
Biomass and Leaf Acclimations to Ultraviolet Solar Radiation in Juvenile Plants of Coffea arabica and C. canephora

 , , , ,
, , , ,  and
and 
Abstract
:1. Introduction
2. Material and Methods
2.1. Experimental Site, Species Description, and Light Microclimate
2.2. Plant Growth Traits
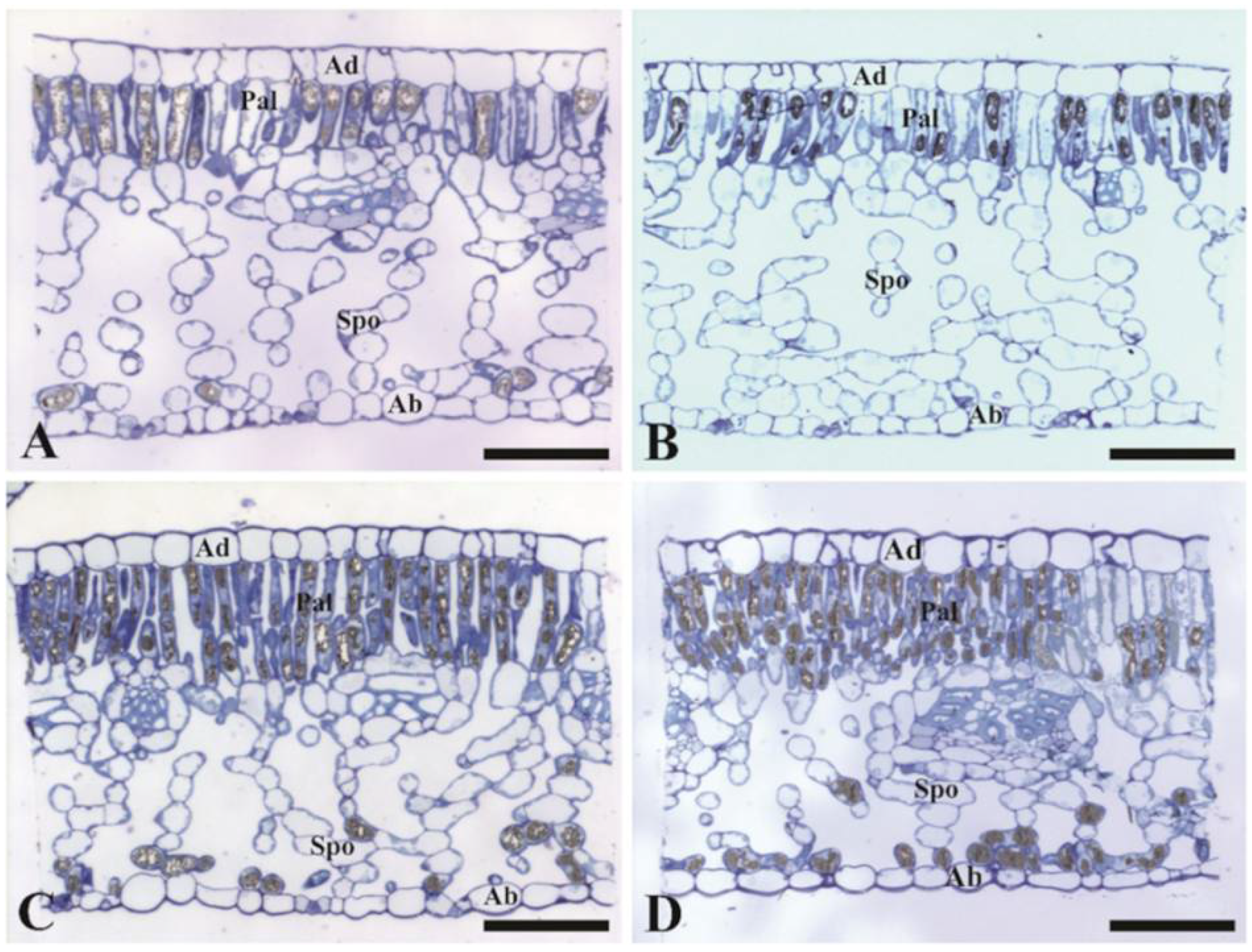
2.3. Leaf Anatomy
2.4. Photosynthetic Pigments Evaluation
2.5. Chlorophyll a Fluorescence
2.6. Statistical Analyses
3. Results
3.1. Light Microclimate
3.2. Plant Scale: Morphology and Growth Traits
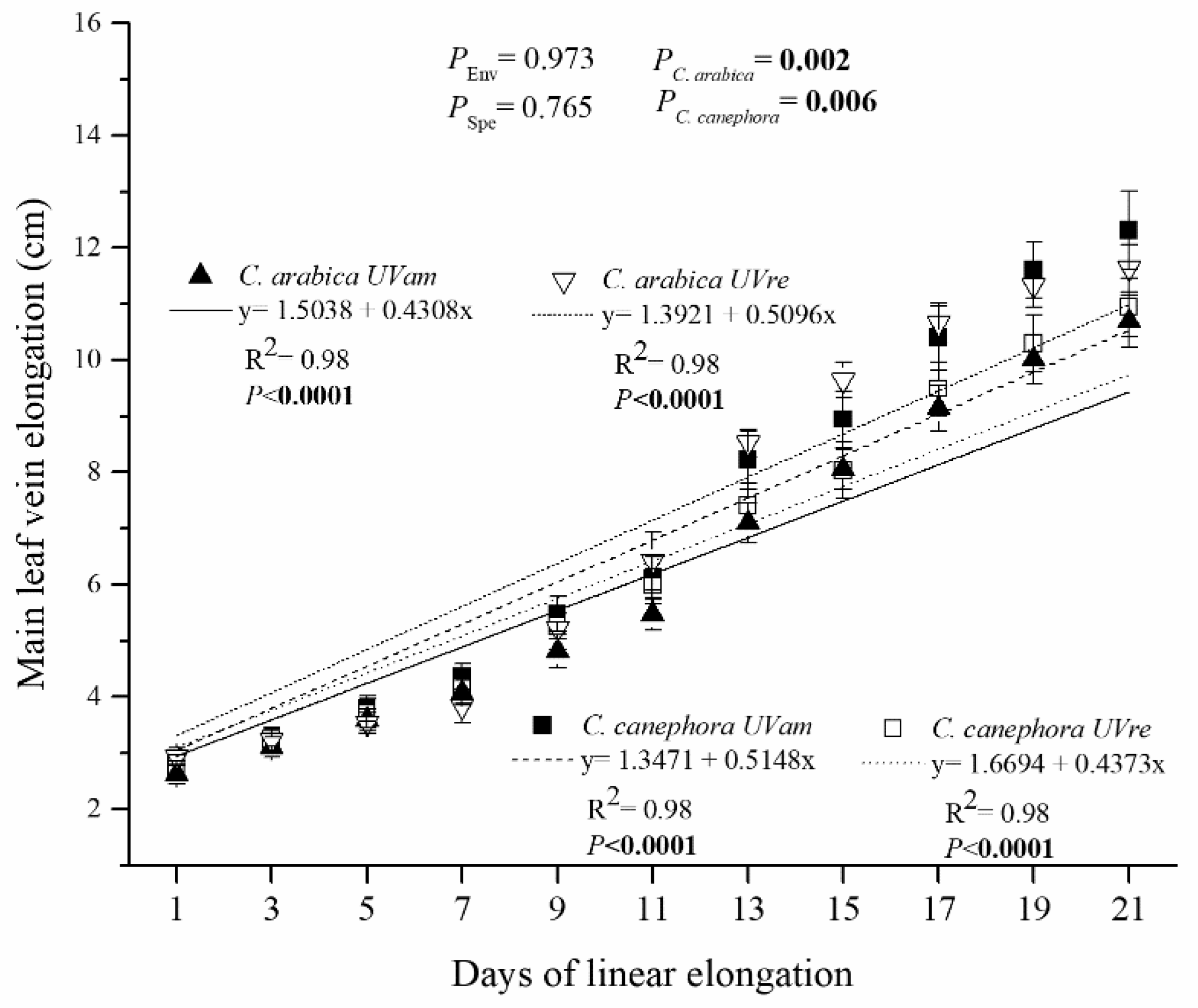
3.3. Leaf Scale: Tissue Thickness over the Vertical Cut and Leaf Elongation Rate
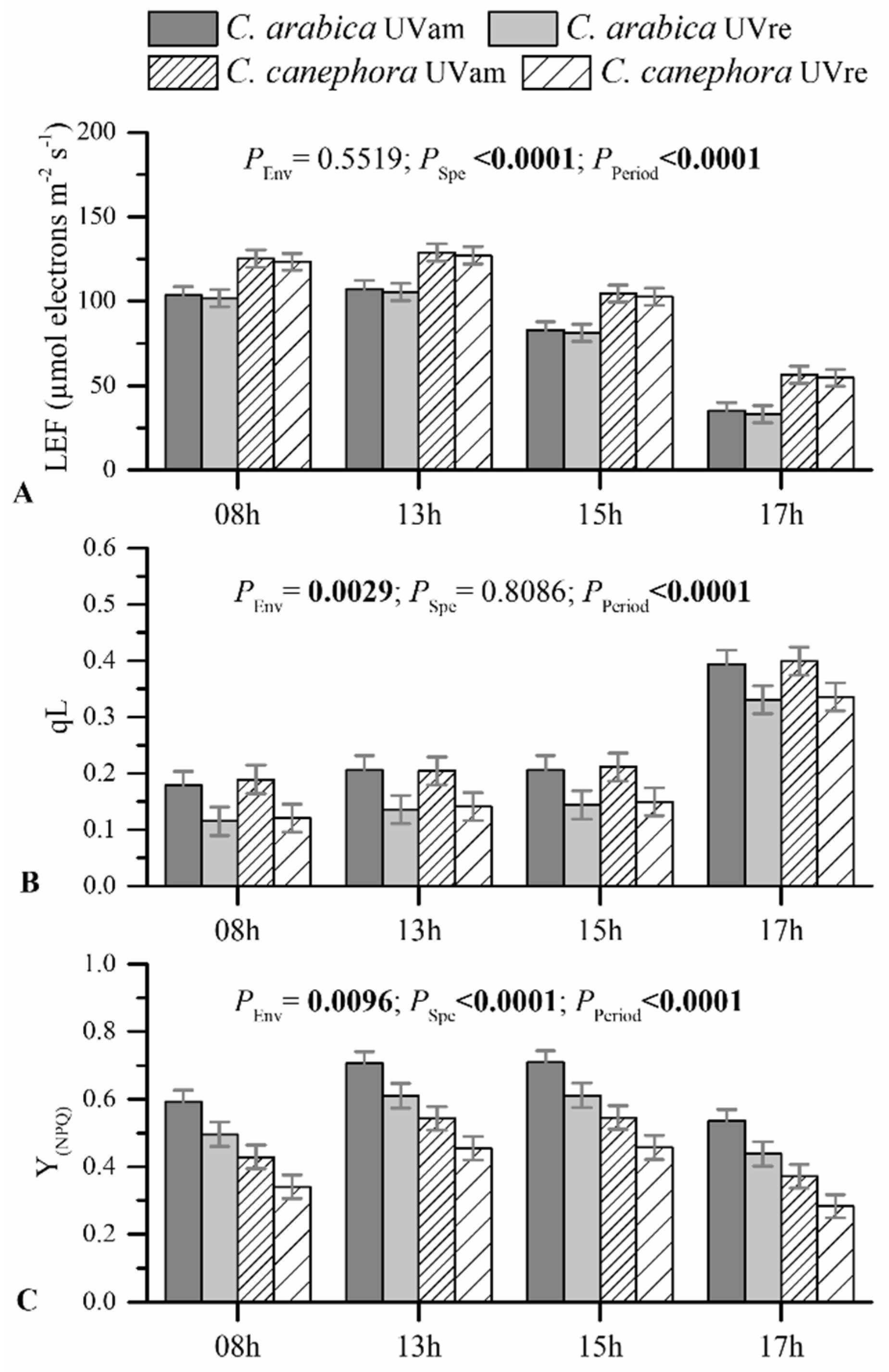
3.4. Leaf Photochemical Responses: Photosynthetic Pigments and Chlorophyll a Fluorescence
4. Discussion
4.1. Morphological and Anatomical Responses to UV Radiation
4.2. Photochemical Responses to UV Radiation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AbC | thickness of the abaxial cuticle layer |
| AdC | thickness of the adaxial cuticle layer |
| Chl | chlorophyll |
| DAT | day after transplanting |
| LEF | linear electron transport |
| PAR | photosynthetic active radiation |
| PSII | photosystem II |
| qL | fraction of PSII centers that were “open” |
| SD | stomatal density |
| SLM | specific leaf mass |
| UV | ultraviolet radiation |
| UVam | near ambient UV environment |
| UVre | reduced UV levels |
| YNPQ | estimate of the yield of energy dissipated through non-photochemical photoprotective processes |
References
- Banerjee, A.; Fyfe, J.C.; Polvani, L.M.; Waugh, D.; Chang, K.-L. A pause in Southern Hemisphere circulation trends due to the Montreal Protocol. Nat. Cell Biol. 2020, 579, 544–548. [Google Scholar] [CrossRef]
- Kataria, S.; Jajoo, A.; Guruprasad, K.N. Impact of increasing Ultraviolet-B (UV-B) radiation on photosynthetic processes. J. Photochem. Photobiol. B Biol. 2014, 137, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Bernhard, G.H.; Neale, R.E.; Barnes, P.W.; Neale, P.J.; Zepp, R.G.; Wilson, S.R.; Andrady, A.L.; Bais, A.F.; McKenzie, R.L.; Aucamp, P.J.; et al. Environmental effects of stratospheric ozone depletion, UV radiation and interactions with climate change: UNEP Environmental effects assessment panel, update 2019. Photochem. Photobiol. Sci. 2020, 19, 542–584. [Google Scholar] [CrossRef]
