
Evaluation of African Maize Cultivars for Resistance to Fall Armyworm Spodoptera frugiperda (J. E. Smith) (Lepidoptera: Noctuidae) Larvae

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Results
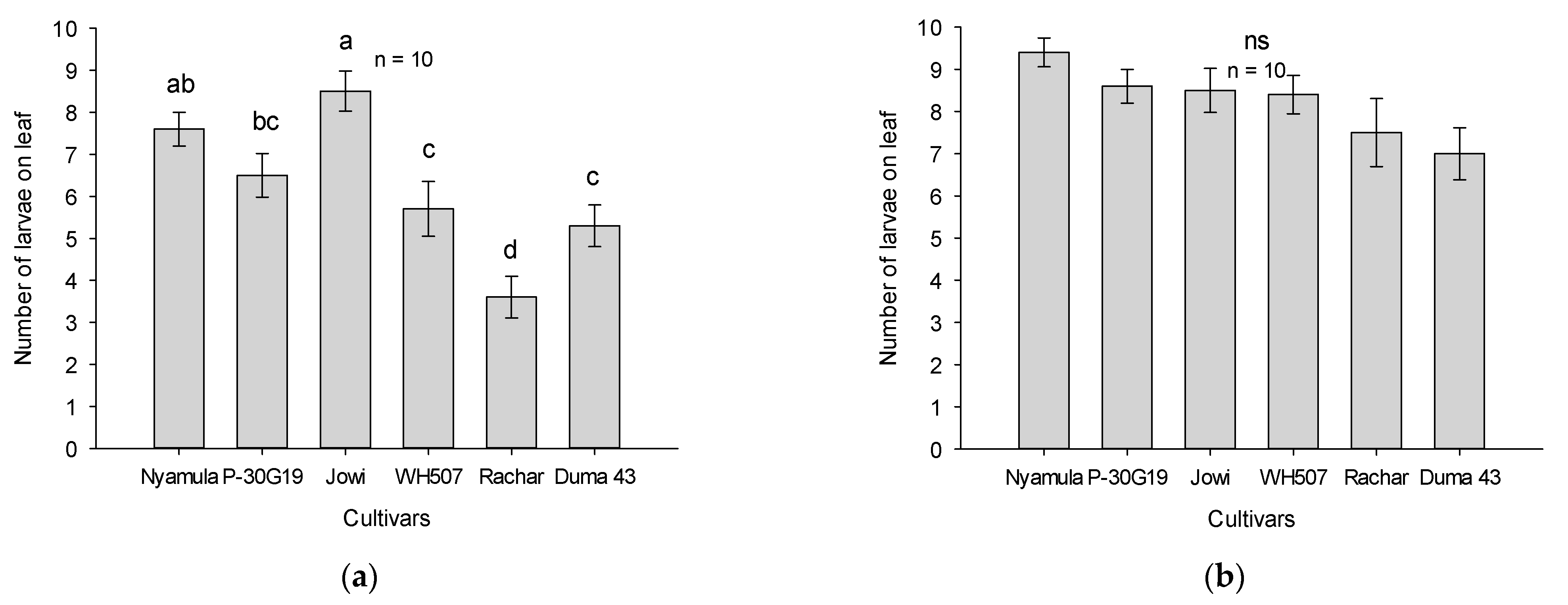
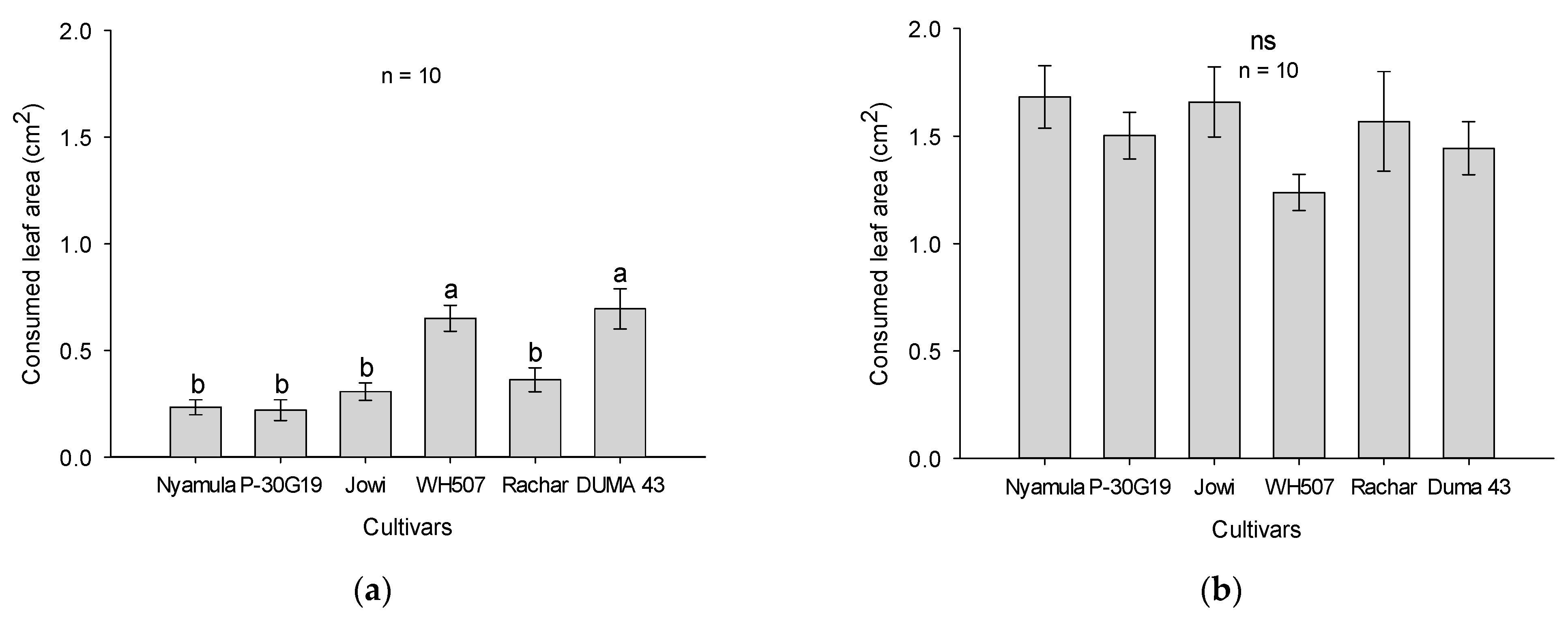
2.1. Arrestment/Dispersal and Feeding Experiments in No-Choice Tests
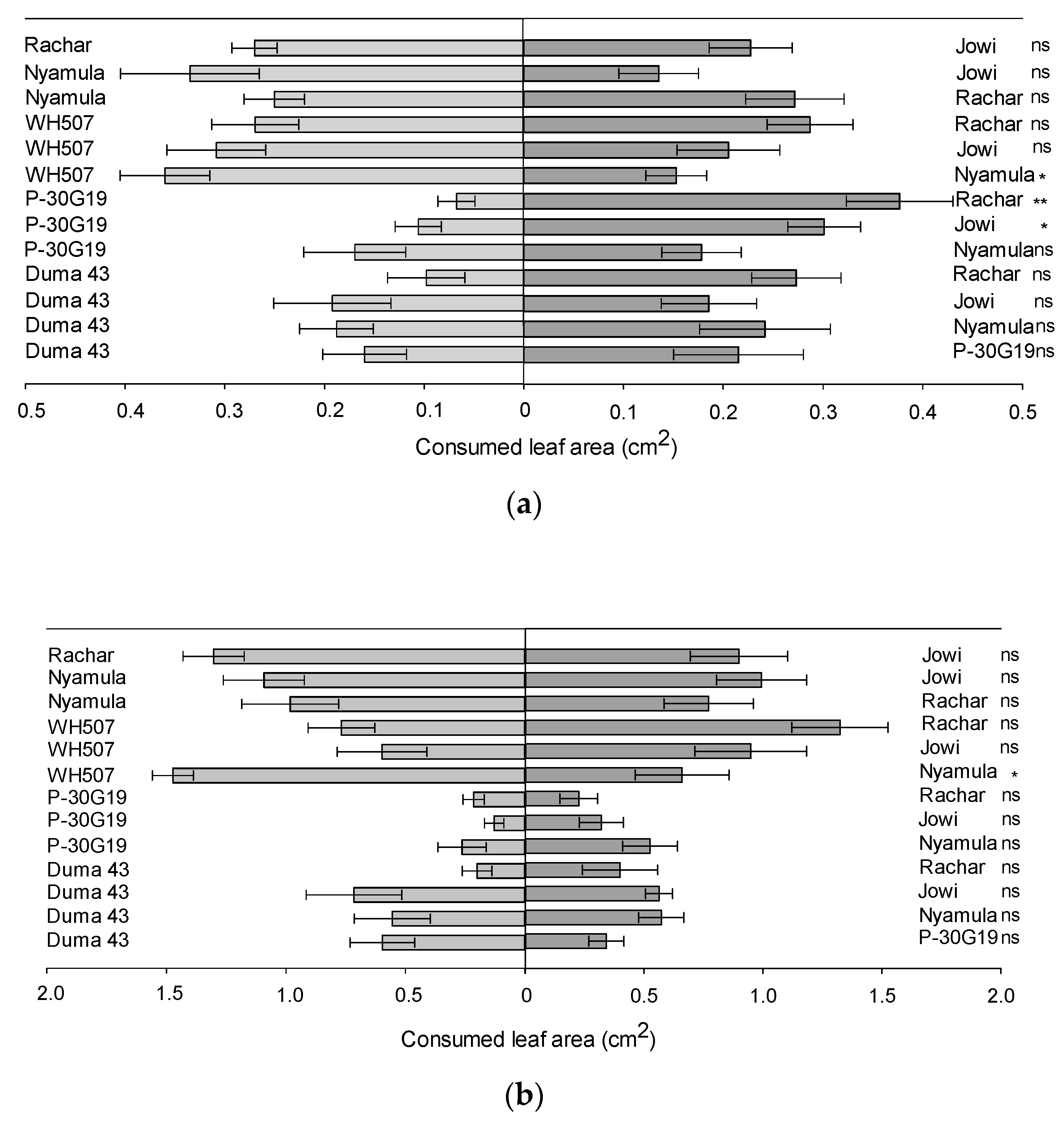
2.2. Orientation/Settling and Feeding in Two-Choice Experiments
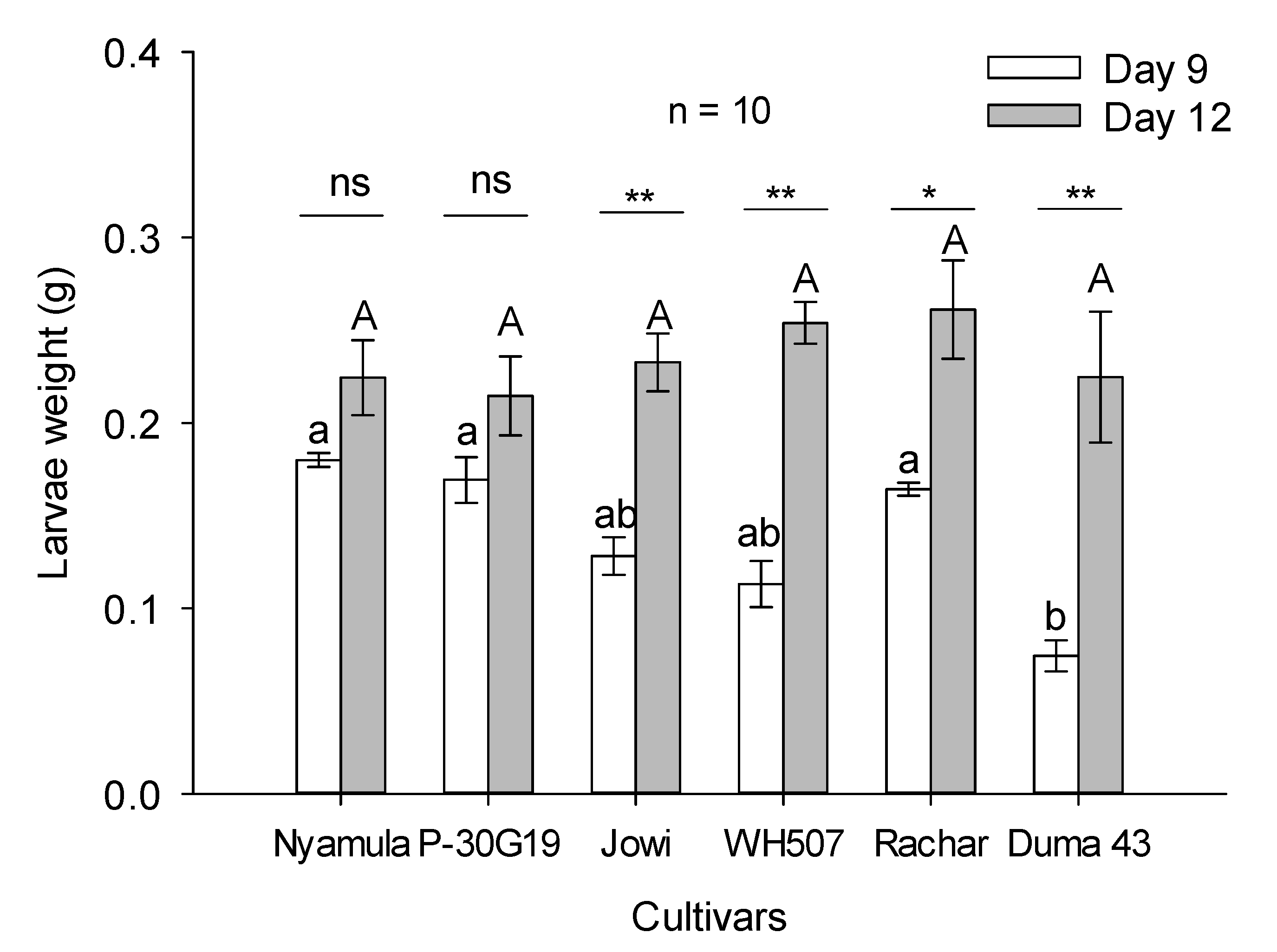
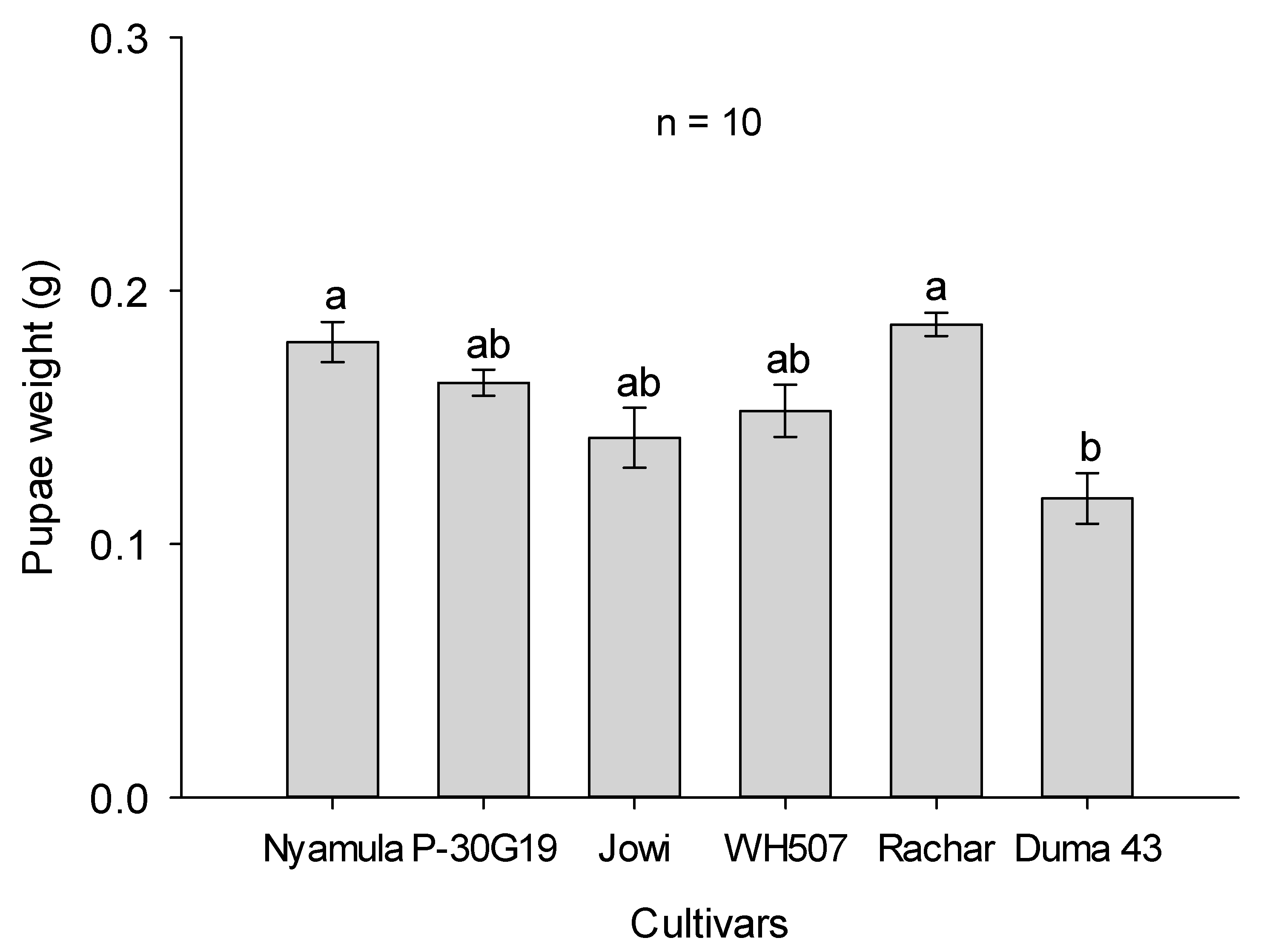
2.3. Larval–Pupal Development Experiment
2.4. Food Assimilation
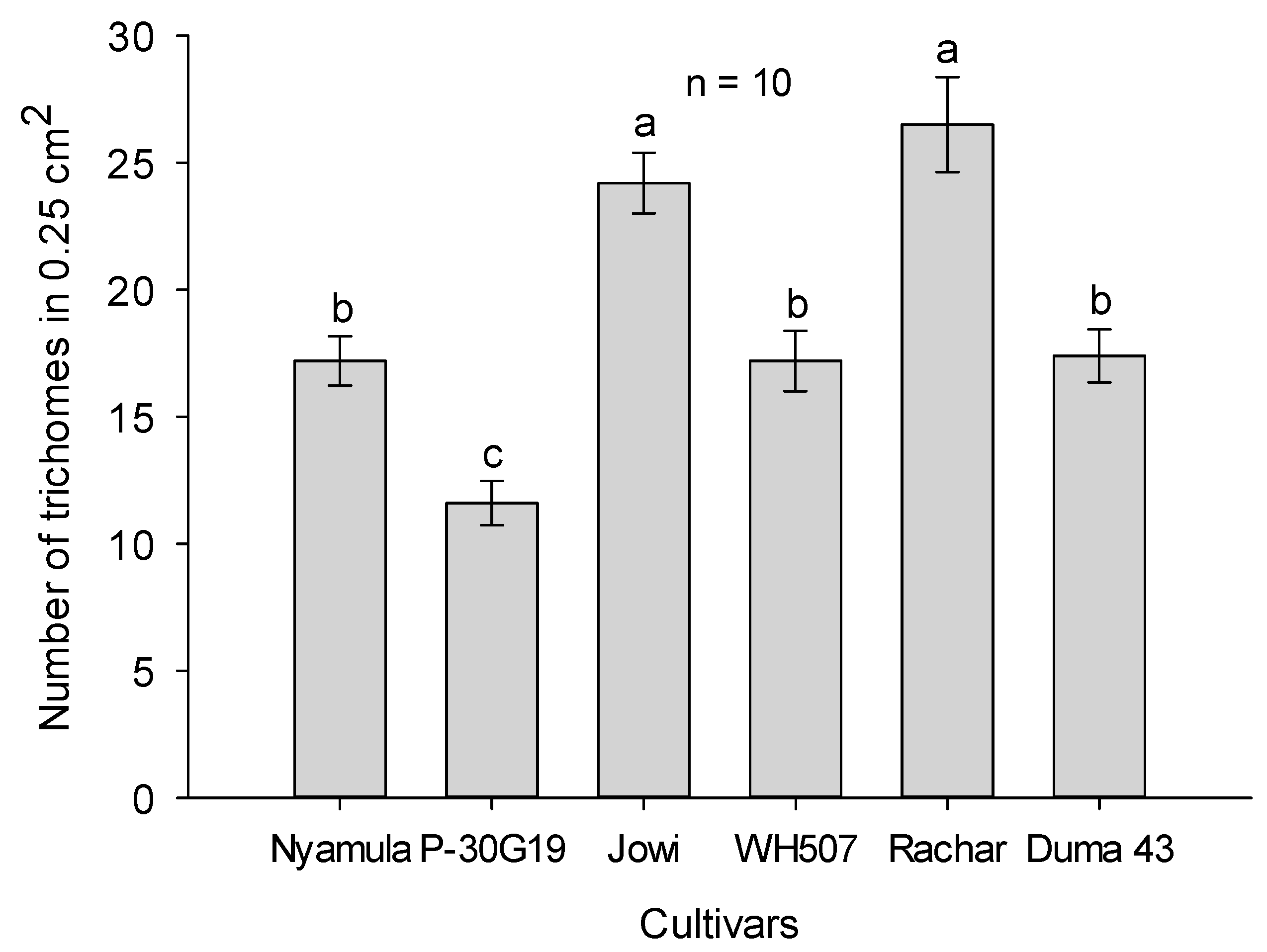
2.5. Leaf Trichome Density
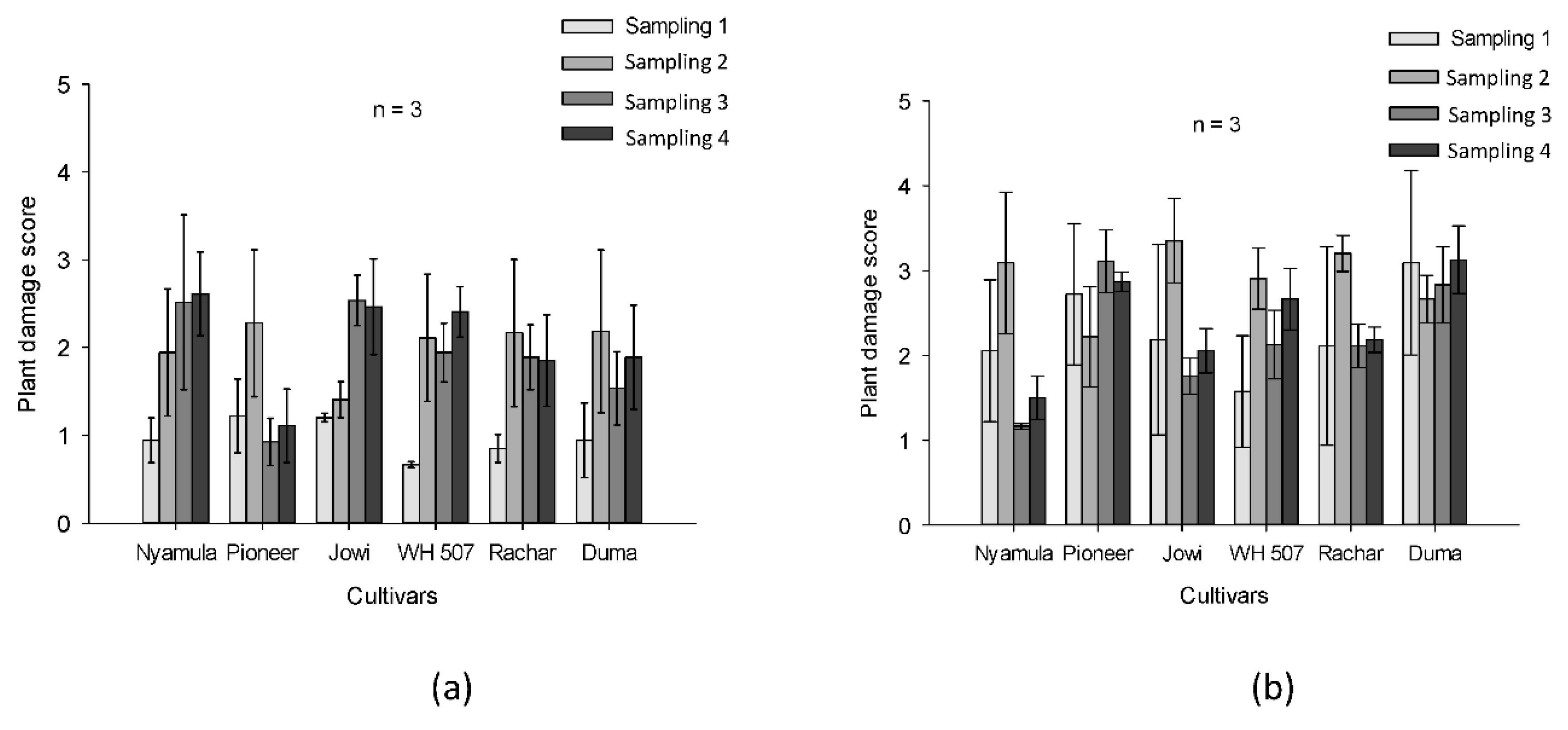
2.6. Field Experiment
3. Discussion
4. Materials and Methods
4.1. Plants
4.2. Insects
4.3. Arrestment-Dispersal and Feeding Experiment
4.4. Orientation-Settling and Feeding Experiment
4.5. Larval-Pupal Development
4.6. Food Assimilation
4.7. Leaf Trichome Density
4.8. Field Experiment
4.9. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Goergen, G.; Kumar, P.L.; Sankung, S.B.; Togola, A.; Tamò, M. First report of outbreaks of the fall armyworm Spodoptera frugiperda (J E Smith) (Lepidoptera, Noctuidae), a new alien invasive pest in West and Central Africa. PLoS ONE 2016, 11, e0165632. [Google Scholar] [CrossRef] [Green Version]
- Day, R.; Abrahams, P.; Bateman, M.; Beale, T.; Clottey, V.; Cock, M.; Colmenarez, Y.; Corniani, N.; Early, R.; Godwin, J.; et al. Fall armyworm: Impacts and implications for Africa. Outlooks Pest Manag. 2017, 28, 196–201. [Google Scholar] [CrossRef] [Green Version]
