
Effects of Shade and Planting Methods on the Growth of Heracleum moellendorffii and Adenophora divaricata in Different Soil Moisture and Nutrient Conditions



Abstract
:1. Introduction
2. Results
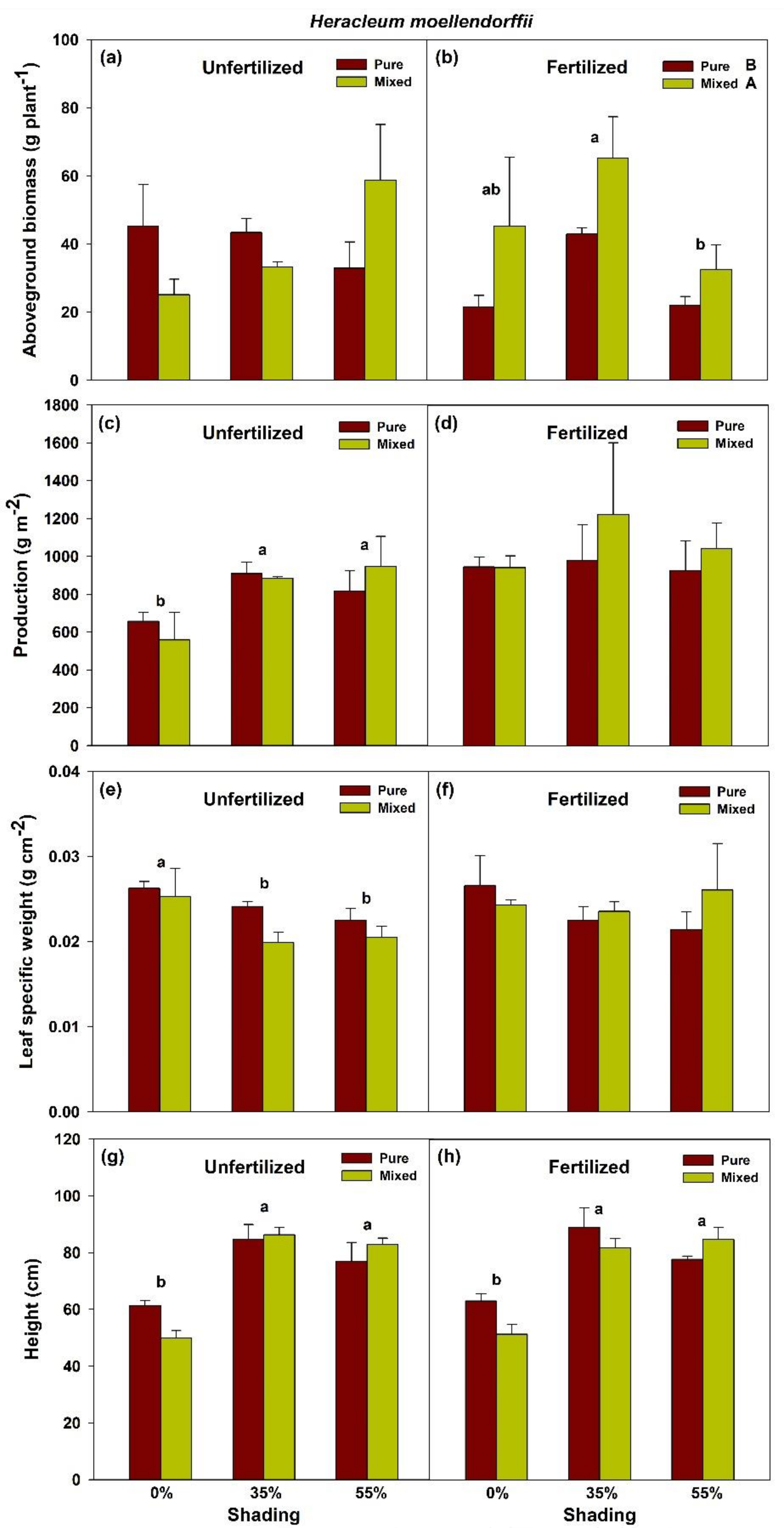
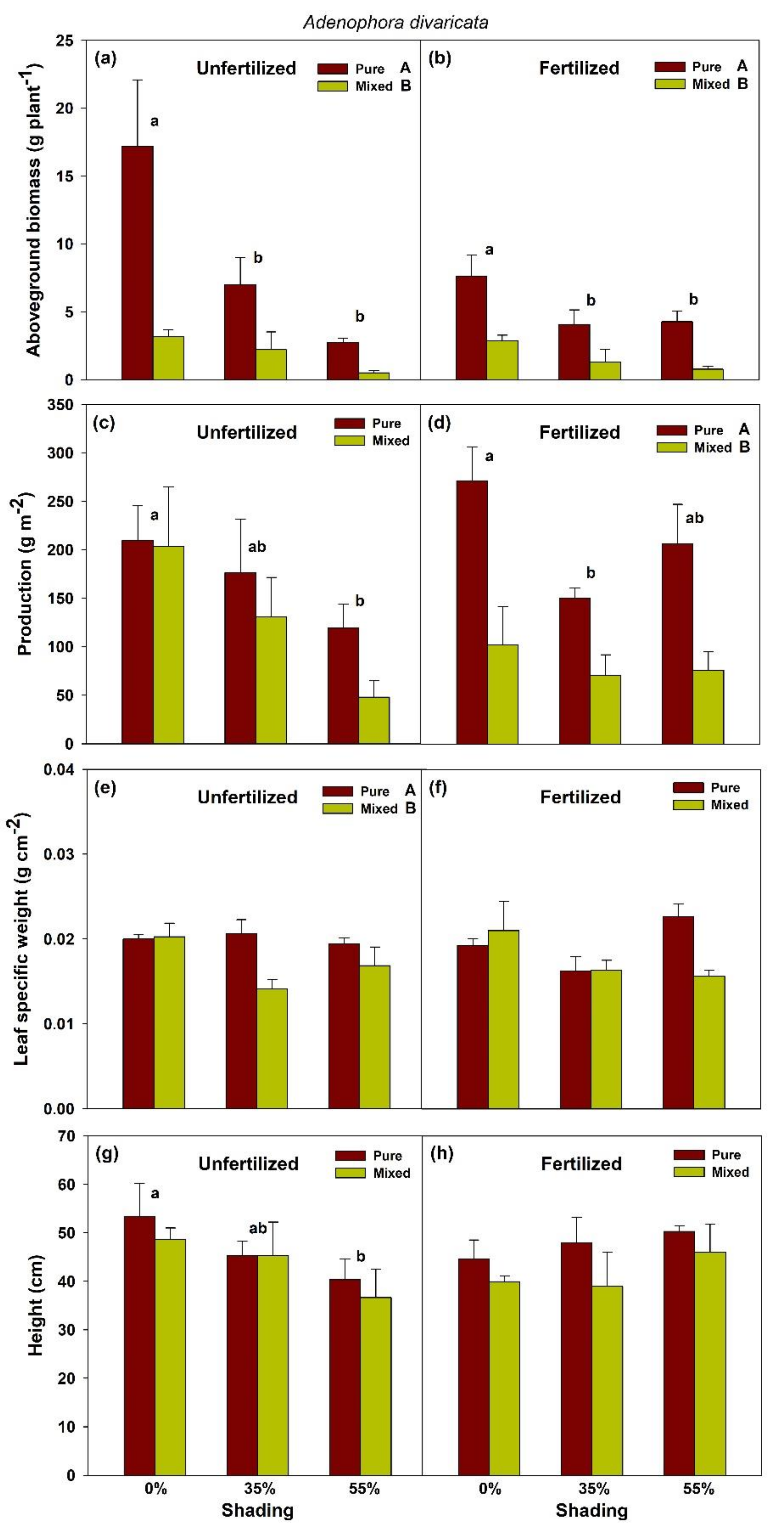
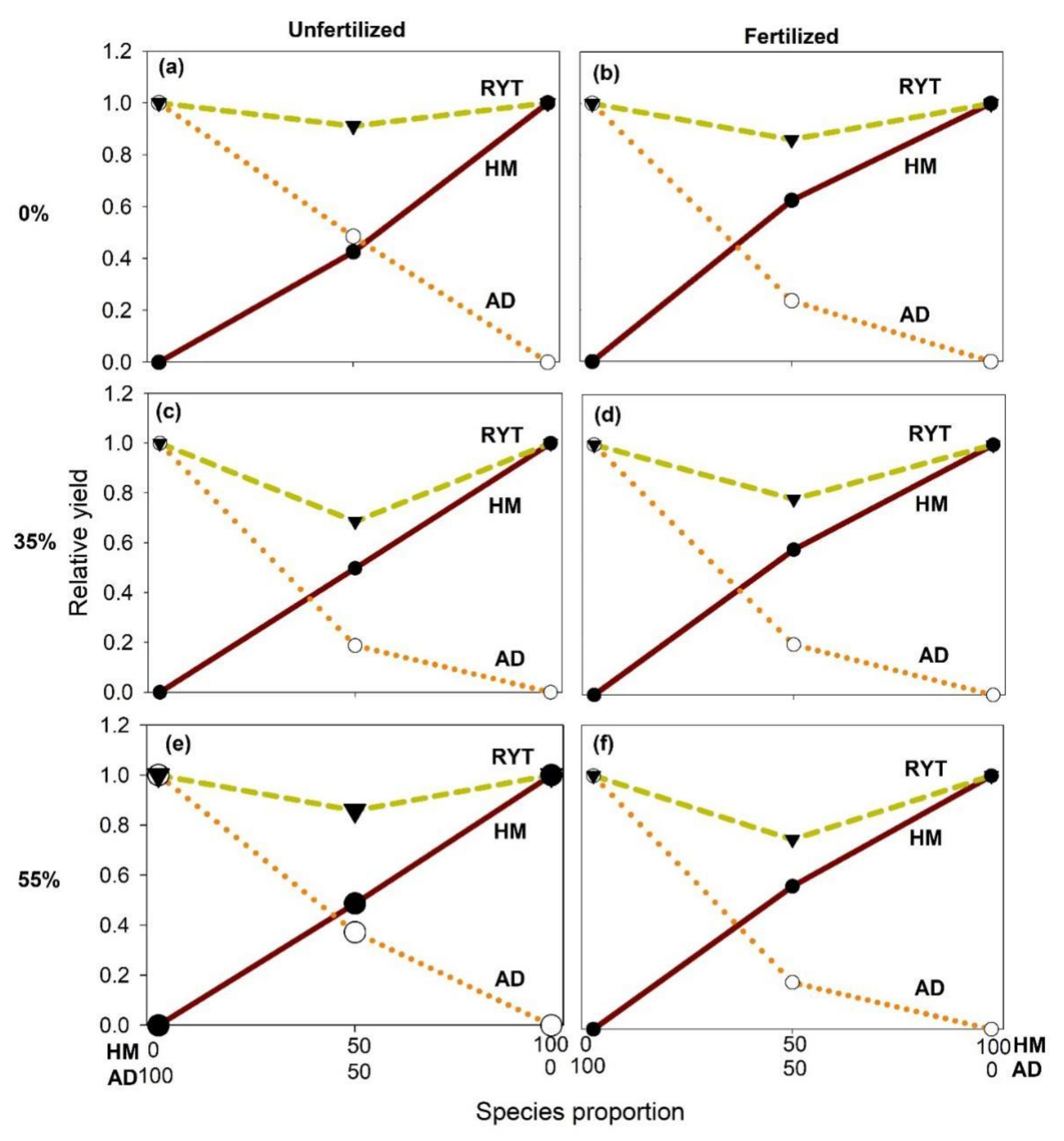