- Davis, A.P.; Govaerts, R.; Bridson, D.M.; Stoffelen, P. An annotated taxonomic conspectus of the genus Coffea (Rubiaceae). Bot. J. Linn. Soc. 2006, 152, 465–512. [Google Scholar] [CrossRef] [Green Version]
- DaMatta, F.M.; Ramalho, J.D.C. Impacts of drought and temperature stress on coffee physiology and production: A review. Braz. J. Plant Physiol. 2006, 18, 55–81. [Google Scholar] [CrossRef]
- DaMatta, F.M.; Rahn, E.; Läderach, P.; Ghini, R.; Ramalho, J.C. Why could the coffee crop endure climate change and global warming to a greater extent than previously estimated? Clim. Chang. 2018, 152, 167–178. [Google Scholar] [CrossRef] [Green Version]
- Kakani, V.; Reddy, K.; Zhao, D.; Sailaja, K. Field crop responses to ultraviolet-B radiation: A review. Agric. For. Meteorol. 2003, 120, 191–218. [Google Scholar] [CrossRef]
- Hectors, K.; Prinsen, E.; De Coen, W.; Jansen, M.A.K.; Guisez, Y. Arabidopsis thalianaplants acclimated to low dose rates of ultraviolet B radiation show specific changes in morphology and gene expression in the absence of stress symptoms. New Phytol. 2007, 175, 255–270. [Google Scholar] [CrossRef] [PubMed]
- Lidon, F.; Ramalho, J. Impact of UV-B irradiation on photosynthetic performance and chloroplast membrane components in Oryza sativa L. J. Photochem. Photobiol. B Biol. 2011, 104, 457–466. [Google Scholar] [CrossRef]
- Lidon, F.J.C.; Reboredo, F.H.; Leitão, A.E.; Silva, M.M.A.; Duarte, M.P.; Ramalho, J.C. Impact of UV-B radiation on photosynthesis—An overview. Emir. J. Food Agric. 2012, 24, 546–556. [Google Scholar] [CrossRef]
- Ramalho, J.C.; Pons, T.L.; Groeneveld, H.W.; Azinheira, H.G.; Nunes, M.A. Photosynthetic acclimation to high light conditions in mature leaves of Coffea arabica L.: Role of xanthophylls, quenching mechanisms and nitrogen nutrition. Funct. Plant Biol. (ex- Aust. J. Plant Physiol.) 2000, 27, 43–51. [Google Scholar] [CrossRef]
