
A Decade’s Change in Vegetation Productivity and Its Response to Climate Change over Northeast China


Abstract
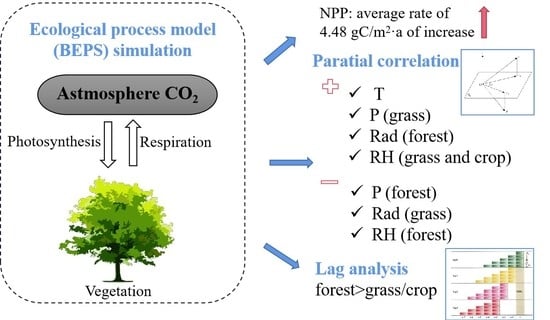
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
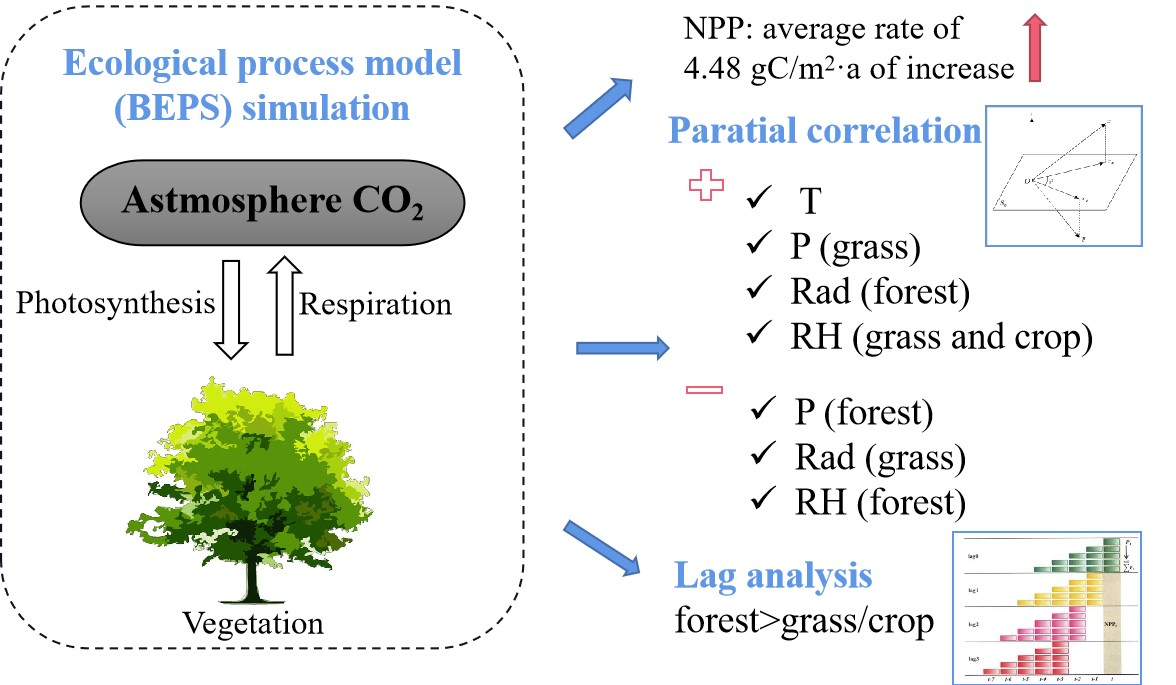