2.1. Effects of Shade and Planting Methods on the Growth of Unfertilized and Fertilized H. moellendorffii and A. divaricata
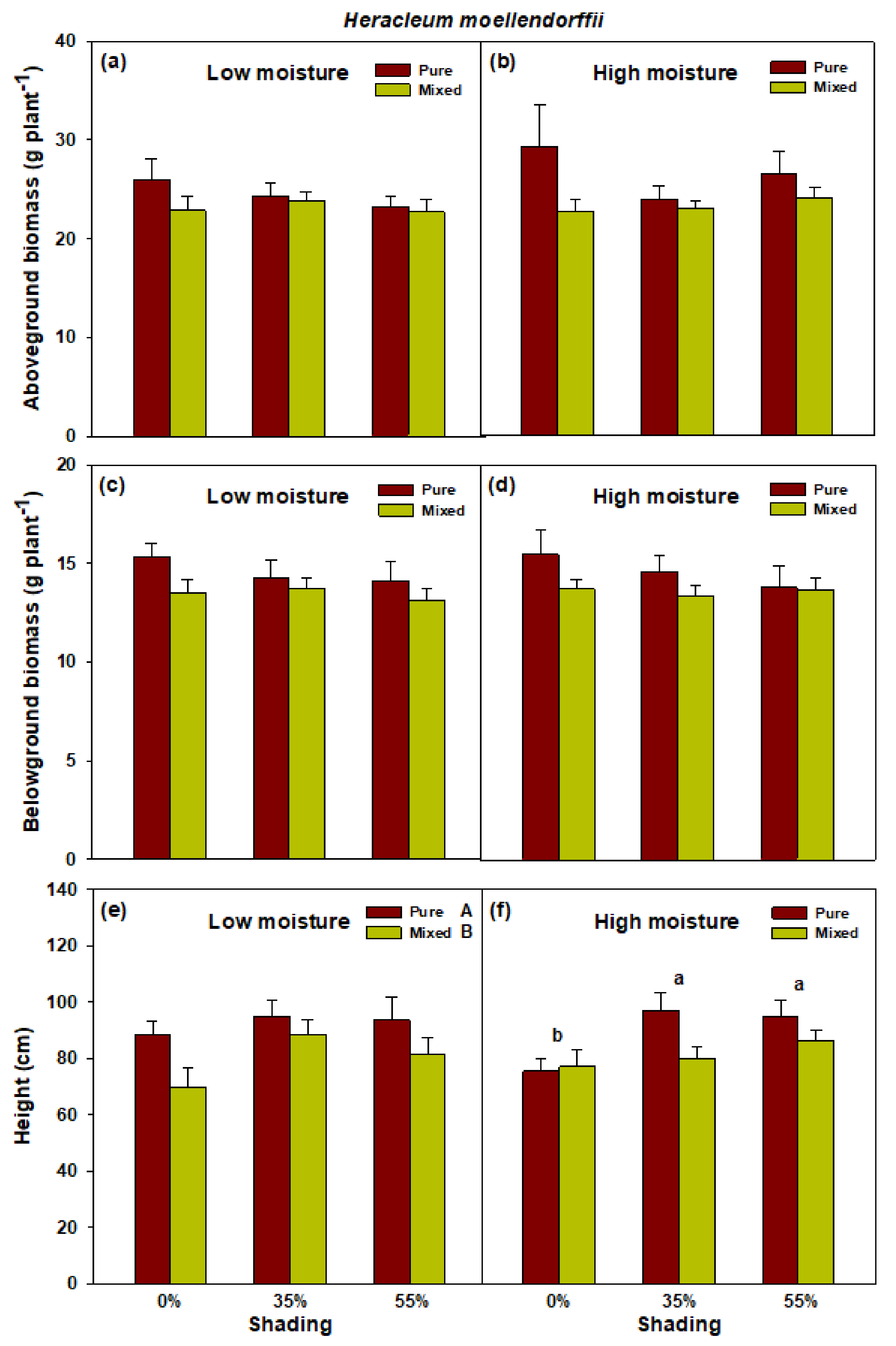
2.2. Effects of Shade and Planting Methods on the Growth of H. moellendorffii and A. divaricata under High and Low Soil Moisture Conditions
3. Discussion
4. Materials and Methods
4.1. Study Site and Experimental Materials
4.2. Experimental Design
4.3. Data Collection
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sartoretto, E.; Henriot, C.; Bassalang, M.M.; Nguiffo, S. How Existing Legal Frameworks Shape Forest Conversion To Agriculture: A Study of the Congo Basin. Food and Agriculture Organization of the United Nations, Rome. 2017. Available online: https://www.fao.org/3/i7947e/i7947e.pdf (accessed on 12 October 2021).
- FAO. State of the World’s Forests 2016 Forests and Agriculture: Land-Use Challenges and Opportunities. Food and Agriculture Organization of the United Nations, Rome. 2016. Available online: https://www.fao.org/3/i5588e/i5588e.pdfs (accessed on 12 October 2021).