- Sharanabassapa; Kalleshwaraswamy, C.M.; Asokan, R.; Swamy, H.M.M.; Maruthi, M.S.; Pavithra, H.B.; Hedge, K.; Navi, S.; Prabhu, S.T.; Goergen, G.E. First report of the Fall armyworm, Spodoptera frugiperda (J E Smith) (Lepidoptera: Noctuidae), an alien invasive pest on maize in India. Pest Manag. Hortic. Syst. 2018, 24, 8. [Google Scholar]
- Sun, X.X.; Hu, C.X.; Jia, H.R.; Wu, Q.L.; Shen, X.J.; Jiang, Y.Y.; Wu, K.M. Case study on the first immigration of fall armyworm Spodoptera frugiperda invading into China. J. Integr. Agric. 2019, 3119, 10. [Google Scholar] [CrossRef]
- Chhetri, L.B.; Acharya, B. Fall armyworm (Spodoptera frugiperda): A threat to food security for south Asian country: Control and management options: A review. Farm. Manag. 2019, 4, 38–44. [Google Scholar] [CrossRef]
- Li, Y.; Wang, Z.; Romeis, J. Managing the Invasive Fall Armyworm through Biotech Crops: A Chinese Perspective. Trends Biotechnol. 2021, 39, 105–107. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Wang, Y.P.; Wu, M.F.; Gao, B.Y.; Liu, J.; Lee, G.S.; Otuka, A.; Hu, G. High risk of the fall armyworm invading Japan and the Korean Peninsula via overseas migration. J. Appl. Entomol. 2019, 143, 911–920. [Google Scholar] [CrossRef]
- Food and Agriculture Organization. Fall Armyworm. 2020. Available online: http://www.fao.org/fall-armyworm/monitoring-tools/faw-map/en/ (accessed on 14 August 2020).
- Baudron, F.; Zaman-Allah, M.A.; Chaipa, I.; Chari, N.; Chinwada, P. Understanding the factors influencing fall armyworm (Spodoptera frugiperda J. E. Smith) damage in African smallholder maize fields and quantifying its impact on yield. A case study in Eastern Zimbabwe. Crop Prot. 2019, 120, 141–150. [Google Scholar] [CrossRef]
- Kumela, T.; Simiyu, J.; Sisay, B.; Likhayo, P.; Mendesil, E.; Gohole, L.; Tefera, T. Farmers’ knowledge, perceptions, and management practices of the new invasive pest, fall armyworm (Spodoptera frugiperda) in Ethiopia and Kenya. Int. J. Pest Manag. 2019, 65, 1–9. [Google Scholar] [CrossRef]