2.1. Validation of the BEPS Model
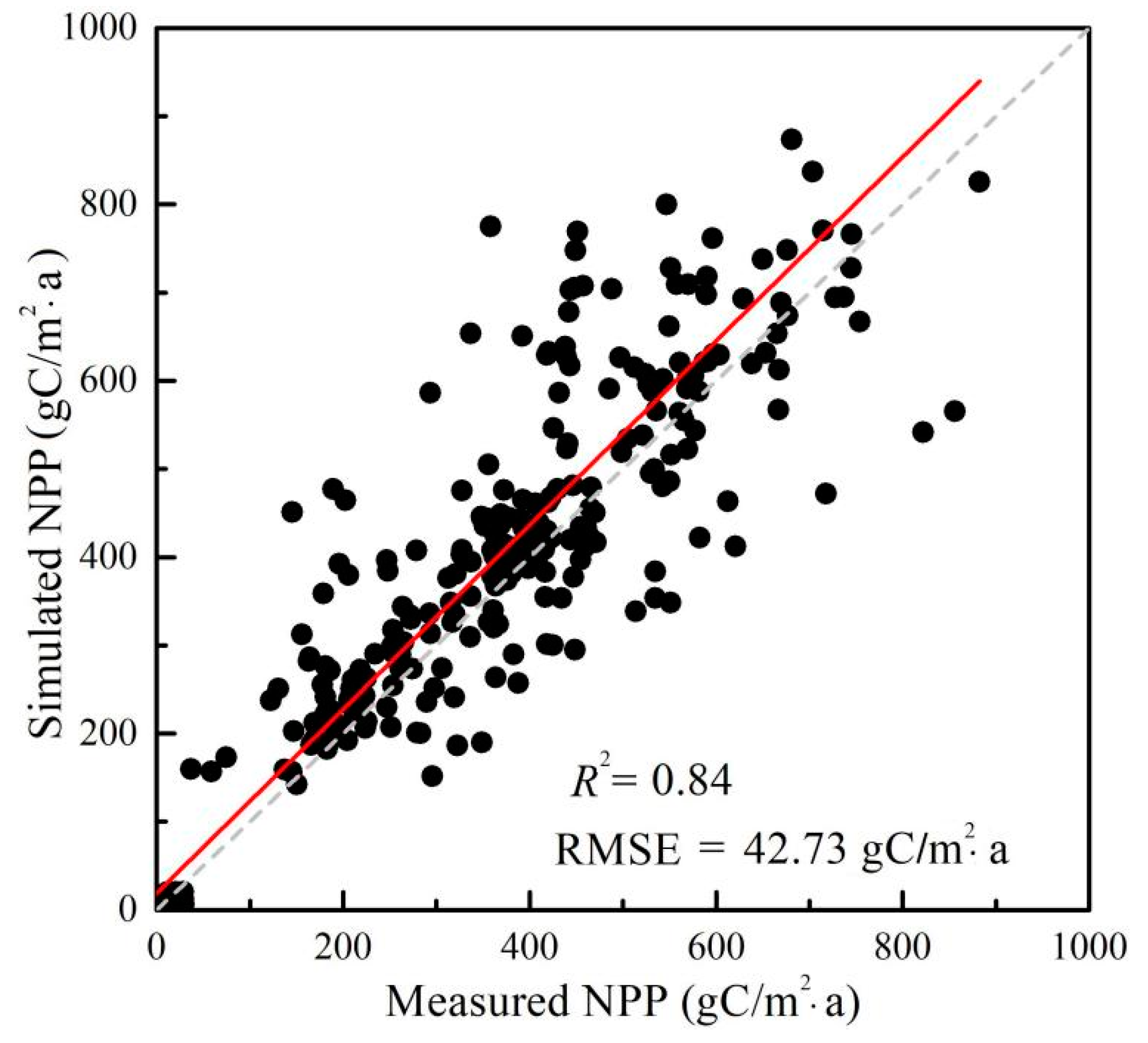
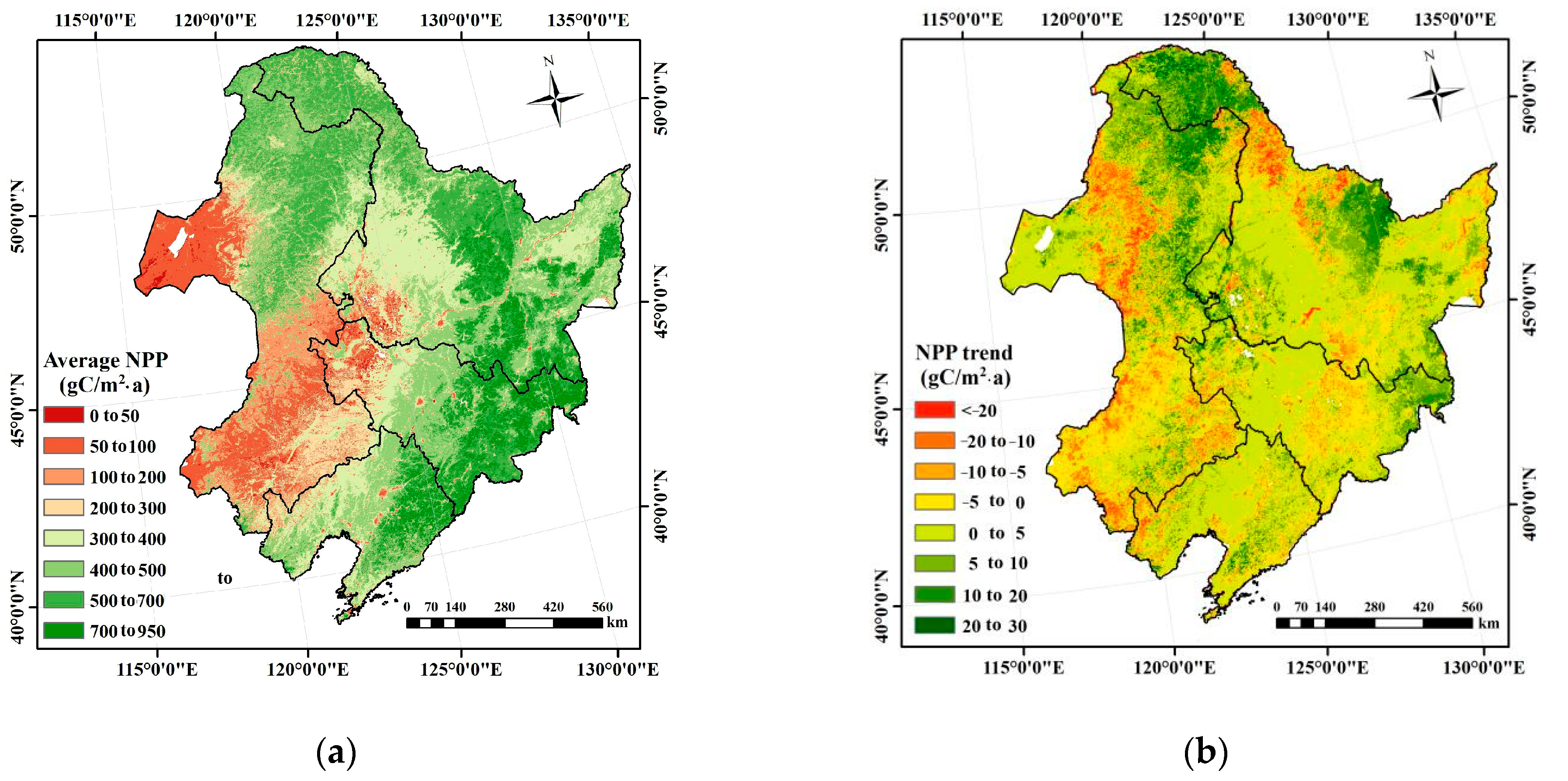
2.2. NPP Spatial Patterns and Temporal Trends
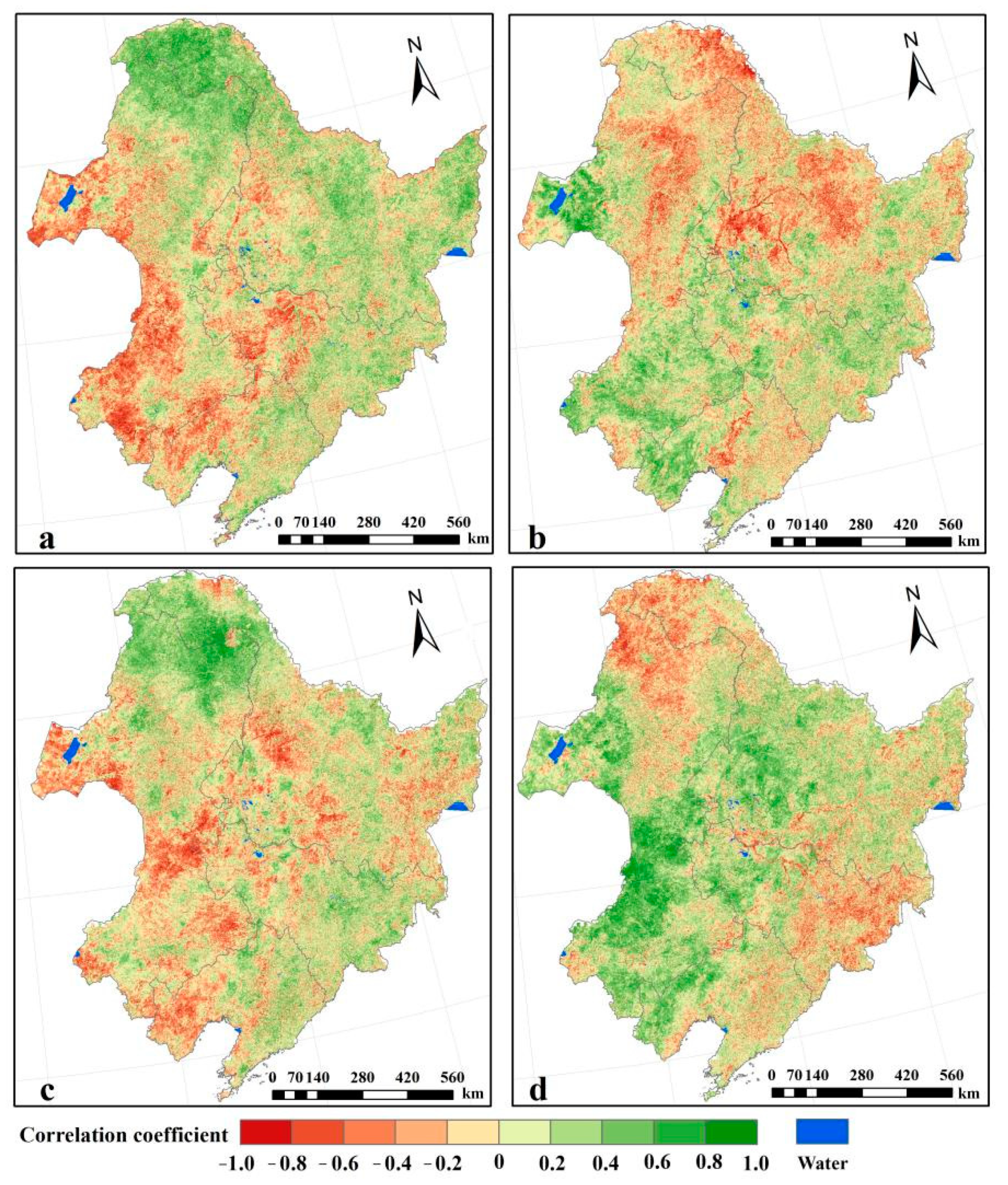
2.3. Partial Correlation Analysis between the NPP and Climatic Factors