- Mudge, K.; Gabriel, S. Farming the Woods: An Integrated Permaculture Approach to Growing Food and Medicinals in Temperate Forests; Chelsea Green Publishing: Winsor County, VT, USA, 2014; p. 359. [Google Scholar]
- Chamberlain, J.L.; Mitchell, D.; Brigham, T.; Hobby, T.; Zabek, L.; Davis, J. Forest farming practices. In North American Agroforestry: An Integrated Science and Practice, 2nd ed.; Garrett, H.E., Ed.; American Society of Agronomy: Madison, WI, USA, 2009; pp. 219–256. [Google Scholar]
- Becker, B.; Workman, S. Farming in the Forests of Florida; University of Florida: Gainesville, FL, USA, 2003; p. 6. [Google Scholar]
- Workman, S.W.; Long, A.J.; Mohan, S.; Monroe, M.C. Agroforestry: Options for landowners; University of Florida: Gainesville, FL, USA, 2002; p. 4. [Google Scholar]
- Suleiman, M.S.; Wasonga, V.O.; Mbau, J.S.; Suleiman, A.; Elhadi, Y.A. Non-timber forest products and their contribution to households income around Falgore Game Reserve in Kano, Nigeria. Ecol. Process. 2017, 6, 23–36. [Google Scholar] [CrossRef] [Green Version]
- Su, X.; Wang, M.; Huang, Z.; Fu, S.; Chen, H.Y.H. Forest understorey vegetation: Colonization and the availability and heterogeneity of resources. Forests 2019, 10, 944. [Google Scholar] [CrossRef] [Green Version]
- Pierson, E.A.; Mack, R.N.; Black, R.A. The effect of shading on photosynthesis, growth, and regrowth following defoliation for Bromus tectorum. Oecologia 1990, 84, 534–543. [Google Scholar] [CrossRef] [PubMed]
- Kabir, M.Y.; Díaz-Pérez, J.C.; Nambeesan, S.U. Effect of shade levels on plant growth, physiology, and fruit yield in bell pepper (Capsicum annuum L.). Acta. Hortic 2020, 1268, 311–318. [Google Scholar] [CrossRef]
- Thakur, M.; Bhatt, V.; Kumar, R. Effect of shade level and mulch type on growth, yield and essential oil composition of damask rose (Rosa damascena Mill.) under mid hill conditions of Western Himalayas. PLoS ONE 2019, 14, e0214672. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, S.; Bhadouria, R.; Srivastava, P.; Devi, R.S.; Chaturvedi, R.; Raghubanshi, A.S. Effects of light availability on leaf attributes and seedling growth of four tree species in tropical dry forest. Ecol. Process. 2020, 9, 1–16. [Google Scholar] [CrossRef]
- Gerhardt, K.; Hytteborn, H. Natural dynamics and regeneration methods in tropical dry forest-an introduction. J. Veg. Sci. 1992, 3, 361–364. [Google Scholar] [CrossRef]
- Huante, P.; Rincón, E. Responses to light changes in tropical deciduous woody seedlings with contrasting growth rates. Oecologia 1997, 113, 53–66. [Google Scholar] [CrossRef] [PubMed]
- Halpern, C.B.; Lutz, J.A. Canopy closure exerts weak controls on understory dynamics: A 30-year study of overstory–understory interactions. Ecol. Monogr. 2013, 83, 221–237. [Google Scholar] [CrossRef] [Green Version]
- Samuel, A.D.; Bungau, S.; Tit, D.M.; Melinte, C.E.; Purza, L.; Badea, G.E. Effects of long term application of organic and mineral fertilizers on soil enzymes. Rev. Chim. 2018, 69, 2608–2612. [Google Scholar] [CrossRef]
- Dao, H.T.T.; Seo, J.M.; Hernandez, J.O.; Han, S.H.; Youn, W.B.; An, J.Y.; Park, B.B. Effective placement methods of vermicompost application in urban tree species: Implications for sustainable urban afforestation. Sustainability 2020, 12, 5822. [Google Scholar] [CrossRef]
- Casey, C.A.; Mangan, F.X.; Herbert, S.J.; Barker, A.V.; Carter, A.K. The effect of light intensity and nitrogen fertilization on plant growth and leaf quality of Ngo Gai (Eryngium foetidum L.) in Massachusetts. Acta. Hortic. 2004, 629, 215–229. [Google Scholar] [CrossRef]
- Canham, C.D.; Berkowitz, A.R.; Kelly, V.R.; Lovett, G.M.; Ollinger, S.V.; Schnurr, J. Biomass allocation and multiple resource limitation in tree seedlings. Can. J. For. Res. 1996, 26, 1521–1530. [Google Scholar] [CrossRef]