- Mihm, J.A. Insect Resistant Maize-Recent Advances and Utilization; Mihm, J.A., Ed.; CIMMYT: El Batan, Mexico, 1997; p. 304. [Google Scholar]
- Mihm, J.A.; Smith, M.E.; Deutsch, J.A. Development of Open-Pollinated Varieties, Non-Conventional Hybrids and Inbred Lines of Tropical Maize with Resistance to Fall Armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae), at CIMMYT. Fla. Entomol. 1988, 71, 262–268. [Google Scholar] [CrossRef]
- Wiseman, B.R.; Davis, F.M. Plant Resistance to Insects Attacking Corn and Grain Sorghum. Fla. Entomol. 1979, 62, 123–130. Available online: http://www.jstor.org/stable/3495461 (accessed on 12 December 2020). [CrossRef]
- Host Plant Resistance to Fall Armyworm: Status and Prospects. Prasanna, B P. WTO Meeting. 19 March 2019. Available online: https://www.wto.org/english/tratop_e/sps_e/faw_2_c_prasanna_thematic_session_faw_19march2019.pdf (accessed on 15 April 2019).
- Odendo, M.; de Groote, H.; Odongo, O.M. Assesment of farmers’ preferences and constraints to maize production in the moist mid-altitute zone of Western Kenya. Afr. Crop Sci. Conf. Proc. 2001, 5, 769–775. [Google Scholar]
- Midega, C.A.O.; Pickett, J.A.; Hooper, A.; Pittchar, J.O.; Khan, Z.R. Maize landaraces are less affected by Striga hermonthica relative to hybrids in Western Kenya. Weed Technol. 2016, 30, 21–28. [Google Scholar] [CrossRef] [Green Version]
- Tamiru, A.; Bruce, T.J.A.; Midega, C.A.O.; Woodcock, C.M.; Birkett, M.A.; Pickett, J.A.; Khan, Z.R. Oviposition Induced Volatile Emissions from African Smallholder Farmers’ Maize Varieties. J. Chem. Ecol. 2012, 38, 231–234. [Google Scholar] [CrossRef]