- Ramalho, J.C.; Marques, N.C.; Semedo, J.N.; Matos, M.C.; Quartin, V.L. Photosynthetic performance and pigment composition of leaves from two tropical species is determined by light quality. Plant Biol. 2002, 4, 112–120. [Google Scholar] [CrossRef]
- Rakocevic, M.; Ribeiro, R.V.; Marchiori, P.E.R.; Filizola, H.F.; Batista, E.R. Structural and functional changes in coffee trees after 4 years under free air CO2 enrichment. Ann. Bot. 2018, 121, 1065–1078. [Google Scholar] [CrossRef]
- Rakocevic, M.; Batista, E.R.; Pazianotto, R.A.A.; Scholz, M.B.S.; Souza, G.A.R.; Campostrini, E.; Ramalho, J.C. Leaf gas exchange and bean quality fluctuations over the whole canopy vertical profile of Arabic coffee cultivated under elevated CO2. Funct. Plant Biol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Verdaguer, D.; Jansen, M.A.; Llorens, L.; Morales, L.O.; Neugart, S. UV-A radiation effects on higher plants: Exploring the known unknown. Plant Sci. 2017, 255, 72–81. [Google Scholar] [CrossRef]
- Meijkamp, B.B.; Doodeman, G.; Rozema, J. The response of Vicia faba to enhanced UV-B radiation under low and near ambient PAR levels. In Responses of Plants to UV-B Radiation; Springer: Dordrecht, The Netherlands, 2001; pp. 135–146. [Google Scholar]
- Kataria, S.; Guruprasad, K.; Ahuja, S.; Singh, B. Enhancement of growth, photosynthetic performance and yield by exclusion of ambient UV components in C3 and C4 plants. J. Photochem. Photobiol. B Biol. 2013, 127, 140–152. [Google Scholar] [CrossRef]
- Zhang, L.; Allen, L.H.; Vaughan, M.M.; Hauser, B.A.; Boote, K.J. Solar ultraviolet radiation exclusion increases soybean internode lengths and plant height. Agric. For. Meteorol. 2014, 184, 170–178. [Google Scholar] [CrossRef]
- Krizek, D.T.; Mirecki, R.M.; Britz, S.J. Inhibitory effects of ambient levels of solar UV-A and UV-B radiation on growth of cucumber. Physiol. Plant. 1997, 100, 886–893. [Google Scholar] [CrossRef]
- Robson, T.M.; Klem, K.; Urban, O.; Jansen, M.A.K. Re-interpreting plant morphological responses to UV-B radiation. Plant Cell Environ. 2014, 38, 856–866. [Google Scholar] [CrossRef]
