
Impact of Germination on the Microstructural and Physicochemical Properties of Different Legume Types




Abstract
:1. Introduction
2. Results
2.1. Appearance of Legume Seeds during Germination Period
2.2. SEM Analysis
2.3. Physical-Chemical Characterization of Legume Seeds during the Germination Period
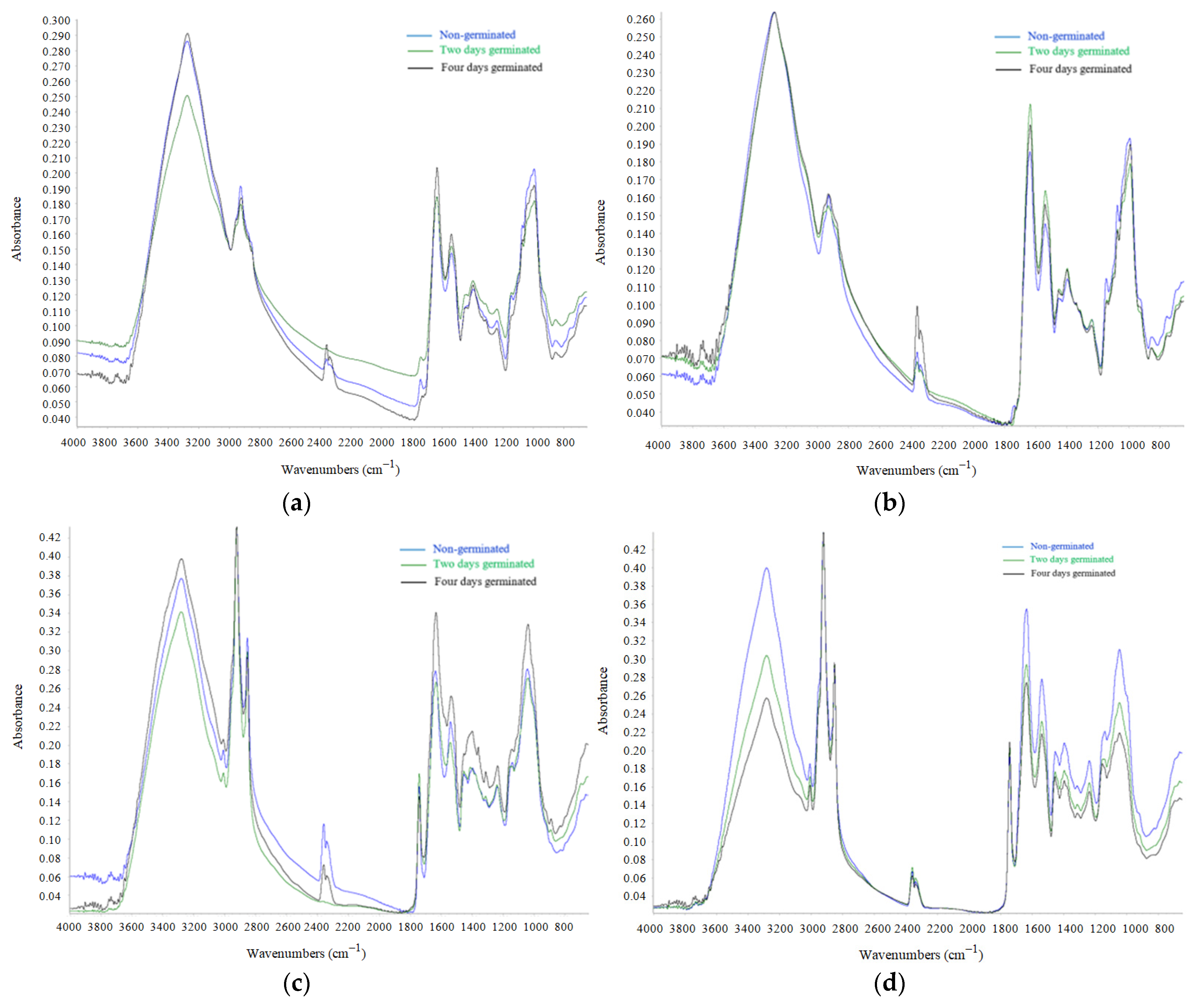
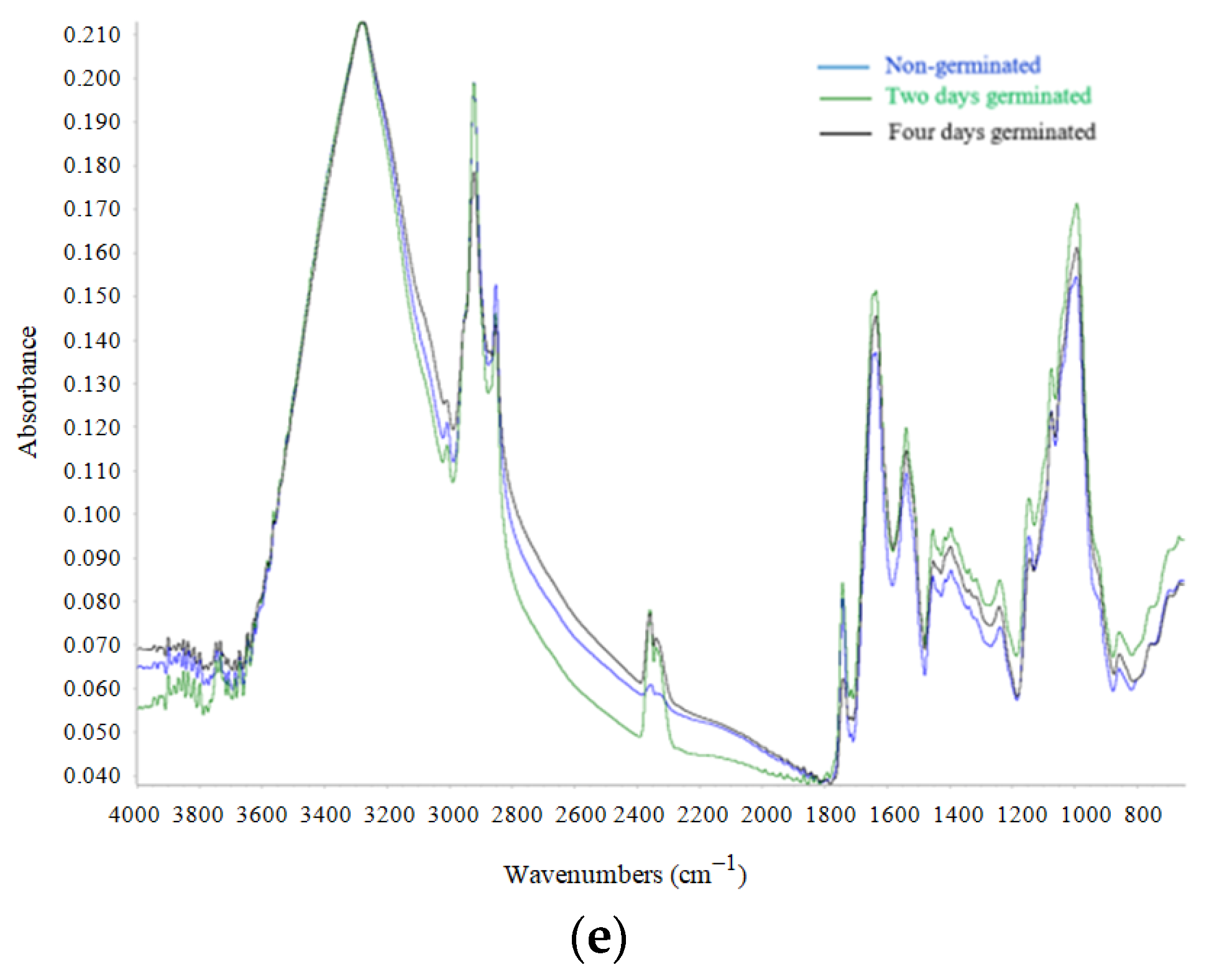
2.4. FT-IR Analysis
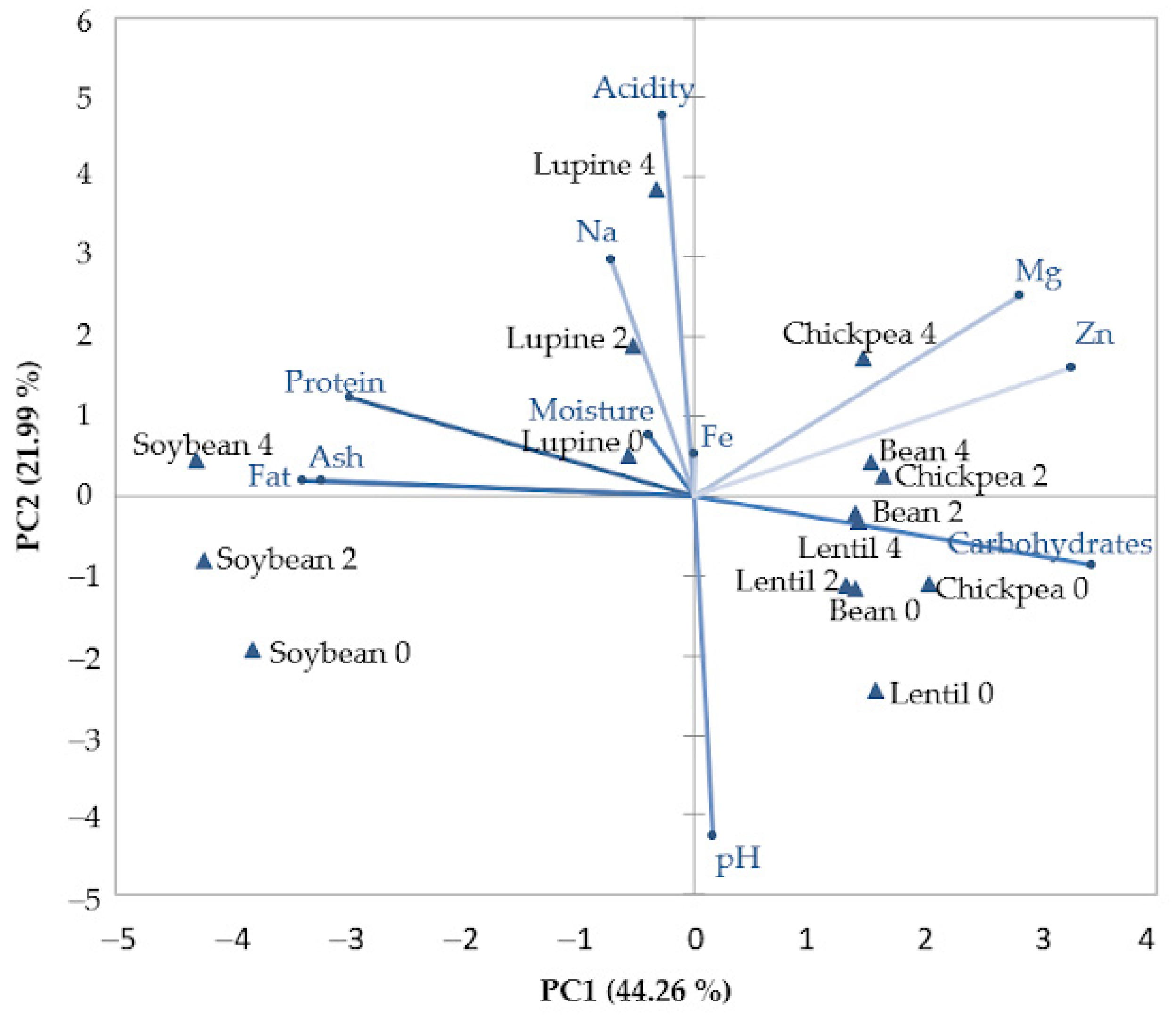
2.5. Relationships between Physico-Chemical Values of Legume Seeds during the Germination Period
3. Discussion
3.1. Appearance of Legume Seeds during Germination Period
3.2. SEM Analysis
3.3. Physical-Chemical Characterization of Legume Seeds during the Germination Period
3.4. FT-IR Analysis
3.5. Relationships between Physico-Chemical Values of Legume Seeds during the Germination Period
4. Materials and Methods
4.1. Materials
4.2. Germination and Lyophilization of Legume Seeds
4.3. Appearance of Legume Seeds Analysis
4.4. SEM Analysis
4.5. Physico-Chemical Composition Analysis
4.6. FT-IR Analysis
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Laleg, K.; Cassan, D.; Abecassis, J.; Micard, V. Procédé de Fabrication de Pâte Destinée a L’alimentation Humaine et/ou Animale Comprenant au Moins 35% de Legumineuse. Patent No. WO 2016097328 A1, 23 June 2016. [Google Scholar]
- Eker, M.E.; Karakaza, S. Influence of the Addition of Chia Seeds and Germinated Seeds and Sprouts on the Nutritional and Beneficial Properties of Yogurt. Int. J. Gastron. Food Sci. 2020, 22, 100276. [Google Scholar] [CrossRef]
- Benmeziane, F.; Raigar, R.K.; Ayat, N.E.H.; Aoufi, D.; Djermoune-Arkoub, L.; Chala, A. Lentil (Lens culinaris) Flour Addition to Yogurt: Impact on Physicochemical, Microbiological and Sensory Attributes during Refrigeration Storage and Microstructure changes. LWT 2021, 140, 110793. [Google Scholar] [CrossRef]
- Martins, Z.E.; Pinho, O.; Ferreira, I.M.P.L.V.O. Food Industry By-products Used as Functional Ingredients of Bakery Products. Trends Food Sci. Technol. 2017, 67, 106–128. [Google Scholar] [CrossRef]
- Mitelut, A.C.; Popa, E.E.; Popescu, P.A.; Popa, M.A. Chapter 7–Trends of Innovation in Bread and Bakery Production. In Trends in Wheat and Bread Making; Academic Press: Cambridge, MA, USA, 2021; pp. 199–226. [Google Scholar]
- Mridula, D.; Sharma, M. Development of Non-dairy Probiotic Drink Utilizing Sporuted Cereals, Legume and Soymilk. LWT 2015, 62, 482–487. [Google Scholar] [CrossRef]
- Ben-Porat, T.; Weiss, R.; Sherf-Dagan, S.; Nabulsi, N.; Maayani, A.; Khalaileh, A.; Abed, S.; Brodie, R.; Harari, R.; Mintz, Y.; et al. Nutritional Deficiencies in Patients with Severe Obesity before Bariatric Surgery: What Should Be the Focus During the Preoperative Assessment? J. Acad. Nutr. Diet 2020, 120, 874–884. [Google Scholar] [CrossRef]
- McLoughlin, R.J.; McKie, K.; Hirsh, M.P.; Cleary, M.A.; Aidlen, J.T. Impact of Nutritional Deficiencies on Children and Young Adults with Crohn’s Disease Undergoing Intraabdominal Surgery. J. Pediatr. Surg. 2020, 55, 1556–1561. [Google Scholar] [CrossRef] [PubMed]
- Zule, S.; Wanik, J.; Holm, E.M.; Bruder, M.B.; Shanley, E.; Sherman, C.Q.; Fitterman, M.; Lerner, J.; Marcello, M.; Parenchuck, N.; et al. Nutritional Deficiency Disease Secondary to ARFID Symptoms Associated with Autism and the Broad Autism Phenotype: A Qualitative Systematic Review of Case Reports and Case Series. J. Acad. Nutr. Diet. 2021, 121, 467–492. [Google Scholar]