- Stout, M.J. Reevaluating the conceptual framework for applied research on host-plant resistance. Insect Sci. 2013, 20, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Gatehouse, J.A. Plant resistance towards insect herbivores: A dynamic interaction. New Phytol. 2002, 156, 145–169. [Google Scholar] [CrossRef] [Green Version]
- Withers, T.M. Examining the hierarchy threshold model in a no-choice feeding assay. Entomol. Exp. Appl. 1999, 91, 89–95. [Google Scholar] [CrossRef]
- Knolhoff, L.M.; Heckel, D.G. Behavioral assays for studies of host plant choice and adaptation in herbivorous insects. Annu. Rev. Entomol. 2014, 59, 263–278. [Google Scholar] [CrossRef] [PubMed]
- Schoonhoven, L.M.; van Loon, J.J.A.; Dicke, M. Insect-Plant Biology, 2nd ed.; Oxford University Press: New York, NY, USA, 2007; p. 421. [Google Scholar]
- Yang, G.; Wiseman, B.R.; Isenhour, D.J.; Espelie, K.E. Chemical and ultrastructural analysis of corn cuticular lipids and their effect on feeding by fall armyworm larvae. J. Chem. Ecol. 1993, 19, 2055–2074. [Google Scholar] [CrossRef]
- Yang, G.; Espelie, K.E.; Wiseman, B.R.; Isenhour, D.J. Effect of Corn Foliar Cuticular Lipids on the Movement of Fall Armyworm (Lepidoptera: Noctuidae ) Neonate Larvae. Fla. Entomol. 1993, 76, 302–316. [Google Scholar] [CrossRef]
- Andama, J.B.; Mujiono, K.; Hojo, Y.; Shinya, T.; Galis, I. Nonglandular silicified trichomes are essential for rice defense against chewing herbivores. Plant Cell Environ. 2020, 43, 2019–2032. [Google Scholar] [CrossRef] [PubMed]
- de Fouchier, A.; Sun, X.; Caballero-Vidal, G.; Travaillard, S.; Jacquin-Joly, E.; Montagné, N. Behavioral effect of plant volatiles binding to Spodoptera littoralis larval odorant receptors. Front. Behav. Neurosci. 2018, 12, 264. [Google Scholar] [CrossRef]