- Victório, C.P.; Leal-Costa, M.V.; Tavares, E.S.; Kuster, R.M.; Lage, C.L.S. Effects of supplemental UV-A on the development, anatomy and metabolite production of Phyllanthus tenellus cultured in vitro. Photochem. Photobiol. 2011, 87, 685–689. [Google Scholar] [CrossRef]
- Reddy, K.R.; Singh, S.K.; Koti, S.; Kakani, V.G.; Zhao, D.; Gao, W.; Reddy, V.R. Quantifying corn growth and physiological responses to ultraviolet-B radiation for modeling. Agron. J. 2013, 105, 1367–1377. [Google Scholar] [CrossRef]
- Nogués, S.; Allen, D.J.; Morison, J.I.; Baker, N.R. Characterization of stomatal closure caused by ultraviolet-B radiation. Plant Physiol. 1999, 121, 489–496. [Google Scholar] [CrossRef] [Green Version]
- Ranjbarfordoei, A.; Samson, R.; Van Damme, P. Photosynthesis performance in sweet almond [Prunus dulcis (Mill) D. Webb] exposed to supplemental UV-B radiation. Photosynthetica 2011, 49, 107–111. [Google Scholar] [CrossRef] [Green Version]
- Hollósy, F. Effects of ultraviolet radiation on plant cells. Micron 2002, 33, 179–197. [Google Scholar] [CrossRef]
- Vass, I.; Turcsányi, E.; Touloupakis, E.; Ghanotakis, D.; Petrouleas, V. The mechanism of UV-A radiation-induced inhibition of photosystem II electron transport studied by EPR and chlorophyll fluorescence. Biochemistry 2002, 41, 10200–10208. [Google Scholar] [CrossRef]
- Kataria, S.; Guruprasad, K. Intraspecific variations in growth, yield and photosynthesis of sorghum varieties to ambient UV (280–400nm) radiation. Plant Sci. 2012, 196, 85–92. [Google Scholar] [CrossRef]
- Swarna, K.; Bhanumathi, G.; Murthy, S.D.S. Studies on the UV-B radiation induced oxidative damage in thylakoid photofunctions and analysis of the role of antioxidant enzymes in maize primary leaves. Bioscan 2012, 7, 609–610. [Google Scholar]
- Rakocevic, M.; Matsunaga, F.T. Variations in leaf growth parameters within the tree structure of adult Coffea arabica in relation to seasonal growth, water availability and air carbon dioxide concentration. Ann. Bot. 2018, 122, 117–131. [Google Scholar] [CrossRef] [Green Version]