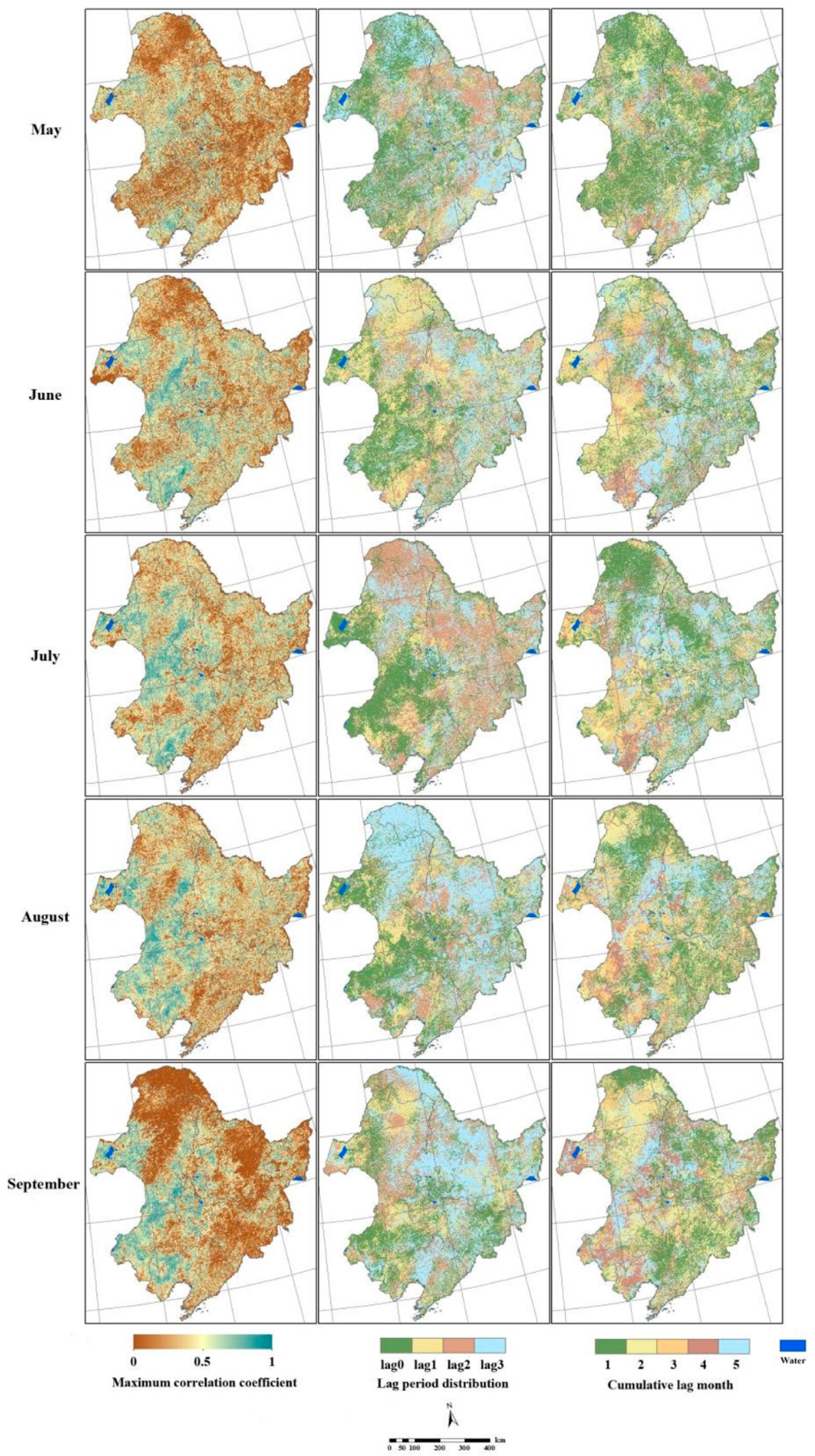
2.4. Lag Response of the NPP to Precipitation
3. Discussion
3.1. Improvement of the Regional Vegetation NPP Simulation
3.2. Response of the Vegetation NPP to Climate Change
4. Materials and Methods