- Hedin, P.A.; Davis, F.M.; Williams, W.P.; Hicks, R.P.; Fisher, T.H. Hemicellulose is an important leaf-feeding resistant factor in corn to the fall armyworm. J. Chem. Ecol. 1996, 22, 1655–1668. [Google Scholar] [CrossRef] [PubMed]
- Bergvinson, D.J.; Arnason, J.T.; Hamilton, R.I.; Mihm, J.A.; Jewell, D.C. Determining leaf toughness and its role in maize resistance to the European corn borer (Lepidoptera: Pyralidae). J. Econ. Entomol. 1994, 87, 1743–1748. [Google Scholar] [CrossRef]
- Chen, Y.; Ni, X.; Buntin, G.D. Physiological, nutritional, and biochemical bases of corn resistance to foliage-feeding fall armyworm. J. Chem. Ecol. 2009, 35, 297–306. [Google Scholar] [CrossRef]
- Wiseman, B.R.; Davis, F.M.; Williams, W.P.; Widstrom, N.W. Resistance of a maize population, FAWCC(C5), to fall armyworm larvae (Lepidoptera: Noctuidae). Fla. Entomol. 1996, 79, 329–336. [Google Scholar] [CrossRef]
- De La Rosa-Cancino, W.; Rojas, J.C.; Cruz-Lopez, L.; Castillo, A.; Malo, E.A. Attraction, feeding preference, and performance of Spodoptera frugiperda larvae (Lepidoptera: Noctuidae) reared on two varieties of maize. Environ. Entomol. 2016, 45, 384–389. [Google Scholar] [CrossRef]
- Wiseman, B.R.; Widstrom, N.W. Mechanisms of Resistance in ‘Zapalote Chico’ Corn Silks to Fall Armyworm (Lepidoptera: Noctuidae) Larvae. J. Econ. Entomol. 1986, 79, 1390–1393. [Google Scholar] [CrossRef]
- Sen-Seong, N.G.; Davis, F.M.; Williams, W.P. Survival, Growth, and Reproduction of the Fall Armyworm (Lepidoptera: Noctuidae) as Affected by Resistant Corn Genotypes1. J. Econ. Entomol. 1985, 78, 967–971. [Google Scholar]