- Carubbi, F.; Barbato, A.; Burlina, A.B.; Francini, F.; Mignani, R.; Pegoraro, E.; Landini, L.; De Danieli, G.; Bruni, S.; Strayyullo, P. Nutrition in Adult Patients with Selected Lysosomal Storage Diseases. Nutr. Metab. Cardiovasc. Dis. 2020, 31, 733–744. [Google Scholar] [CrossRef]
- Wang, H.; Liu, F.; Ma, H.; Zin, H.; Wang, P.; Bai, B.; Guo, L.; Geng, Q. Associations between Depression, Nutrition, and Outcomes among Individuals with Coronary Artery Disease. Nutrition 2021, 86, 111157. [Google Scholar] [CrossRef]
- Rajakumari, R.; Oluwafemi, O.S.; Thomas, S.; Kalarikkal, N. Dietary Supplements Containing Vitamins and Minerals: Formulation, Optimization and Evaluation. Powder Technol. 2018, 336, 481–492. [Google Scholar] [CrossRef]
- Scaife, J.C.; Godier, L.R.; Reinecke, A.; Harmer, C.J.; Park, R.J. Differential Activation of the Frontal Pole to High vs. Low Calorie Foods: The Neural Basis of Food Preference in Anorexia Nervosa? Psychiatry Res. Neuroimaging 2016, 258, 44–53. [Google Scholar] [CrossRef]
- de Vries, R.; de Vet, E.; de Graaf, K.; Boesveldt, S. Foraging Minds in Modern Environments: High-calorie and Savory-taste Biases in Human Food Spatial Memory. Appetite 2020, 152, 104718. [Google Scholar] [CrossRef]
- Lee, M.; Lee, J.H. Automatic Attentional Bias Toward High-calorie Food Cues and Body Shape Concerns in Individuals with a High Level of Weight Suppression: Preliminary Findings. Eat. Behav. 2021, 40, 101471. [Google Scholar] [CrossRef]
- Racine, S.; Suissa-Rocheleau, L.; Martin, S.J.; Benning, S.D. Implicit and Explicit Motivational Responses to High- and Low-calorie Food in Women with Disordered Eating. Int. J. Psychophysiol. 2021, 159, 37–46. [Google Scholar] [CrossRef]
- Kan, L.; Nie, S.; Hu, J.; Wang, S.; Bai, Z.; Wang, J.; Zhou, Y.; Jiang, J.; Zeng, Q.; Song, K. Comparative Study on the Chemical Composition, Anthocyanins, Tocopherols and Carotenoids of Selected Legumes. Food Chem. 2018, 260, 317–326. [Google Scholar] [CrossRef]
- Kumar, S.; Pandey, G. Biofortification of Pulses and Legumes to Enhance Nutrition. Heliyon 2020, 6, e03682. [Google Scholar] [CrossRef] [PubMed]
- Oreno-Valdespino, C.A.; Luna-Vital, D.; Camacho-Ruiz, R.M.; Mojica, L. Bioactive Proteins and Phytochemicals from Legumes: Mechanisms of Action Preventing Obesity and Type-2 diabetes. Food Res. Int. 2020, 130, 108905. [Google Scholar] [CrossRef]
- Liu, W.; Dehghan, M.; Mente, A.; Wang, C.; Yan, R.; Rangarajan, S.; Tse, L.A.; Yusuf, S.; Liu, X.; Wang, Y.; et al. Fruit, Vegetable, and Legume Intake and the Risk of All-cause, Cardiovascular, and Cancer Mortality: A Prospective Study. Clin. Nutr. 2021. [Google Scholar] [CrossRef] [PubMed]
- Hefnawy, T.H. Effect of Processing Methods on Nutritional Composition and Anti-nutritional Factors in Lentils (Lens culinaris). Ann. Agric. Sci. 2011, 56, 57–61. [Google Scholar] [CrossRef] [Green Version]
- Rui, S.; Hua, W.; Rui, G.; Qin, L.; Lei, P.; Jianan, L.; Zhihui, H.; Chanyou, C. The Diversity of Four Anti-nutritional Factors in Common Bean. Hortic. Plant J. 2016, 2, 97–104. [Google Scholar]
- Rahate, K.A.; Madhumita, M.; Pradhakar, P.K. Nutritional Composition, Anti-nutritional Factors, Pretreatments-cum-processing Impact and Food Formulation Potential of Faba Bean (Vicia faba L.): A Comprehensive Review. LWT 2021, 138, 110796. [Google Scholar] [CrossRef]
- Swieca, M.; Gawlik-Dziki, U.; Jakubczyk, A.; Bochnak, J.; Sikora, M.; Suliburska, J. Nutritional Quality of Fresh and Stored Legumes Sprouts—Effect of Lactobacillus plantarum 299v Enrichment. Food Chem. 2019, 288, 325–332. [Google Scholar] [CrossRef]