- Ramalho, J.C.; Rodrigues, A.P.; Semedo, J.N.; Pais, I.P.; Martins, L.D.; Simões-Costa, M.C.; Leitão, A.E.; Fortunato, A.S.; Batista-Santos, P.; Palos, I.M.; et al. Sustained photosynthetic performance of Coffea spp. under long-term enhanced [CO2]. PLoS ONE 2013, 8, e82712. [Google Scholar] [CrossRef] [Green Version]
- Hiscox, J.D.; Israelstam, G.F. A method for the extraction of chlorophyll from leaf tissue without maceration. Can. J. Bot. 1979, 57, 1332–1334. [Google Scholar] [CrossRef]
- Wellburn, A.R. The spectral determination of chlorophylls a and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Huang, W.-D.; Lin, K.-H.; Hsu, M.-H.; Huang, M.-Y.; Yang, Z.-W.; Chao, P.-Y.; Yang, C.-M. Eliminating interference by anthocyanin in chlorophyll estimation of sweet potato (Ipomoea batatas L.) leaves. Bot. Stud. 2014, 55, 11. [Google Scholar] [CrossRef] [Green Version]
- Mancinelli, A.L.; Yang, C.-P.H.; Lindquist, P.; Anderson, O.R.; Rabino, I. Photocontrol of anthocyanin synthesis III. The action of streptomycin on the synthesis of chlorophyll and anthocyanin. Plant Physiol. 1975, 55, 251–257. [Google Scholar] [CrossRef] [Green Version]
- Kramer, D.M.; Johnson, G.; Kiirats, O.; Edwards, G.E. New fluorescence parameters for the determination of QA redox state and excitation energy fluxes. Photosynth. Res. 2004, 79, 209–218. [Google Scholar] [CrossRef]
- Kuhlgert, S.; Austic, G.; Zegarac, R.; Osei-Bonsu, I.; Hoh, D.; Chilvers, M.I.; Roth, M.G.; Bi, K.; TerAvest, D.; Weebadde, P.; et al. MultispeQ Beta: A tool for large-scale plant phenotyping connected to the open PhotosynQ network. R. Soc. Open Sci. 2016, 3, 160592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snedecor, G.W.; Cochran, W.G. Statistical Methods, 8th ed.; Iowa State University Press: Ames, IA, USA, 1989. [Google Scholar]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; R Core Team. Nlme: Linear and Nonlinear Mixed Effects Models. R package version 3.1-137; R Foundation: Vienna, Austria, 2018. [Google Scholar]
- Lenth, R.; Singmann, H.; Love, J.; Buerkner, P.; Herve, M. Emmeans: Estimated Marginal Means, aka Least-Squares Means, R Package Version 1.3.4; Available online: https://CRAN.R-project.org/package=emmeans (accessed on 11 November 2020).