- Chuang, W.P.; Ray, S.; Acevedo, F.E.; Peiffer, M.; Felton, G.W.; Luthe, D.S. Herbivore cues from the fall armyworm (Spodoptera frugiperda) larvae trigger direct defenses in maize. Mol. Plant Microbe Interact. 2014, 27, 461–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chuang, W.P.; Herde, M.; Ray, S.; Castano-Duque, L.; Howe, G.A.; Luthe, D.S. Caterpillar attack triggers accumulation of the toxic maize protein RIP2. New Phytol. 2014, 201, 928–939. [Google Scholar] [CrossRef] [PubMed]
- Hariprasad, K.V.; van Emden, H.F. Mechanisms of partial plant resistance to diamondback moth (Plutella xylostella) in brassicas. Int. J. Pest Manag. 2010, 56, 15–22. [Google Scholar] [CrossRef]
- Rojas, J.C.; Kolomiets, M.V.; Bernal, J.S. Nonsensical choices? Fall armyworm moths choose seemingly best or worst hosts for their larvae, but neonate larvae make their own choices. PLoS ONE 2018, 13, e0197628. [Google Scholar] [CrossRef]
- Mihm, J.A. Efficient Mass-Rearing and Infestation Techniques to Screen for Host Plant Resistance to Fall Armyworm, Spodoptera Frugiperda; Centro Internacional de Mejoramiento de Maiz y Trigo-CIMMYT, Ed.; CIMMYT: El Batan, Mexico, 1983. [Google Scholar]
- Rosenthal, J.P.; Welter, S.C. Tolerance to herbivory by a stemboring caterpillar in architecturally distinct maizes and wild relatives. Oecologia 1995, 102, 146–155. [Google Scholar] [CrossRef]
- Ni, X.; Xu, W.; Blanco, M.H.; Williams, W.P. Evaluation of fall armyworm resistance in maize germplasm lines using visual leaf injury rating and predator survey. Insect. Sci. 2014, 21, 541–555. [Google Scholar] [CrossRef]
- Sueldo, M.R.; Bruzzone, O.A.; Virla, E.G. Characterization of the earwig, Doru lineare, as a predator of larvae of the fall armyworm, Spodoptera frugiperda: A functional response study. J. Insect. Sci. 2010, 10, 1–10. [Google Scholar] [CrossRef] [Green Version]