- Ohanenye, I.C.; Tsopmo, A.; Ejike, C.E.C.C.; Udenigwe, C.C. Germination as a Bioprocess for Enhancing the Quality and Nutritional Prospects of Legume Proteins. Trends Food Sci. Technol. 2020, 101, 213–222. [Google Scholar] [CrossRef]
- Chavan, M.; Gat, Y.; Harmalkar, M.; Waghmare, R. Development of Non-dairy Fermented Probiotic Drink Based on Germinated and Ungerminated Cereals and Legume. LWT 2018, 91, 339–344. [Google Scholar] [CrossRef]
- Kaczmarska, K.T.; Chandra-Hioe, M.V.; Frank, D.; Arcot, J. Enhancing Wheat Muffin Aroma through Addition of Germinated and Fermented Australian Sweet Lupin (Lupinus angustifolius L.) and Soybean (Glycine max L.) Flour. LWT 2018, 96, 205–214. [Google Scholar] [CrossRef]
- Ujiroghene, O.J.; Liu, L.; Zhang, S.; Lu, J.; Zhang, C.; Lv, J.; Pang, X.; Zhang, M. Antioxidant Capacity of Germinated Quinoa-based Yoghurt and Concomitant Effect of Sprouting on Its Functional Properties. LWT 2019, 116, 108592. [Google Scholar] [CrossRef]
- Torres, A.; Frias, J.; Granito, M.; Vidal-Valverde, C. Germinated Cajanus cajan Seeds as Ingredients in Pasta Products: Chemical, Biological and Sensory Evaluation. Food Chem. 2007, 101, 202–211. [Google Scholar] [CrossRef]
- Polat, H.; Capar, T.D.; Inanir, C.; Ekici, L.; Yalcin, H. Formulation of Functional Crackers Enriched with Germinated Lentil extract: A Response Surface Methodology Box-Behnken Design. LWT 2020, 123, 109065. [Google Scholar] [CrossRef]
- López-Martínez, L.X.; Leyva-López, N.; Gutiérrez-Grijalva, E.P.; Heredia, J.B. Effect of Cooking and Germination on Bioactive Compounds in Pulses and Their Health Benefits. J. Funct. Foods 2017, 38, 624–634. [Google Scholar] [CrossRef]
- Xu, M.; Jin, Z.; Peckrul, A.; Chen, B. Pulse Seed Germination Improves Antioxidative Activity of Phenolic Compounds in Stripped Soybean Oil-in-water Emulsions. Food Chem. 2018, 250, 140–147. [Google Scholar] [CrossRef]
- Miano, A.C.; Garcia, J.A.; Augusto, P.E.D. Correlation between Morphology, Hydration Kinetics and Mathematical Models on Andean Lupin (Lupinusmutabilis Sweet) Grains. LWT Food Sci. Techol. 2015, 290–298. [Google Scholar] [CrossRef]
- Ma, F.; Cholewa, E.; Mohamed, T.; Peterson, C.A.; Gijzen, M. Cracks in the Palisade Cuticle of Soybean Seed Coats Correlate with Their Permeability to Water. Ann. Bot. 2004, 94, 213–228. [Google Scholar] [CrossRef] [Green Version]
- Marconi, E.; Ruggeri, S.; Cappelloni, M.; Leonardi, D.; Carnovale, E. Physicochemical, Nutritional, and Microstructural Characteristics of Chickpeas (Cicerarietinum L.) and Common Beans (Phaseolus vulgaris L.) Following Microwave Cooking. J. Agric. Food Chem. 2000, 48, 598–694. [Google Scholar] [CrossRef]
- Biaszczak, W.; Doblado, R.; Frias, J.; Vidal-Valverde, C.; Sadowska, J.; Fornal, J. Microstructural and Biochemical Changes in Raw and Germinatedcowpea Seeds upon High-pressure Treatment. Food Res. Int. 2007, 40, 415–423. [Google Scholar] [CrossRef]
- Swanson, B.; Hughes, J.S.; Rasmussen, H.P. Seed Microstructure: Review of Water Imbibition in Legumes. Food Struct. 1985, 4, 115–124. [Google Scholar]
- Atudorei, D.; Stroe, S.G.; Codină, G.G. Physical, Physiological and Minerals Changes of Different Legumes Types during the Germination Process. Food Technol. 2020, 9, 844–863. [Google Scholar]
- Borek, S.; Ratajczack, W.; Ratajczack, L. Regulation of Storage Lipid Metabolism in Developing and Germinating Lupin (Lupinus spp.) Seeds. Acta Physiol. Plant 2015, 37, 119. [Google Scholar] [CrossRef] [Green Version]