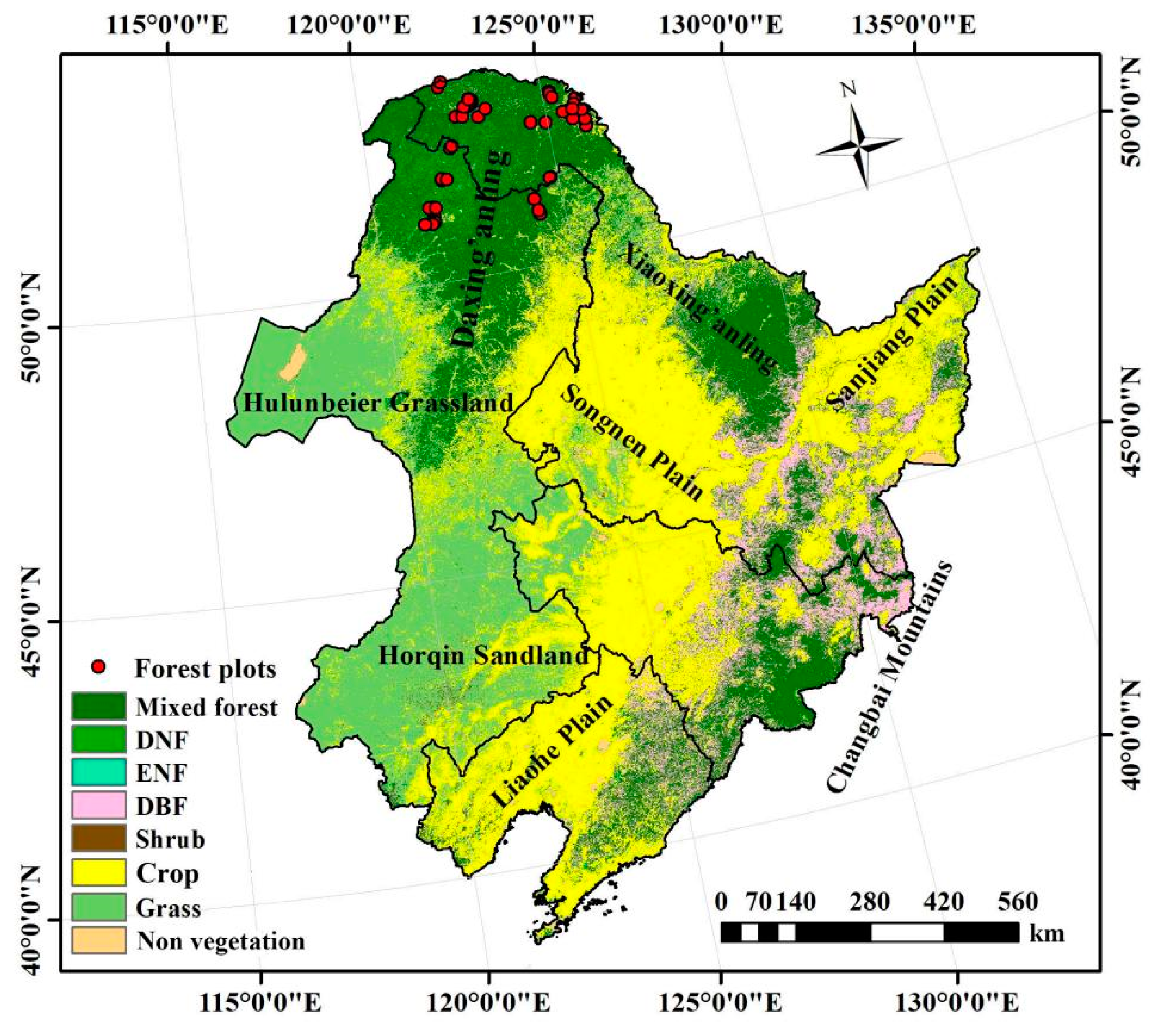
4.1. Research Area
4.2. Dataset
4.2.1. Meteorological Data
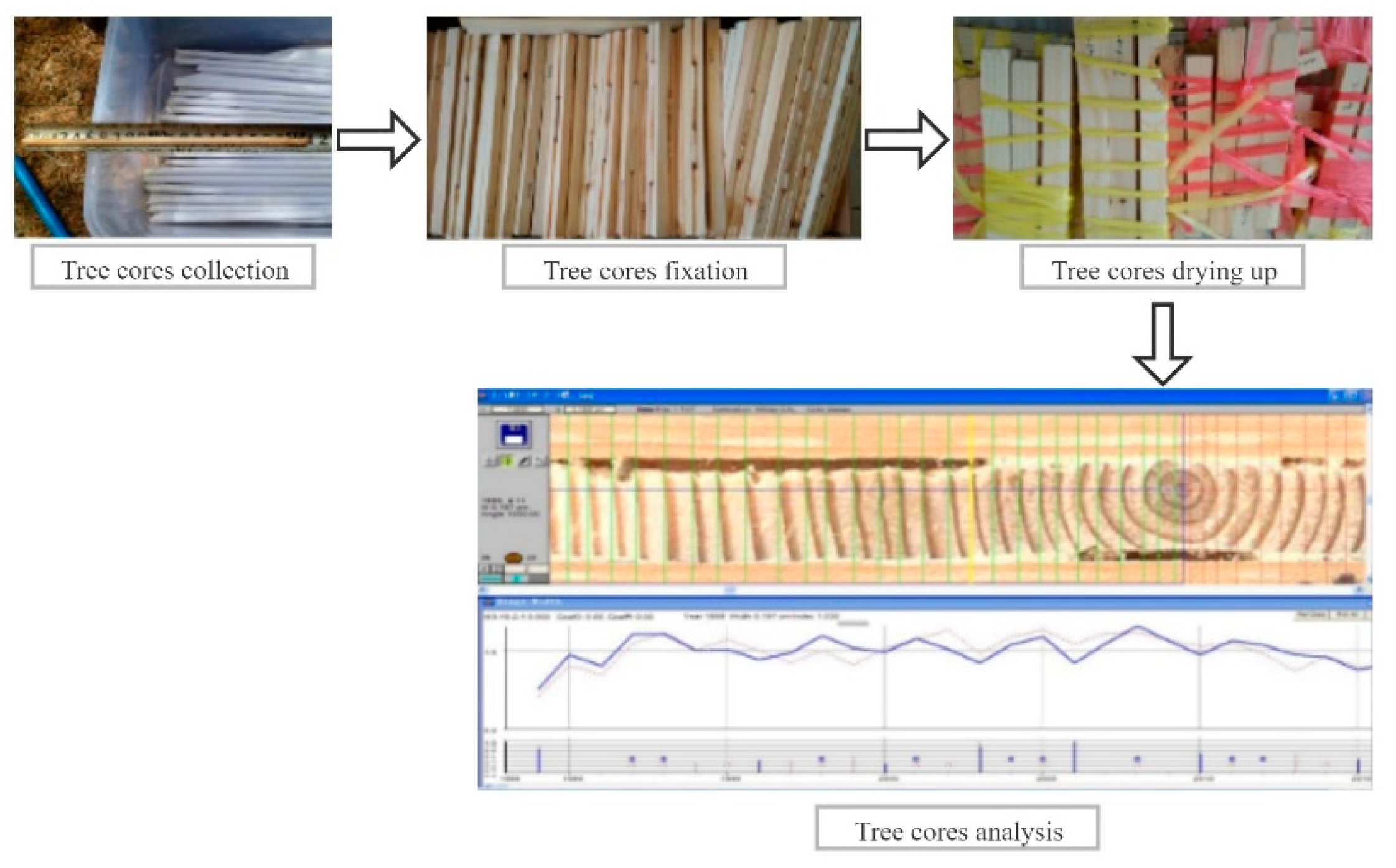
4.2.2. Field Survey Data
4.2.3. Remote Sensing Data
4.2.4. Soil Texture Data
4.3. BEPS Model
4.4. Trend Analysis
4.5. Partial Correlation Analysis
4.6. Lag Analysis
- (1)
- Calculate the correlation coefficient between the vegetation NPP and precipitation in July between 2003 and 2012, i.e., for a lag period of 0 months (lag0) at a cumulative number of lag months of 1.