- van Huis, A. Integrated Pest Management in the Small Farmer’s Maize Crop in Nicaragua; H. Veenman & Zonen, B.V.: Wageningen, The Netherlands, 1981; p. 221. [Google Scholar]
- Turlings, T.C.J.; Tumlinson, J.H.; Lewis, W.J. Exploitation of Herbivore-Induced Plant Odors By Host-Seeking Parasitic Wasps. Science 1990, 250, 1251–1253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roque-Romero, L.; Cisneros, J.; Rojas, J.C.; Ortiz-Carreon, F.R.; Malo, E.A. Attraction of Chelonus insularis to host and host habitat volatiles during the search of Spodoptera frugiperda eggs. Biol Control 2020, 140. [Google Scholar] [CrossRef]
- Bernasconi Ockroy, M.L.; Turlings, T.C.J.; Edwards, P.J.; Fritzsche-Hoballah, M.E.; Ambrosetti, L.; Bassetti, P.; Dorn, S. Response of natural populations of predators and parasitoids to artificially induced volatile emissions in maize plants (Zea mays L.). Agric. Entomol. 2001, 3, 201–209. [Google Scholar] [CrossRef]
- Publication 75A, Guide to Weed Control: Field Crops. Ministry of Agriculture, Food and Rural Affairs Ontario. 2020. Available online: http://www.omafra.gov.on.ca/english/crops/pub75/pub75A/pub75A.pdf (accessed on 18 December 2020).
- Köhler, A.; Maag, D.; Veyrat, N.; Glauser, G.; Wolfender, J.L.; Turlings, T.C.J.; Erb, M. Within-plant distribution of 1,4-benzoxazin-3-ones contributes to herbivore niche differentiation in maize. Plant. Cell Environ. 2015, 38, 1081–1093. [Google Scholar] [CrossRef]
- Smith, C.M.; Khan, Z.K.; Pathak, M.D. (Eds.) Techniques for Evaluating Resistance in Crop Plants; Lewis Publishers CRC Press: Boca Raton, FL, USA, 1994; pp. 79, 320. [Google Scholar]