- Atudorei, D.; Codină, G.G. Perspectives on the Use of Germinated Legumes in the Bread Making Process, A Review. Appl. Sci. 2020, 10, 6244. [Google Scholar] [CrossRef]
- Kassegn, H.H.; Atsbha, T.W.; Weldeabezgi, L.T.; Yildiz, F. Effect of Germination Process on Nutrients and Phytochemicals Contents of Faba Bean (Viciafaba L.) for Weaning Food Preparation. Cogent Food Agric. 2018, 4. [Google Scholar] [CrossRef]
- Huang, P.V.; Yen, N.T.H.; Phi, N.T.L.; Tien, N.P.H.; Trung, N.T.T. Nutritional Composition, Enzyme Activities and Bioactive Compounds Ofmung Bean (Vignaradiata L.) Germinated under Dark and Light Conditions. LWT Food Sci. Techol. 2020, 133, 110100. [Google Scholar] [CrossRef]
- Chinma, C.E.; Adedeji, O.E.; Etim, I.I.; Aniaka, G.I.; Mathew, E.O.; Ekeh, U.B.; Anumba, N.L. Physicochemical, Nutritional, and Sensory Properties of Chips Produced from Germinated African Yam Bean (Sphenostylisstenocarpa). LWT Food Sci. Techol. 2021, 136, 110330. [Google Scholar] [CrossRef]
- Domínguez-Arispuro, D.M.; Cuevas-Rodríguez, E.O.; Milán-Carrillo, J.; León-López, L.; Gutiérrez-Dorado, R.; Reyes-Moreno, C. Optimal Germination Condition Impacts on the Antioxidant Activity and Phenolic Acids Profile in Pigmented Desi Chickpea (Cicerarietinum L.) Seeds. J. Food. Sci. Technol. 2018, 55, 638–647. [Google Scholar] [CrossRef]
- Xu, M.; Jin, Z.; Simsek, S.; Hall, C.; Rao, J.; Chen, B. Effect of Germination on the Chemical Composition, Thermal, Pasting, and Moisture Sorption Properties of Flours from Chickpea, Lentil, and Yellow Pea. Food Chem. 2019, 295, 579–587. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, C.D.; Bubolz, V.K.; da Silva, J.; Dittgen, C.L.; Ziegler, V.; de Oliveira Raphaelli, C.; de Oliveira, M. Canges in the Chemical Composition and Bioactive Compounds of Chickpea (Cicer arietinum L.) Fortified by Germination. LWT 2019, 111, 363–369. [Google Scholar] [CrossRef]
- Kayembe, N.C.; Rensburg, J. Germination as a Processing Technique for Soybeans in Small-scale Farming. S. Afr. J. Anim. Sci. 2013, 42, 167–173. [Google Scholar] [CrossRef] [Green Version]
- Upadhyay, N.; Jaiswal, P.; Jha, S.N. Application of Attenuated Total Reflectance Fourier Transform Infrared Spectroscopy (ATR–FTIR) in MIR Range Coupled with Chemometrics for Detection of Pig Body Fat in Pure Ghee (Heat Clarified Milk Fat). J. Mol. Struct. 2018, 1153, 275–281. [Google Scholar] [CrossRef]
- Jaiswal, P.; Jha, S.N.; Kaur, J.; Borah, A.; Ramya, H.G. Detection of Aflatoxin M1 in Milk Using Spectroscopy and Multivariate Analyses. Food Chem. 2018, 238, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Andrade, J.; Pereira, C.G.; de Almeida Junior, J.C.; Viana, C.C.R.; de Oliveira Neves, L.N.; da Silva, P.H.F.; Bell, M.J.V.; de Calvalho dos Anjos, V. FTIR-ATR Determination of Protein Content to Evaluate Whey Protein Concentrate Adulteration. LWT 2019, 99, 166–172. [Google Scholar] [CrossRef]
- Fazio, E.; Gualandi, G.; Palleschi, S.; Footitt, S.; Silvestroni, L. Optically Functionalized Biomorphism of Bean Seeds. J. Lumin. 2017, 182, 189–195. [Google Scholar] [CrossRef]
- Cheng, Y.D.; Bai, Y.X.; Jia, M.; Chen, Y.; Wang, D.; Wu, T.; Wang, G.; Yang, H.W. Potential Risks of Nicotine on the Germination, Growth, and Nutritional Properties of Broad Bean. Ecotoxicology 2021, 209, 111797. [Google Scholar]
- Elezz, A.A.; Ahmed, T. The Efficacy Data of Two Household Cleaning and Disinfecting Agents on Lens Culinaris Medik and Vicia Faba Seed Germination. Data Brief 2021, 35, 106811. [Google Scholar] [CrossRef] [PubMed]