- Guruprasad, K.; Bhattacharjee, S.; Kataria, S.; Yadav, S.; Tiwari, A.; Baroniya, S.; Rajiv, A.; Mohanty, P. Growth enhancement of soybean (Glycine max) upon exclusion of UV-B and UV-B/A components of solar radiation: Characterization of photosynthetic parameters in leaves. Photosynth. Res. 2007, 96, 115. [Google Scholar] [CrossRef] [Green Version]
- Kataria, S.; Guruprasad, K. Solar UV-B and UV-A/B exclusion effects on intraspecific variations in crop growth and yield of wheat varieties. Field Crop Res. 2012, 125, 8–13. [Google Scholar] [CrossRef]
- Ferrão, M.A.G.; Ferrão, R.G.; da Fonseca, A.F.; Verdin Filho, A.C.; Volpi, P.S. Origin, geographical dispersion, taxonomy and genetic diversity of Coffea canephora. In Conilon Coffee; Incaper: Vitoria, Brazil, 2019; Chapter 4; pp. 85–109. [Google Scholar]
- Koester, R.P.; Skoneczka, J.A.; Cary, T.R.; Diers, B.W.; Ainsworth, E.A. Historical gains in soybean (Glycine max Merr.) seed yield are driven by linear increases in light interception, energy conversion, and partitioning efficiencies. J. Exp. Bot. 2014, 65, 3311–3321. [Google Scholar] [CrossRef]
- Murali, N.S.; Teramura, A.H. Effectiveness of UV-B radiation on the growth and physiology of field-grown soybean modified by water stress. Photochem. Photobiol. 1986, 44, 215–219. [Google Scholar] [CrossRef]
- Britz, S.J.; Adamse, P. UV-B-induced increase in specific leaf weight of cucumber as a consequence of increased starch content. Photochem. Photobiol. 1994, 60, 116–119. [Google Scholar] [CrossRef]
- John, G.P.; Scoffoni, C.; Buckley, T.N.; Villar, R.; Poorter, H.; Sack, L. The anatomical and compositional basis of leaf mass per area. Ecol. Lett. 2017, 20, 412–425. [Google Scholar] [CrossRef] [Green Version]
- Weraduwage, S.M.; Kim, S.-J.; Renna, L.; Anozie, F.C.; Sharkey, T.D.; Brandizzi, F. Pectin methylesterification impacts the relationship between photosynthesis and plant growth in Arabidopsis thaliana. Plant Physiol. 2016, 171, 833–848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Yang, N.; Xiang, S.; Li, G. Different responses in leaf pigments and leaf mass per area to altitude between evergreen and deciduous woody species. Aust. J. Bot. 2013, 61, 424–435. [Google Scholar] [CrossRef]
- Thakur, D.; Rathore, N.; Chawla, A. Increase in light interception cost and metabolic mass component of leaves are coupled for efficient resource use in the high altitude vegetation. Oikos 2018, 128, 254–263. [Google Scholar] [CrossRef]
- Paoletti, E. UV-B and Mediterranean forest species: Direct effects and ecological consequences. Environ. Pollut. 2005, 137, 372–379. [Google Scholar] [CrossRef] [PubMed]
- Domínguez, E.; Heredia-Guerrero, J.A.; Heredia, A. The biophysical design of plant cuticles: An overview. New Phytol. 2011, 189, 938–949. [Google Scholar] [CrossRef]
- Krauss, P.; Markstadter, C.; Riederer, M. Attenuation of UV radiation by plant cuticles from woody species. Plant Cell Environ. 1997, 20, 1079–1085. [Google Scholar] [CrossRef]
- Kolb, C.A.; Pfundel, E.E. Origins of non-linear and dissimilar relationships between epidermal UV absorbance and UV absorbance of extracted phenolics in leaves of grapevine and barley. Plant Cell Environ. 2005, 28, 580–590. [Google Scholar] [CrossRef]
- Peng, Q.; Zhou, Q. Antioxidant capacity of flavonoid in soybean seedlings under the joint actions of rare earth element La(III) and ultraviolet-B stress. Biol. Trace Elem. Res. 2008, 127, 69–80. [Google Scholar] [CrossRef]