- Cheruiyot, D.; Midega, C.A.O.; van den Berg, J.; Pickett, J.A.; Khan, Z.R. Suitability of brachiaria grass as a trap crop for management of Chilo partellus. Entomol. Exp. Appl. 2017, 166, 139–148. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Maize Cultivar | Percentage of Mortality (%) | Percentage of Pupation (%) | Mean Larval Period (Days) | Growth Index |
|---|---|---|---|---|
| Nyamula | 50 | 50 | 15.0 | 3.33 |
| Pioneer 30G19 | 30 | 70 | 15.0 | 4.67 |
| Jowi | 30 | 70 | 15.9 | 4.41 |
| WH507 | 40 | 60 | 15.0 | 4.00 |
| Rachar | 20 | 80 | 15.0 | 5.33 |
| SC Duma 43 | 50 | 50 | 16.8 | 2.98 |
| Maize Cultivar | Assimilated Food Weight (g) | Consumed Leaf Weight (g) |
|---|---|---|
| Nyamula | 0.042 ± 0.006 | 0.08 ± 0.01 |
| Pioneer 30G19 | 0.050 ± 0.007 | 0.08 ± 0.01 |
| Jowi | 0.037 ± 0.003 | 0.11 ± 0.01 |
| WH507 | 0.057 ± 0.011 | 0.10 ± 0.03 |
| Rachar | 0.056 ± 0.006 | 0.07 ± 0.01 |
| SC Duma 43 | 0.043 ± 0.004 | 0.08 ± 0.02 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chiriboga Morales, X.; Tamiru, A.; Sobhy, I.S.; Bruce, T.J.A.; Midega, C.A.O.; Khan, Z. Evaluation of African Maize Cultivars for Resistance to Fall Armyworm Spodoptera frugiperda (J. E. Smith) (Lepidoptera: Noctuidae) Larvae. Plants 2021, 10, 392. https://doi.org/10.3390/plants10020392
Chiriboga Morales X, Tamiru A, Sobhy IS, Bruce TJA, Midega CAO, Khan Z. Evaluation of African Maize Cultivars for Resistance to Fall Armyworm Spodoptera frugiperda (J. E. Smith) (Lepidoptera: Noctuidae) Larvae. Plants. 2021; 10(2):392. https://doi.org/10.3390/plants10020392
Chicago/Turabian StyleChiriboga Morales, Xavier, Amanuel Tamiru, Islam S. Sobhy, Toby J. A. Bruce, Charles A. O. Midega, and Zeyaur Khan. 2021. "Evaluation of African Maize Cultivars for Resistance to Fall Armyworm Spodoptera frugiperda (J. E. Smith) (Lepidoptera: Noctuidae) Larvae" Plants 10, no. 2: 392. https://doi.org/10.3390/plants10020392