- Cruz, F.; Batista-Santos, P.; Monteiro, S.; Neves-Martins, J.; Ferreira, R.B. Maximizing Blad-containing Oligomer Fungicidal Activity in Sweet Cultivars of Lupinus Albus Seeds. Ind. Crops Prod. 2021, 162, 113242. [Google Scholar] [CrossRef]
- Kim, S.L.; Lee, J.E.; Kwon, Y.U.; Kim, W.H.; Jung, G.H.; Kim, D.W.; Lee, C.K.; Lee, Y.Y.; Kim, M.J.; Kim, Y.H.; et al. The Germination of Soybeans Increases the Water-soluble Components and Could Generate Innovations in Soy-based Foods. Food Chem. 2013, 136, 491–500. [Google Scholar] [CrossRef] [PubMed]
- Bueno, D.B.; da Silva Júnior, S.I.; Chiarotto, A.B.S.; Cardoso, T.M.; Neto, J.A.; dos Reis, G.C.; Glória, M.B.A.; Tavano, O.L. The Germination of Soybeans Increases the Water-soluble Components and Could Generate Innovations in Soy-based Foods. LWT 2020, 117, 108599. [Google Scholar] [CrossRef]
- Baudoin, J.P.; Maquet, A. Improvement of Protein and Amino Acids Content in Seeds of Food Legumes. A Case Study in Phaseolus. Biotechnol. Agron. Soc. Environ. 1999, 3, 220–224. [Google Scholar]
- Zhao, M.; Zhang, H.; Yan, H.; Qiu, L.; Baskin, C.C. Mobilization and Role of Starch, Protein, and Fat Reserves During Seed Germination of Six Wild Grassland Species. Front. Plant Sci. 2018, 9, 234. [Google Scholar] [CrossRef] [Green Version]
- Ulrichs, T.; Drotleff, A.M.; Ternes, W. Determination of Heat-induced Changes in the Protein Secondary Structure of Reconstituted Livetins (Water-soluble Proteins from Hen’s Egg Yolk) by FTIR. Food Chem. 2015, 172, 909–920. [Google Scholar] [CrossRef]
- Harwood, J.L. Lipid Synthesis by Germinating Soya Bean. Phytochemistry 1975, 14, 1985–1990. [Google Scholar] [CrossRef]
- Inyang, C.; Zakari, U.M. Effect of Germination and Fermentation of Pearl Millet on Proximate, Chemical and Sensory Properties of Instant “Fura”-A Nigerian Cereal Food. Pak. J. Nutr. 2008, 7, 9–12. [Google Scholar] [CrossRef]
- Otutu, O.L.; Ikuomola, D.S.; Oloruntoba, R.O. Effect of Sprouting Days on the Chemical and Physicochemical Properties of Maize Starch. J. Food Nutr. Res. 2014, 2, 131–149. [Google Scholar]
- Chinma, C.E.; Adewuyi, O.; Abu, J.O. Effect of Germination on the Chemical, Functional and Pasting Properties of Flour from Brown and Yellow Varieties of Tigernut (Cyperus esculentus). Food Res. Int. 2009, 42, 1004–1009. [Google Scholar] [CrossRef]
- Echendu, C.A.; Obizoba, I.C.; Aniyka, J.U. Effects of Germination on Chemical Composition of Ground Bean (Kerstingiellageocarpa harm) Seeds. Pak. J. Nutr. 2009, 8, 1849–1854. [Google Scholar] [CrossRef] [Green Version]
- Cornejo, F.; Novillo, G.; Villacrés, E.; Rosell, C.M. Evaluation of the Physicochemical and Nutritional Changes in Two Amaranth Species (Amaranthus quitensis and Amaranthus caudatus) after Germination. Food Res. Int. 2019, 121, 933–939. [Google Scholar] [CrossRef] [PubMed]
- Chinma, C.E.; Anuonze, J.C.; Simon, O.C.; Ahiare, R.O.; Danbaba, N. Effect of Germination on the Physicochemical and Antioxidant Characteristics of Rice Flour from Three Rice Varieties from Nigeria. Fodd. Chem. 2015, 185, 454–458. [Google Scholar] [CrossRef] [PubMed]
- Maetens, E.; Hettiarachchy, N.; Dewettinck, K.; Horax, R.; Meons, K.; Moseley, D.O. Physicochemical and Nutritional Properties of a Healthy Snack Chip Developed from Germinated Soybeans. LWT 2017, 84, 505–510. [Google Scholar] [CrossRef]
- Nkhata, S.G.; Ayua, E.; Kamau, E.H.; Shingiro, J.B. Fermentation and Germination Improve Nutritional Value of Cereals and Legumes through Activation of Endogenous Enzymes. Food Sci. Nutr. 2018, 6, 2446–2458. [Google Scholar] [CrossRef] [Green Version]