- (2)
- Calculate the correlation coefficient between the vegetation NPP in July and cumulative precipitation in June and July between 2003 and 2012, i.e., for a lag period of 0 months (lag0) and a cumulative number of lag months of 2.
- (3)
- On the same basis, calculate the correlation coefficients of the vegetation NPP for a lag period of 0 months and the cumulative precipitation of 3, 4, and 5 months.
- (4)
- Calculate the correlation coefficient between the vegetation NPP in July and precipitation in June, i.e., for a lag period of 1 month (lag 1) and a cumulative number of lag months of 1.
- (5)
- Calculate the correlation coefficient between the vegetation NPP in July and the cumulative precipitation in May and June, i.e., for a lag period of 1 month (lag 1) and a cumulative number of lag months of 2.
- (6)
- On the same basis, calculate the correlation coefficients between the vegetation NPP in July at a lag period of 1 month and the cumulative precipitation of 3, 4, and 5 months.
- (7)
- On the same basis, calculate the correlation coefficients of the vegetation NPP in July for different lag periods with different cumulative numbers of lag months.
4.7. Evaluation and Analysis of the Modeled Estimates
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dlugokencky, E.; Tans, P. Trends in Atmospheric Carbon Dioxide, National Oceanic & Atmospheric Administration, Earth System Research Laboratory (NOAA/ESRL). 2019. Available online: http://www.esrl.noaa.gov/gmd/ccgg/trends/global.html/ (accessed on 4 September 2018).
- Le Quéré, C.; Andrew, R.M.; Friedlingstein, P.; Sitch, S.; Hauck, J.; Pongratz, J.; Pickers, P.A.; Korsbakken, J.I.; Peters, G.P.; Canadell, J.G.; et al. Global Carbon Budget 2018. Earth Syst. Sci. Data 2018, 10, 2141–2194. [Google Scholar] [CrossRef] [Green Version]
- Beer, C.; Reichstein, M.; Tomelleri, E.; Ciais, P.; Jung, M.; Carvalhais, N.; Rödenbeck, C.; Arain, M.A.; Baldocchi, D.; Bonan, G.B.; et al. Terrestrial gross carbon dioxide uptake: Global distribution and covariation with climate. Science 2010, 329, 834–838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Masri, B.; Barman, R.; Meiyappan, P.; Song, Y.; Liang, M.; Jain, A.K. Carbon dynamics in the Amazonian Basin: Integration of eddy covariance and ecophysiological data with a land surface model. Agric. For. Meteorol. 2013, 182–183, 156–167. [Google Scholar] [CrossRef]
- Piao, S.; Fang, J.; Zhou, L.; Zhu, B.; Tan, K.; Tao, S. Changes in vegetation net primary productivity from 1982 to 1999 in China. Glob. Biogeochem. Cycles 2005, 19, 2027. [Google Scholar] [CrossRef] [Green Version]
- Zhao, M.; Running, S.W. Drought-Induced Reduction in Global Terrestrial Net Primary Production from 2000 Through. Science 2010, 329, 940–943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Running, S.W.; Hunt, E.R. Generalization of a forest ecosystem process model for other biomes, Biome-BGC, and an application for global-scale models. Scaling processes between leaf and landscape levels. In Scaling Physiological Processes: Leaf to Globe; Ehleringer, J.R., Field, C.B., Eds.; Academic Press: San Diego, CA, USA, 1993; pp. 141–158. [Google Scholar]
- Matsushita, B.; Xu, M.; Chen, J.; Kameyama, S.; Tamura, M. Estimation of regional net primary productivity (NPP) using a process-based ecosystem model: How important is the accuracy of climate data. Ecol. Model. 2004, 178, 371–388. [Google Scholar] [CrossRef]
- Cuddington, K.; Fortin, M.-J.; Gerber, L.R.; Hastings, A.; Liebhold, A.; O’Connor, M.; Ray, C. Process-based models are required to manage ecological systems in a changing world. Ecosphere 2013, 4, 1–12. [Google Scholar] [CrossRef]
- Liu, J. A process-based boreal ecosystem productivity simulator using remote sensing inputs. Remote Sens. Environ. 1997, 62, 158–175. [Google Scholar] [CrossRef]
- Zhang, F.; Chen, J.M.; Chen, J.; Gough, C.M.; Martin, T.A.; Dragoni, D. Evaluating spatial and temporal patterns of MODIS GPP over the conterminous U.S. against flux measurements and a process model. Remote Sens. Environ. 2012, 124, 717–729. [Google Scholar] [CrossRef]