- Rozema, J.; Chardonnens, A.; Tosserams, M.; Hafkenscheid, R.; Bruijnzeel, S.; Gieskes, W.W.C.; Geijn, S.C.; Nolan, C.; Boois, H. Leaf thickness and UV-B absorbing pigments of plants in relation to an elevational gradient along the Blue Mountains, Jamaica. In UV-B and Biosphere; Springer: Dordrecht, The Netherlands, 1997; pp. 150–159. [Google Scholar]
- Day, T.A.; Martin, G.; Vogelmann, T.C. Penetration of UV-B radiation in foliage: Evidence that the epidermis behaves as a non-uniform filter. Plant Cell Environ. 1993, 16, 735–741. [Google Scholar] [CrossRef]
- Bilger, W.; Veit, M.; Schreiber, L.; Schreiber, U. Measurement of leaf epidermal transmittance of UV radiation by chlorophyll fluorescence. Physiol. Plant. 1997, 101, 754–763. [Google Scholar] [CrossRef]
- Barnes, P.W.; Kersting, A.R.; Flint, S.D.; Beyschlag, W.; Ryel, R.J. Adjustments in epidermal UV-transmittance of leaves in sun-shade transitions. Physiol. Plant. 2013, 149, 200–213. [Google Scholar] [CrossRef] [PubMed]
- Nybakken, L.; Bilger, W.; Johanson, U.; Zielke, M. Epidermal UV-screening in vascular plants from Svalbard (Norwegian Arctic). Polar Biol. 2004, 27, 383–390. [Google Scholar] [CrossRef]
- Caldwell, M.M.; Robberecht, R.; Flint, S.D. Internal filters: Prospects for UV-acclimation in higher plants. Physiol. Plant. 1983, 58, 445–450. [Google Scholar] [CrossRef]
- Jones, H.G. Stomatal control of photosynthesis and transpiration. J. Exp. Bot. 1998, 1, 387–398. [Google Scholar] [CrossRef]
- Dai, Q.; Chavez, A.Q.; Vergara, B.S.; Peng, S. Effects of UVB radiation on stomatal density and opening in rice (Oryza sativa L.). Ann. Bot. 1995, 76, 65–70. [Google Scholar] [CrossRef]
- Gitz, D.C.; Liu-Gitz, L.; Britz, S.J.; Sullivan, J.H. Ultraviolet-B effects on stomatal density, water-use efficiency, and stable carbon isotope discrimination in four glasshouse-grown soybean (Glyicine max) cultivars. Environ. Exp. Bot. 2005, 53, 343–355. [Google Scholar] [CrossRef]
- Morais, H.; Medri, M.E.; Marur, C.J.; Caramori, P.H.; Ribeiro, A.M.D.A.; Gomes, J.C. Modifications on leaf anatomy of Coffea arabica caused by shade of Pigeonpea (Cajanus cajan). Braz. Arch. Biol. Technol. 2004, 47, 863–871. [Google Scholar] [CrossRef] [Green Version]
- Dow, G.J.; Bergmann, D.C.; Berry, J.A. An integrated model of stomatal development and leaf physiology. New Phytol. 2013, 201, 1218–1226. [Google Scholar] [CrossRef] [PubMed]
- Ren, T.; Weraduwage, S.M.; Sharkey, T.D. Prospects for enhancing leaf photosynthetic capacity by manipulating mesophyll cell morphology. J. Exp. Bot. 2018, 70, 1153–1165. [Google Scholar] [CrossRef] [PubMed]
- Hoober, J.K.; Eggink, L.L.; Chen, M. Chlorophylls, ligands and assembly of light-harvesting complexes in chloroplasts. Photosynth. Res. 2007, 94, 387–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, R.; Tanaka, A. Chlorophyll cycle regulates the construction and destruction of the light-harvesting complexes. Biochim. Biophys. Acta Bioenerg. 2011, 1807, 968–976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, A.; Yamamoto, Y.; Tsuji, H. Formation of chlorophyll-protein complexes during Greening. 2. Redistribution of chlorophyll among apoproteins. Plant Cell Physiol. 1991, 32, 195–204. [Google Scholar] [CrossRef]
- Nunes, M.A.; Ramalho, J.D.C.; Dias, M.A. Effect of nitrogen supply on the photosynthetic performance of leaves from coffee plants exposed to bright light. J. Exp. Bot. 1993, 44, 893–899. [Google Scholar] [CrossRef]
- Agrawal, S.; Rathore, D. Changes in oxidative stress defense system in wheat (Triticum aestivum L.) and mung bean (Vigna radiata L.) cultivars grown with and without mineral nutrients and irradiated by supplemental ultraviolet-B. Environ. Exp. Bot. 2007, 59, 21–33. [Google Scholar] [CrossRef]