- Sokrab, A.M.; Ahmed, I.A.M.; Babiker, E.E. Effect of Germination on Antinutritional Factors, Total, and Extractable Minerals of High and Low Phytate Corn (Zea mays L.) Genotypes. J. Saudi Soc. Agric. Sci. 2012, 11, 123–128. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.; Xie, W.; Jin, X.; Wang, Q.; He, Y. Effects of Germination on Iron, Zinc, Calcium, Manganese, and Copper Availability from Cereals and Legumes. CYTA J. Food 2014, 12, 22–26. [Google Scholar] [CrossRef] [Green Version]
- El-Adawy, T.A.; Rahma, E.H.; El-Bedawey, A.A.; El-Beltagy, A.E. Nutritional Potential and Functional Properties of Germinated Mung Bean, Pea and Lentil Seeds. Plants. Food Hum. Nutr. 2003, 58, 1–13. [Google Scholar] [CrossRef]
- Yasmin, A.; Zeb, A.; Khalil, A.W.; Paracha, G.M. Effect of Processing on Anti-nutritional Factors of Red Kidney Bean (Phaseolus vulgaris) Grains. Food Bioproc. Tech. 2008, 1, 415–419. [Google Scholar] [CrossRef]
- Ogbonna, A.C.; Abuajah, C.I.; Udofia, U.S. Effect of Malting Conditions on the Nutritional and Anti-nutritional Factors of Sorghum Grist. Ann. Univ. Dunarea Jos Galati 2012, 36, 64–72. [Google Scholar]
- Chinma, C.E.; Abu, J.O.; Asikwe, B.N.; Sunday, T.; Adebo, O.A. Effect of Germination on the Physicochemical, Nutritional, Functional, Thermal Properties and in vitro Digestibility of Bambara Groundnut Flours. LWT 2021, 140, 110749. [Google Scholar] [CrossRef]
- Kajla, P.; Sharma, A.; Sood, D.R. Effect of Germination on Proximate Principles, Minerals and Antinutrients of Flaxseeds. J. Dairy Res. 2017, 36, 52–57. [Google Scholar] [CrossRef] [Green Version]
- Andrade, J.; Pereira, C.G.; Ranquine, T.; Azarias, C.A.; Bell, M.J.V.; de Carvalho dos Anjos, V. Long-Term Ripening Evaluation of Ewes’ Cheeses by Fourier-Transformed Infrared Spectroscopy under Real Industrial Conditions. J. Spectrosc. 2018, 1381864. [Google Scholar] [CrossRef]
- Wang, X.; Esquerre, C.; Downey, G.; Henihan, L.; O’Callaghan, D.; O’Donnell, C. Feasibility of Discriminating Dried Dairy Ingredients and Preheat Treatments Using Mid-Infrared and Raman Spectroscopy. Food Anal. Methods 2017, 11, 1380–1389. [Google Scholar] [CrossRef]
- El Darra, N.; Rajha, H.N.; Saleh, F.; Al-Oweini, R.; Maroun, R.G.; Louka, N. Food Fraud Detection in Commercial Pomegranate Molasses Syrups by UV–VIS Spectroscopy, ATR-FTIR Spectroscopy and HPLC Methods. Food Control 2017, 78, 132–137. [Google Scholar] [CrossRef]
- Oroian, M.; Ursachi, F.; Dranca, F. Ultrasound-Assisted Extraction of Polyphenols from Crude Pollen. Antioxidants 2020, 9, 322. [Google Scholar] [CrossRef]
- Pini, R.; Furlanetto, G.; Castellano, L.; Saliu, F.; Rizzi, A.; Tramelli, A. Effects of Stepped-combustion on Fresh Pollen Grains: Morphoscopic, Thermogravimetric, and Chemical Proxies for the Interpretation of Archeological Charred Assemblages. Rev. Palaeobot. Palynol. 2018, 259, 142–158. [Google Scholar] [CrossRef]
- Kasprzyk, I.; Depciuch, J.; Grabek-Lejko, D.; Parlinska-Wojtan, M. FTIR-ATR Spectroscopy of Pollen and Honey as a Tool for Unifloral Honey Authentication. The Case Study of Rape honey. Food Control 2018, 84, 33–40. [Google Scholar] [CrossRef]
- Castiglioni, S.; Astolfi, P.; Conti, C.; Monaci, E.; Stefano, M.; Carloni, P. Morphological, Physicochemical and FTIR Spectroscopic Properties of Bee Pollen Loads from Different Botanical Origin. Molecules 2019, 24, 3974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elbakush, A.E.; Güven, D. Evaluation of Ethanol Tolerance in Relation to Intracellular Storage Compounds of Saccharomyces cerevisiae Using FT-IR Spectroscopy. Process Biochem. 2021, 101, 266–273. [Google Scholar] [CrossRef]