- Zhang, F.; Ju, W.; Shen, S.; Wang, S.; Yu, G.; Han, S. Variations of Terrestrial Net Primary Productivity in East Asia. Terr. Atmos. Ocean. Sci. 2012, 23, 425. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Chen, J.M.; Cihlar, J. Mapping evapotranspiration based on remote sensing: An application to Canada’s landmass. Water Resour. Res. 2003, 39. [Google Scholar] [CrossRef]
- Wang, P.; Xie, D.; Zhou, Y.; E., Y.; Zhu, Q. Estimation of net primary productivity using a process-based model in Gansu Province, Northwest China. Environ. Earth Sci. 2013, 71, 647–658. [Google Scholar] [CrossRef]
- Peng, C.; Zhou, X.; Zhao, S.; Wang, X.; Zhu, B.; Piao, S.; Fang, J. Quantifying the response of forest carbon balance to future climate change in Northeastern China: Model validation and prediction. Glob. Planet. Chang. 2009, 66, 179–194. [Google Scholar] [CrossRef]
- Xie, Y.; Lei, X.; Shi, J. Impacts of climate change on biological rotation of Larix olgensis plantations for timber production and carbon storage in northeast China using the 3-PGmix model. Ecol. Model. 2020, 435, 109267. [Google Scholar] [CrossRef]
- Kumari, S.; Singh, T.P.; Prasad, S. Climate Smart Agriculture and Climate Change. Int. J. Curr. Microbiol. Appl. Sci. 2019, 8, 1112–1137. [Google Scholar] [CrossRef]
- Zhu, W.-Q.; Pan, Y.-Z.; Liu, X.; Wang, A.-L. Spatio-temporal distribution of net primary productivity along the Northeast China Transect and its response to climatic change. J. For. Res. 2006, 17, 93–98. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhang, L.; Xiao, J.; Williams, C.A.; Vitkovskaya, I.; Bao, A. Spatiotemporal transition of institutional and socioeconomic impacts on vegetation productivity in Central Asia over last three decades. Sci. Total Environ. 2019, 658, 922–935. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.H.; Zhao, X.; Liang, S.L.; Zhou, T.; Huang, K.C.; Tang, B.J.; Zhao, W.Q. Time-lag effects of global vegetation responses to climate change. Glob. Chang. Biol. 2015, 21, 3520–3531. [Google Scholar] [CrossRef]
- Wen, Y.; Liu, X.; Yang, J.; Lin, K.; Du, G. NDVI indicated inter-seasonal non-uniform time-lag responses of terrestrial vegetation growth to daily maximum and minimum temperature. Glob. Planet. Chang. 2019, 177, 27–38. [Google Scholar] [CrossRef]
- Marino, S.; Hogue, I.B.; Ray, C.J.; Kirschner, D.E. A methodology for performing global uncertainty and sensitivity analysis in systems biology. J. Theor. Biol. 2008, 254, 178–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michele, C.; Stefano, T.; Andrea, S. Sensitivity and uncertainty analysis in spatial modelling based on GIS. Agric. Ecosyst. Environ. 2000, 81, 71–79. [Google Scholar]
- Ruiz, J.; Pulido, M. Parameter Estimation Using Ensemble-Based Data Assimilation in the Presence of Model Error. Mon. Weather Rev. 2015, 143, 1568–1582. [Google Scholar] [CrossRef] [Green Version]
- Bhattarai, P.; Zheng, Z.; Bhatta, K.; Adhikari, Y.; Zhang, Y. Climate-Driven Plant Response and Resilience on the Tibetan Plateau in Space and Time: A Review. Plants 2021, 10, 480. [Google Scholar] [CrossRef]
- Braswell, B.H.; Schimel, D.S.; Linder, E.; Moore, B. The Response of Global Terrestrial Ecosystems to Interannual Temperature Variability. Science 1997, 278, 870–873. [Google Scholar] [CrossRef]
- Wang, J.; Rich, P.M.; Price, K.P. Temporal responses of NDVI to precipitation and temperature in the central Great Plains, USA. Int. J. Remote Sens. 2003, 24, 2345–2364. [Google Scholar] [CrossRef]
- Bala, G.; Caldeira, K.; Wickett, M.; Phillips, T.J.; Lobell, D.B.; Delire, C.; Mirin, A. Combined climate and carbon-cycle effects of large-scale deforestation. Proc. Natl. Acad. Sci. USA 2007, 104, 6550–6555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malhi, Y.; Roberts, J.T.; Betts, R.A.; Killeen, T.J.; Li, W.; Nobre, C.A. Climate Change, Deforestation, and the Fate of the Amazon. Science 2008, 319, 169–172. [Google Scholar] [CrossRef] [Green Version]
- Choat, B.; Jansen, S.; Brodribb, T.J.; Cochard, H.; Delzon, S.; Bhaskar, R.; Bucci, S.J.; Feild, T.S.; Gleason, S.M.; Hacke, U.G.; et al. Global convergence in the vulnerability of forests to drought. Nature 2012, 491, 752–755. [Google Scholar] [CrossRef] [Green Version]