- Buss, J.; Stratechuk, K.; Pinno, B.D. Growth and competition among understory plants varies with reclamation soil and fertilization. Ecol. Process. 2018, 7, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Li, F.H.; Yu, P.; Song, C.H.; Wu, J.J.; Tian, Y.; Wu, X.F.; Zhang, X.W.; Liu, Y.M. Differential protein analysis of Heracleum moellendorffii Hance seeds during stratification. Plant Physiol. Biochem. 2019, 145, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Fu, P.; Chen, Y.A. Herbaceous Flora of Northeast China; Science Press: Changchun, China, 1998. [Google Scholar]
- Cho, S.Y.; Kim, Y.S.; Jeon, Y.H. First Report of Cucumber Green Mottle mosaic virus infecting Heracleum moellendorffii in Korea. Plant Dis. 2015, 99, 897. [Google Scholar] [CrossRef]
- Cheon, K.S.; Kim, K.A.; Yoo, K.O. The complete chloroplast genome sequences of three adenophora species and comparative analysis with campanuloid species (Campanulaceae). PLoS ONE 2017, 12, e0183652. [Google Scholar] [CrossRef] [Green Version]
- Alam, M.B.; Seo, B.J.; Zhao, P.; Lee, S.H. Anti-Melanogenic activities of Heracleum moellendorffii via ERK1/2-mediated MITF Downregulation. Int. J. Mol. Sci. 2016, 17, 1844. [Google Scholar] [CrossRef] [Green Version]
- Jang, J.; Park, S.H.; Chang, K.S.; Ji, S.J.; Jung, S.Y.; Lee, H.J.; Hwang, H.S.; Lee, Y.M. Diversity of vascular plants in Daebudo and its adjacent regions, Korea. J. Asia Pac. Biodiverse 2013, 6, 261–280. [Google Scholar] [CrossRef] [Green Version]
- Kong, W.S.; Watts, D. Arctic-alpine & alpine floristic elements. In The Plant Geography of Korea, 1st ed.; Kong, W.S., Watts, P., Eds.; Springer: Dordrecht, The Netherlands, 1993; Volume 19, pp. 83–104. [Google Scholar] [CrossRef]
- Kim, S.J.; Yu, D.J.; Kim, T.C.; Lee, H.J. Growth and photosynthetic characteristics of blueberry (Vaccinium corymbosum cv. Bluecrop) under various shade levels. Sci. Hortic. 2011, 129, 486–492. [Google Scholar] [CrossRef]
- Rengel, Z. Mechanistic simulation models of nutrient uptake: A review. Plant Soil. 1993, 152, 161–173. [Google Scholar] [CrossRef]
- Meena, S.K.; Pandey, R.; Sharma, S.; Kumar, T.; Singh, M.P.; Dikshit, H.K. Physiological Basis of Combined Stress Tolerance to Low Phosphorus and Drought in a Diverse Set of Mungbean Germplasm. Agronomy 2021, 11, 99. [Google Scholar] [CrossRef]
- Kupers, S.J.; Wirth, C.; Engelbrecht, B.M.; Hernández, A.; Condit, R.; Wright, S.J.; Rüger, N. Performance of tropical forest seedlings under shade and drought: An interspecific trade-off in demographic responses. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coomes, D.A.; Kunstler, G.; Canham, C.D.; Wright, E. A greater range of shade-tolerance niches in nutrient-rich forests: An explanation for positive richness-productivity relationships? J. Ecol. 2009, 97, 705–717. [Google Scholar] [CrossRef]
- Humbert, L.; Gagnon, D.; Kneeshaw, D.; Messier, C. A shade tolerance index for common understory species of northeastern North America. Ecol. Indic. 2007, 7, 195–207. [Google Scholar] [CrossRef] [Green Version]
- Nagashima, H.; Hikosaka, K. Plants in a crowded stand regulate their height growth so as to maintain similar heights to neighbours even when they have potential advantages in height growth. Ann. Bot. 2011, 108, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Smith, H. Light quality, photoperception, and plant strategy. Annu. Rev. Plant Physiol. 1982, 33, 481–518. [Google Scholar] [CrossRef]
- Weiner, J.; Thomas, S.C. Competition and allometry in three species of annual plants. Ecology 1992, 73, 648–656. [Google Scholar] [CrossRef] [Green Version]