- Feng, G.D.; Zhang, F.; Cheng, L.H.; Xu, X.H.; Zhang, L.; Chen, H.L. Evaluation of FT-IR and Nile Red Methods for Microalgal Lipid Characterization and Biomass Composition Determination. Bioresour. Technol. 2013, 18, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Klaassen, M.; De Vries, E.G.; Masen, M.A. Interpersonal Differences in the Friction Response of Skin Relate to FTIR Measures for Skin Lipids and Hydration. Colloids. Surf. B Biointerfaces 2020, 189, 110883. [Google Scholar] [CrossRef] [PubMed]
- Grace, C.E.E.; Laksmi, P.K.; Meenakshi, S.; Vaidyanathan, S.; Srisudha, S.; Mary, M.B. Biomolecular Transitions and Lipid Accumulation in Green Microalgae Monitored by FTIR and Raman Analysis. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2020, 224, 117382. [Google Scholar] [CrossRef] [PubMed]
- Mujumdar, A.S.; Law, C.L.; Woo, M.W. Freeze Drying: Effects on Sensory and Nutritional Properties. In Encyclopedia of Food and Health; Caballero, B., Finglas, P.M., Toldrá, F., Eds.; Academic Press: Cambridge, MA, USA, 2016; pp. 99–103. ISBN 9780123849533. [Google Scholar]
- Waghmare, R.B.; Perumal, A.B.; Moses, J.A.; Anandharamakrishnan, C. Recent developments in freeze drying of foods. In Innovative Food Processing Technologies; Elsevier: Amsterdam, The Netherlands, 2021; pp. 82–99. [Google Scholar]
- Franceschi, C.R.B.; Smidt, E.C.; Vieira, L.N.; Ribas, L.L.F. Storage and in vitro Germination of Orchids (Orchidaceae) Seeds from Atlantic Forest–Brazil. An. Acad. Bras. Ciênc. 2019, 91, e20180439. [Google Scholar] [CrossRef] [PubMed]
- Koene, F.M.; Amano, É.; Ribas, L.L.F. Asymbiotic Seed Germination and in vitro Seedling Development of Aciantheraprolifera (Orchidaceae). S. Afr. J. Bot. 2019, 121, 83–91. [Google Scholar] [CrossRef]
- Athapattu, M.; Saveh, A.H.; Kazemi, S.M.; Wang, B.; Chizari, M. Measurement of the Femoral Head Diameter at Hemiarthroplasty of the Hip. Proc. Technol. 2014, 17, 217–222. [Google Scholar] [CrossRef] [Green Version]
- Ertas, N.; Aslan, M. Antioxidant and Physicochemical Properties of Cookies Containing Raw and Roasted Hemp Flour. Acta Sci. Pol. Technol. Aliment. 2020, 19, 177–184. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Legume Type | Germination Period, Days | Protein (%) | Fat (%) | Ash (%) | Moisture (%) | Carbohydrates (%) | pH | Acidity (°) |
|---|---|---|---|---|---|---|---|---|
| Bean | 0 | 22.6 ± 0.28 cC | 1.6 ± 0.07 aD | 3.6 ± 0.07 aB | 10.9 ± 0.07 aA | 61.3 ± 0.14 aB | 6.55 ± 0.007 aB | 7.15 ± 0.007 cB |
| 2 | 24.1 ± 0.14 bO | 1.2 ± 0.14 aO | 3.4 ± 0.07 abM | 10.5 ± 0.14 abL | 60.6 ± 0.14 bL | 6.50 ± 0.021 aL | 8.12 ± 0.021 bP | |
| 4 | 26.0 ± 0.28 aY | 1.4 ± 0.07 aY | 3.0 ± 0.14 bV | 10.1 ± 0.14 bUV | 59.7 ± 0.14 cV | 6.41 ± 0.021 bV | 15.07 ± 0.056 aY | |
| Lentil | 0 | 28.6 ± 0.28 bB | 1.2 ± 0.07 aD | 2.6 ± 0.07 bD | 7.6 ± 0.07 aC | 60.0 ± 0.07 aC | 6.63 ± 0.014 aA | 5.16 ± 0.035 cD |
| 2 | 30.0 ± 0.14 aN | 1.1 ± 0.00 aP | 2.8 ± 0.07 bN | 8.9 ± 0.14 aN | 58.9 ± 0.07 bM | 6.51 ± 0.021 bL | 10.66 ± 0.021 bM | |
| 4 | 29.5 ± 0.14 abX | 1.0 ± 0.07 aY | 3.1 ± 0.14 aV | 8.8 ± 0.28 aX | 59.0 ± 0.14 bX | 6.48 ± 0.007 bU | 16.28 ± 0.021 aX | |
| Soybean | 0 | 40.3 ± 0.07 aA | 16.6 ± 0.14 bA | 4.5 ± 0.07 aA | 9.8 ± 0.28 bC | 28.8 ± 0.14 aE | 6.62 ± 0.028 aA | 5.68 ± 0. 007 cC |