- Sterling, S.M.; Ducharne, A.; Polcher, J. The impact of global land-cover change on the terrestrial water cycle. Nat. Clim. Chang. 2012, 3, 385–390. [Google Scholar] [CrossRef]
- Brando, P.M.; Balch, J.K.; Nepstad, D.C.; Morton, D.C.; Putz, F.E.; Coe, M.T.; Silvério, D.; Macedo, M.N.; Davidson, E.A.; Nóbrega, C.C.; et al. Abrupt increases in Amazonian tree mortality due to drought-fire interactions. Proc. Natl. Acad. Sci. USA 2014, 111, 6347–6352. [Google Scholar] [CrossRef] [Green Version]
- Yang, K.; He, J.; Tang, W.; Qin, J.; Cheng, C.C. On downward shortwave and longwave radiations over high altitude regions: Observation and modeling in the Tibetan Plateau. Agric. For. Meteorol. 2010, 150, 38–46. [Google Scholar] [CrossRef]
- Yan, M.; Tian, X.; Li, Z.; Chen, E.; Li, C.; Fan, W. A long-term simulation of forest carbon fluxes over the Qilian Mountains. Int. J. Appl. Earth Obs. Geoinf. 2016, 52, 515–526. [Google Scholar] [CrossRef] [Green Version]
- Tian, X.; Van Der Tol, C.; Su, Z.; Li, Z.; Chen, E.; Li, X.; Yan, M.; Chen, X.; Wang, X.; Pan, X.; et al. Simulation of Forest Evapotranspiration Using Time-Series Parameterization of the Surface Energy Balance System (SEBS) over the Qilian Mountains. Remote Sens. 2015, 7, 15822–15843. [Google Scholar] [CrossRef] [Green Version]
- Correa-Díaz, A.; Silva, L.C.R.; Horwath, W.R.; Gómez-Guerrero, A.; Vargas-Hernández, J.; Villanueva-Díaz, J.; Velázquez-Martínez, A.; Suárez-Espinoza, J. Linking Remote Sensing and Dendrochronology to Quantify Climate-Induced Shifts in High-Elevation Forests Over Space and Time. J. Geophys. Res. Biogeosci. 2019, 124, 166–183. [Google Scholar] [CrossRef]
- Robinson, W.J.; Evans, R. A microcomputer-based tree-ring measuring system. Tree Ring Bull. 1980, 40, 59–64. [Google Scholar]
- Wu, G.; Feng, Z.W. Study on the biomass of LARIX SPP. Forest community in the frigid-temperate zone and the temperate zone of China. J. Northeast. For. Univ. 1995, 23, 95–101. [Google Scholar]
- Xiao, Z.; Liang, S.; Wang, J.; Chen, P.; Yin, X.; Zhang, L.; Song, J. Use of General Regression Neural Networks for Generating the GLASS Leaf Area Index Product from Time-Series MODIS Surface Reflectance. IEEE Trans. Geosci. Remote. Sens. 2014, 52, 209–223. [Google Scholar] [CrossRef]
- Liu, J.; Chen, J.M.; Cihlar, J.; Chen, W. Net primary productivity distribution in the BOREAS region from a process model using satellite and surface data. J. Geophys. Res. Space Phys. 1999, 104, 27735–27754. [Google Scholar] [CrossRef]
- Chen, J.; Liu, J.; Cihlar, J.; Goulden, M. Daily canopy photosynthesis model through temporal and spatial scaling for remote sensing applications. Ecol. Model. 1999, 124, 99–119. [Google Scholar] [CrossRef] [Green Version]
- Birkes, D.; Dodge, Y. Alternative Methods of Regression; John Wiley & Sons: New York, NY, USA, 1993. [Google Scholar]
- Rundquist, B.C.; Harrington, J.A., Jr. The Effects of Climatic Factors on Vegetation Dynamics of Tallgrass and Shortgrass Cover. Geocarto Int. 2000, 15, 33–38. [Google Scholar] [CrossRef]
- Anderson, L.O.; Malhi, Y.; Aragão, L.E.O.C.; Ladle, R.; Arai, R.; Barbier, N.; Phillips, O. Remote sensing detection of droughts in Amazonian forest canopies. New Phytol. 2010, 187, 733–750. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; De Jeu, R.; Liu, Y.; Van der Werf, G.; Dolman, A. Using satellite based soil moisture to quantify the water driven variability in NDVI: A case study over mainland Australia. Remote Sens. Environ. 2014, 140, 330–338. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, M.; Xue, M.; Zhang, L.; Tian, X.; Chen, B.; Dong, Y. A Decade’s Change in Vegetation Productivity and Its Response to Climate Change over Northeast China. Plants 2021, 10, 821. https://doi.org/10.3390/plants10050821
Yan M, Xue M, Zhang L, Tian X, Chen B, Dong Y. A Decade’s Change in Vegetation Productivity and Its Response to Climate Change over Northeast China. Plants. 2021; 10(5):821. https://doi.org/10.3390/plants10050821
Chicago/Turabian StyleYan, Min, Mei Xue, Li Zhang, Xin Tian, Bowei Chen, and Yuqi Dong. 2021. "A Decade’s Change in Vegetation Productivity and Its Response to Climate Change over Northeast China" Plants 10, no. 5: 821. https://doi.org/10.3390/plants10050821