- Niinemets, U.; Kull, O.; Tenhunen, J.D. An analysis of light effects on foliar morphology, physiology, and light interception in temperate deciduous woody species of contrasting shade tolerance. Tree Physiol. 1998, 18, 681–696. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Ren, X.; Jeong, H.K.; Wei, H.; Jeong, B.R. Growth and Physiological Responses of Adenophora triphylla (Thunb.) A.DC. Plug Seedlings to Day and Night Temperature Regimes. Agronomy 2018, 8, 173. [Google Scholar] [CrossRef] [Green Version]
- Haan, J.L.; Vasseur, L. Above and below Ground interactions in monoculture and Intercropping of onion and lettuce in Greenhouse conditions. Am. J. Plant Sci. 2014, 5, 3319–3327. [Google Scholar] [CrossRef] [Green Version]
- Mariotti, M.; Masoni, A.; Ercoli, L.; Arduini, I. Above- and below-ground competition between barley, wheat, lupin and vetch in a cereal and legume intercropping system. Grass Forage Sci. 2009, 64, 401–412. [Google Scholar] [CrossRef]
- Tilman, D. Plant Strategies and the Dynamics and Structure of Plant Communities; Princeton University Press: Princeton, NJ, USA, 1988; p. 376. [Google Scholar]
- Seo, J.M.; Dao, H.T.T.; Park, B.B. Growth and nutrient responses of Betula platyphylla, Larix kaempferi, and Chamaecyparis obtusa to different application methods of solid compound fertilizer. Turk. J. Agric. For. 2021, 45, 253–265. [Google Scholar] [CrossRef]
- Lee, E.; Han, S.K.; Im, S. Performance Analysis of Log Extraction by a Small Shovel Operation in Steep Forests of South Korea. Forests 2019, 10, 585. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Jiang, X.; Cheng, Y.; Yu, X. First Report of Powdery Mildew Caused by Erysiphe heraclei on Heracleum moellendorffii in China. Plant Dis. 2019, 103, 2690. [Google Scholar] [CrossRef]
- Ham, Y.A.; Choi, H.J.; Chung, M.J.; Ham, S.S. Component analysis and antioxidant activity of Adenophora triphylla. J. Korean Soc. Food Sci. Nutr. 2009, 38, 274–279. [Google Scholar] [CrossRef]
- Kim, H.S.; Lee, S.M.; Lee, J. Analysis of Vegetation-Environment Telationships of Main Wild Vegetables on Short-Term Income Forest Products, in Korea. Korean J. Environ. Ecol. 2019, 33, 447–452. [Google Scholar] [CrossRef]
- Carlyle, C.N.; Fraser, L.H.; Turkington, R. Using three pairs of competitive indices to test for changes in plant competition under different resource and disturbance levels. J. Veg. Sci. 2010, 21, 1025–1034. [Google Scholar] [CrossRef] [Green Version]
- Weigelt, A.; Jolliffe, P. Indices of plant competition. J. Ecol. 2003, 91, 707–720. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| pH | Organic Matter (%) | Total N (%) | Available P (mg kg−1) | CEC (cmolc kg−1) | Exchangeable Cations (cmolc kg−1) | |||
|---|---|---|---|---|---|---|---|---|
| K+ | Ca2+ | Mg2+ | Na+ | |||||
| 6.2 (0.1) | 1.63 (0.24) | 0.09 (0.01) | 102.3 (11.7) | 4.35 (0.35) | 0.18 (0.01) | 0.06 (0.00) | 3.51 (0.25) | 0.80 (0.02) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Youn, W.B.; Hernandez, J.O.; Park, B.B. Effects of Shade and Planting Methods on the Growth of Heracleum moellendorffii and Adenophora divaricata in Different Soil Moisture and Nutrient Conditions. Plants 2021, 10, 2203. https://doi.org/10.3390/plants10102203
Youn WB, Hernandez JO, Park BB. Effects of Shade and Planting Methods on the Growth of Heracleum moellendorffii and Adenophora divaricata in Different Soil Moisture and Nutrient Conditions. Plants. 2021; 10(10):2203. https://doi.org/10.3390/plants10102203
Chicago/Turabian StyleYoun, Woo Bin, Jonathan Ogayon Hernandez, and Byung Bae Park. 2021. "Effects of Shade and Planting Methods on the Growth of Heracleum moellendorffii and Adenophora divaricata in Different Soil Moisture and Nutrient Conditions" Plants 10, no. 10: 2203. https://doi.org/10.3390/plants10102203