenesis in B16F10 Cells. Int. J. Mol. Sci. 2018, 19, 3149. [Google Scholar] [CrossRef] [Green Version]
- Seo, G.Y.; Ha, Y.; Park, A.H.; Kwon, O.W.; Kim, Y.J. Leathesia difformis Extract Inhibits alpha-MSH-Induced Melanogenesis in B16F10 Cells via Down-Regulation of CREB Signaling Pathway. Int. J. Mol. Sci. 2019, 20, 536. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.D.; Hwang, S.L.; Jeon, H.J.; Jeon, Y.H.; Nedumaran, B.; Kim, K.; Lee, S.E. B-cell translocation gene 2 enhances fibroblast growth factor 21 production by inducing Kruppel-like factor 15. Sci. Rep. 2019, 9, 3730. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.; Wang, C.H.; Ok, Y.S.; Lee, S.E. Heart developmental toxicity by carbon black waste generated from oil refinery on zebrafish embryos (Danio rerio): Combined toxicity on heart function by nickel and vanadium. J. Hazard. Mater. 2019, 363, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Bertolotto, C.; Abbe, P.; Hemesath, T.J.; Bille, K.; Fisher, D.E.; Ortonne, J.P.; Ballotti, R. Microphthalmia gene product as a signal transducer in cAMP-induced differentiation of melanocytes. J. Cell Biol. 1998, 142, 827–835. [Google Scholar] [CrossRef] [Green Version]
- Eo, S.H.; Kim, S.J. Resveratrol-mediated inhibition of cyclooxygenase-2 in melanocytes suppresses melanogenesis through extracellular signal-regulated kinase 1/2 and phosphoinositide 3-kinase/Akt signalling. Eur. J. Pharmacol. 2019, 860, 172586. [Google Scholar] [CrossRef] [PubMed]
- Yasumoto, K.; Yokoyama, K.; Shibata, K.; Tomita, Y.; Shibahara, S. Microphthalmia-associated transcription factor as a regulator for melanocyte-specific transcription of the human tyrosinase gene. Mol. Cell. Biol. 1994, 14, 8058–8070. [Google Scholar] [CrossRef]
- Couteau, C.; Coiffard, L. Overview of Skin Whitening Agents: Drugs and Cosmetic Products. Cosmetics 2016, 3, 27. [Google Scholar] [CrossRef]
- Manosroi, A.; Kitdamrongtham, W.; Ishii, K.; Shinozaki, T.; Tachi, Y.; Takagi, M.; Ebina, K.; Zhang, J.; Manosroi, J.; Akihisa, R.; et al. Limonoids from Azadirachta indica var. siamensis extracts and their cytotoxic and melanogenesis-inhibitory activities. Chem. Biodivers. 2014, 11, 505–531. [Google Scholar] [CrossRef] [PubMed]
- Akihisa, T.; Takahashi, A.; Kikuchi, T.; Takagi, M.; Watanabe, K.; Fukatsu, M.; Fujita, Y.; Banno, N.; Tokuda, H.; Yasukawa, K. The melanogenesis-inhibitory, anti-inflammatory, and chemopreventive effects of limonoids in n-hexane extract of Azadirachta indica A. Juss. (neem) seeds. J. Oleo Sci. 2011, 60, 53–59. [Google Scholar] [CrossRef] [Green Version]
- Patwardhan, C.A.; Fauq, A.; Peterson, L.B.; Miller, C.; Blagg, B.S.; Chadli, A. Gedunin inactivates the co-chaperone p23 protein causing cancer cell death by apoptosis. J. Biol. Chem. 2013, 288, 7313–7325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subramani, R.; Gonzalez, E.; Nandy, S.B.; Arumugam, A.; Camacho, F.; Medel, J.; Alabi, D.; Lakshmanaswamy, R. Gedunin inhibits pancreatic cancer by altering sonic hedgehog signaling pathway. Oncotarget 2017, 8, 10891–10904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasan, A.; Haque, E.; Hameed, R.; Maier, P.N.; Irfan, S.; Kamil, M.; Nazir, A.; Mir, S.S. Hsp90 inhibitor gedunin causes apoptosis in A549 lung cancer cells by disrupting Hsp90:Beclin-1:Bcl-2 interaction and downregulating autophagy. Life Sci. 2020, 256, 118000. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeon, H.-J.; Kim, K.; Kim, C.; Kim, M.-J.; Kim, T.-O.; Lee, S.-E. Molecular Mechanisms of Anti-Melanogenic Gedunin Derived from Neem Tree (Azadirachta indica) Using B16F10 Mouse Melanoma Cells and Early-Stage Zebrafish. Plants 2021, 10, 330. https://doi.org/10.3390/plants10020330
Jeon H-J, Kim K, Kim C, Kim M-J, Kim T-O, Lee S-E. Molecular Mechanisms of Anti-Melanogenic Gedunin Derived from Neem Tree (Azadirachta indica) Using B16F10 Mouse Melanoma Cells and Early-Stage Zebrafish. Plants. 2021; 10(2):330. https://doi.org/10.3390/plants10020330
Chicago/Turabian StyleJeon, Hwang-Ju, Kyeongnam Kim, Chaeeun Kim, Myoung-Jin Kim, Tae-Oh Kim, and Sung-Eun Lee. 2021. "Molecular Mechanisms of Anti-Melanogenic Gedunin Derived from Neem Tree (Azadirachta indica) Using B16F10 Mouse Melanoma Cells and Early-Stage Zebrafish" Plants 10, no. 2: 330. https://doi.org/10.3390/plants10020330