| 2 | 40.3 ± 0.07 aL | 17.6 ± 0.07 aL | 4.7 ± 0.07 aL | 10.4 ± 0.07 aL | 27.0 ± 0.07 bO | 6.50 ± 0.007 bL | 9.83 ± 0.042 bO | |
| 4 | 40.2 ± 0.07 aU | 17.9 ± 0.07 aU | 5.1 ± 0.28 aU | 10.5 ± 0.14 aU | 26.3 ± 0.14 cZ | 6.32 ± 0.021 cX | 16.29 ± 0.028 aX | |
| Chickpea | 0 | 19.4 ± 0.07 bD | 5.9 ± 0.14 aC | 3.1 ± 0.07 bC | 10.3 ± 0.28 aB | 61.3 ± 0.14 aA | 6.42 ± 0.007 aC | 4.2 ± 0.141 cE |
| 2 | 20.7 ± 0.28 aP | 5.4 ± 0.00 bN | 3.5 ± 0.07 aM | 9.7 ± 0.07 abM | 60.7 ± 0.07 bL | 6.22 ± 0.028 bM | 10.24 ± 0.021 bN | |
| 4 | 21.1 ± 0.00 aZ | 5.2 ± 0.07 bX | 3.6 ± 0.07 aV | 9.4 ± 0.07 bVX | 60.7 ± 0.07 bU | 6.11 ± 0.014 cY | 20.45 ± 0.629 aV | |
| Lupine | 0 | 39.9 ± 0.14 aA | 9.3 ± 0.21 aB | 3.3 ± 0.07 aB | 7.7 ± 0.14 bD | 39.8 ± 0.14 bD | 5.62 ± 0.007 aD | 14.53 ± 0.014 cA |
| 2 | 39.3 ± 0.07 bM | 7.5 ± 0.00 bM | 3.3 ± 0.07 aM | 10.1 ± 0.07 aL | 39.8 ± 0.07 bN | 5.55 ± 0.007 bN | 20.87 ± 0.035 bL | |
| 4 | 39.4 ± 0.07 bV | 6.9 ± 0.07 cV | 3.4 ± 0.07 aV | 10.3 ± 0.14 aU | 40.0 ± 0.07 aY | 5.53 ± 0.007 bZ | 31.9 ± 0. 007 aU |
| Legume Type | Germination Period, (Days) | Na (mg 100 g−1) | Mg (mg 100 g−1) | Fe (mg 100 g−1) | Zn (mg 100 g−1) |
|---|---|---|---|---|---|
| Bean | 0 | 38.15 ± 2.61 bB | 141.65 ± 0.17 cB | 7.57 ± 0.75 aA | 3.22 ± 0.20 aA |
| 2 | 62.62 ± 2.18 aL | 148.50 ± 0.11 bM | 7.83 ± 0.75 aL | 3.23 ± 0.20 aL | |
| 4 | 64.42 ± 0.71 aV | 152.15 ± 0.01 aX | 7.89 ± 0.80 aU | 3.25 ± 0.20 aU | |
| Lentil | 0 | 26.08 ± 0.61 cD | 122.35 ± 0.06 cD | 2.62 ± 0.18 aC | 3.07 ± 0.20 aA |
| 2 | 48.89 ± 0.94 bM | 126.82 ± 0.09 bO | 2.66 ± 0.18 aN | 3.10 ± 0.20 aL | |
| 4 | 53.52 ± 1.57 aX | 146.63 ± 0.12 aY | 2.75 ± 0.20 aX | 3.12 ± 0.22 aU | |
| Soybean | 0 | 47.50 ± 1.06 cA | 90.30 ± 0.10 cE | 5.31 ± 0.14 aB | 1.91 ± 0.15 aB |
| 2 | 60.38 ± 1.59 bL | 92.41 ± 0.27 bP | 5.32 ± 0.20 aM | 1.93 ± 0.10 aM | |
| 4 | 70.19 ± 2.03 aUV | 106.88 ± 0.08 aZ | 5.38 ± 0.21 aV | 1.94 ± 0.15 aV | |
| Chickpea | 0 | 40.11 ± 1.03 cB | 168.31 ± 0.05 cA | 5.27 ± 0.30 aB | 3.25 ± 0.20 aA |
| 2 | 58.66 ± 1.31 bL | 172.56 ± 0.16 bL | 5.29 ± 0.29 aM | 3.26 ± 0.18 aL | |
| 4 | 74.81 ± 3.50 aU | 174.06 ± 0.03 aU | 5.36 ± 0.30 aV | 3.28 ± 0.20 aU | |
| Lupine | 0 | 31.94 ± 0.66 cC | 128.98 ± 0.02 cC | 4.58 ± 0.26 aB | 3.22 ± 0.20 aA |
| 2 | 45.20 ± 1.25 bM | 137.75 ± 0.11 bN | 4.61 ± 0.20 aM | 3.23 ± 0.20 aL | |
| 4 | 69.43 ± 2.08 aUV | 166.63 ± 0.12 aV | 4.73 ± 0.35 aV | 3.25 ± 0.20 aU |
| Element | Wavelength (nm) | Slit Width (nm) | Fuel Gas Flow Rate (L/min) | Support Gas Flow Rate (L/min) | Flame Type | Pre-Spray Time (s) | Integration Time (s) | Response Time (s) |
|---|---|---|---|---|---|---|---|---|
| Na | 589.0 | 0.2 | 1.8 | 15.0 | Air-C2H2 | 10 | 5 | 1 |
| Mg | 285.2 | 0.7 | 1.8 | 15.0 | Air-C2H2 | 10 | 5 | 1 |
| Fe | 248.3 | 0.2 | 2.2 | 15.0 | Air-C2H2 | 10 | 5 | 1 |
| Zn | 213.9 | 0.7 | 2.0 | 15.0 | Air-C2H2 | 10 | 5 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Atudorei, D.; Stroe, S.-G.; Codină, G.G. Impact of Germination on the Microstructural and Physicochemical Properties of Different Legume Types. Plants 2021, 10, 592. https://doi.org/10.3390/plants10030592
Atudorei D, Stroe S-G, Codină GG. Impact of Germination on the Microstructural and Physicochemical Properties of Different Legume Types. Plants. 2021; 10(3):592. https://doi.org/10.3390/plants10030592
Chicago/Turabian StyleAtudorei, Denisa, Silviu-Gabriel Stroe, and Georgiana Gabriela Codină. 2021. "Impact of Germination on the Microstructural and Physicochemical Properties of Different Legume Types" Plants 10, no. 3: 592. https://doi.org/10.3390/plants10030592