- Lidon, F.J.C.; Teixeira, M.; Ramalho, J.C. Decay of the chloroplast pool of ascorbate switches on the oxidative burst in UV-B-irradiated rice. J. Agron. Crop Sci. 2011, 198, 130–144. [Google Scholar] [CrossRef]
- Müller, P.; Li, X.-P.; Niyogi, K.K. Non-photochemical quenching. a response to excess light energy. Plant Physiol. 2001, 125, 1558–1566. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | C. arabica | C. canephora | p-Value * | ||||
|---|---|---|---|---|---|---|---|
| Environment | UVam | UVre | UVam | UVre | Environment | Species | Environment: Species |
| Parameters | Plant Growth Traits | ||||||
| Plant height (cm) | 32.3 ± 1.52 | 32.4 ± 1.58 | 23.0 ± 1.52 | 23.1 ± 1.52 | 0.9280 | <0.0001 | 0.7247 |
| Basal stem diameter (mm) | 12.6 ± 0.62 | 13.9 ± 0.65 | 11.4 ± 0.62 | 12.7 ± 0.62 | 0.0647 | 0.0723 | 0.3961 |
| Total number of leaves | 148.0 ± 5.58 | 153.0 ± 5.56 | 124.0 ± 5.58 | 156.0 ± 5.58 | 0.0033 | 0.0629 | 0.0235 |
| Leaf area (m2) | 0.58 ± 0.03 | 0.61 ± 0.03 | 0.47 ± 0.03 | 0.67 ± 0.02 | 0.0006 | 0.3345 | 0.0149 |
| Leaf dry mass (g) | 54.3 ± 2.45 | 47.8 ± 2.61 | 45.4 ± 2.45 | 61.6 ± 2.45 | 0.0454 | 0.4046 | 0.0002 |
| Stem dry mass (g) | 31.9 ± 1.65 | 39.9 ± 1.71 | 29.9 ± 1.65 | 37.9 ± 1.65 | <0.0001 | 0.1925 | 0.2338 |
| Root dry mass (g) | 40.3 ± 4.61 | 43.70 ± 4.92 | 48.8 ± 4.61 | 70.1 ± 4.61 | 0.0060 | 0.0008 | 0.0520 |
| Total dry mass (g) | 128.2 ± 6.35 | 129.1 ± 6.35 | 122.5 ± 6.35 | 171.2 ± 6.35 | 0.9083 | 0.4965 | 0.0005 |
| SLM (g m−2) | 84.5 ± 2.96 | 74.7 ± 3.09 | 92.5 ± 2.96 | 82.6 ± 2.96 | 0.0122 | 0.0303 | 0.3791 |
| Leaf anatomy | |||||||
| Abaxial cuticle (µm, 40×) | 3.45 ± 0.17 | 2.72 ± 0.17 | 3.29 ± 0.17 | 2.56 ± 0.17 | 0.0003 | 0.2870 | 0.2917 |
| Adaxial cuticle (µm, 40×) | 4.31 ± 0.14 | 2.70 ± 0.14 | 3.73 ± 0.14 | 2.90 ± 0.14 | <0.0001 | 0.2069 | 0.0149 |
| Abaxial epidermis (µm, 40×) | 17.2 ± 0.87 | 13.9 ± 0.87 | 13.1 ± 0.87 | 13.0 ± 0.87 | 0.0173 | 0.0050 | 0.0781 |
| Adaxial epidermis (µm, 40×) | 21.8 ± 0.94 | 21.3 ± 0. 94 | 20.3 ± 0. 94 | 19.8 ± 0. 94 | 0.5222 | 0.0548 | 0.1939 |
| Palisade parenchyma (µm, 20×) | 57.1 ± 2.97 | 55.6 ± 2.97 | 63.4 ± 2.97 | 66.0 ± 2.97 | 0.8523 | 0.0162 | 0.5138 |
| Spongy parenchyma (µm, 20×) | 164. 0 ± 7.12 | 161.0 ± 7.12 | 138.0 ± 7.12 | 135.0 ± 7.12 | 0.6692 | 0.0049 | 0.1013 |
| Stomatal density (number mm−2) | 192.0 ± 11.7 | 200.0 ± 12.5 | 320.0 ± 11.7 | 291.0 ± 11.7 | 0.5687 | <0.0001 | 0.0884 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bernado, W.d.P.; Rakocevic, M.; Santos, A.R.; Ruas, K.F.; Baroni, D.F.; Abraham, A.C.; Pireda, S.; Oliveira, D.d.S.; Cunha, M.D.; Ramalho, J.C.; et al. Biomass and Leaf Acclimations to Ultraviolet Solar Radiation in Juvenile Plants of Coffea arabica and C. canephora. Plants 2021, 10, 640. https://doi.org/10.3390/plants10040640
Bernado WdP, Rakocevic M, Santos AR, Ruas KF, Baroni DF, Abraham AC, Pireda S, Oliveira DdS, Cunha MD, Ramalho JC, et al. Biomass and Leaf Acclimations to Ultraviolet Solar Radiation in Juvenile Plants of Coffea arabica and C. canephora. Plants. 2021; 10(4):640. https://doi.org/10.3390/plants10040640
Chicago/Turabian StyleBernado, Wallace de Paula, Miroslava Rakocevic, Anne Reis Santos, Katherine Fraga Ruas, Danilo Força Baroni, Ana Cabrera Abraham, Saulo Pireda, Dhiego da Silva Oliveira, Maura Da Cunha, José Cochicho Ramalho, and et al. 2021. "Biomass and Leaf Acclimations to Ultraviolet Solar Radiation in Juvenile Plants of Coffea arabica and C. canephora" Plants 10, no. 4: 640. https://doi.org/10.3390/plants10040640