
Genetic and Molecular Characterization of a Self-Compatible Brassica rapa Line Possessing a New Class II S Haplotype

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Results
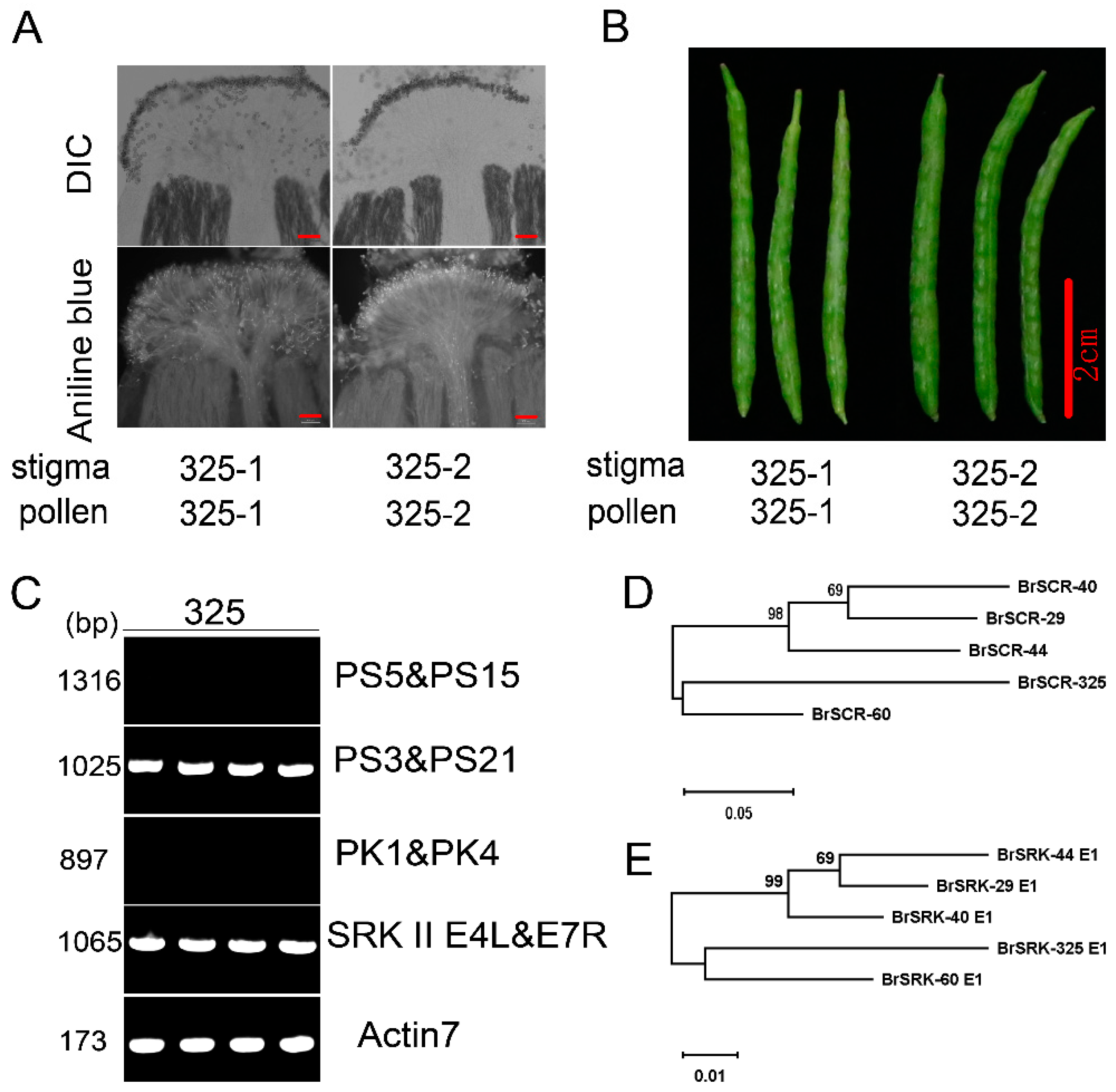
2.1. Determination of a New Class II S Haplotype in an SC B. rapa
2.2. Genetic Analysis of SC Trait in ‘325’
2.3. Nanopore Sequencing and De Novo Assembly the Genome of SC Line ‘325’
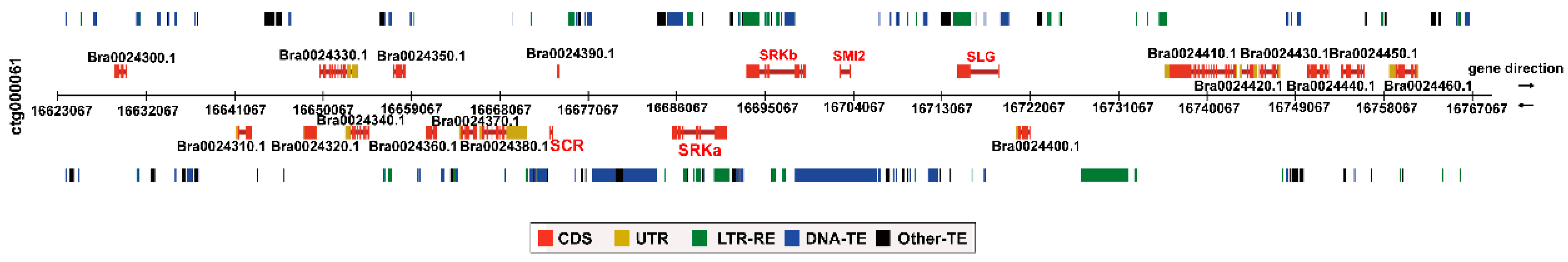
2.4. S Locus Organization of BrS-325
2.5. Comparative Analysis of the S locus of Two Class II S Haplotypes
2.6. Expression Analysis of SRK and SCR
2.7. Amino Acid Sequence Alignment and Variation Analysis of Class II SRK and SCR
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. S Haplotype Determination and Genetic Analysis
4.3. Nanopore Sequencing and Genome Assembly
4.3.1. Genomic DNA Extraction, Nanopore Sequencing, and Genome Assembly
4.3.2. Genomic Annotation
4.4. Aniline Blue Staining Assay
4.5. RNA Extraction and RT-qPCR
4.6. Sequence Alignment and Phylogenetic Analysis
4.7. Synteny Assay of Two Class II S Locus
4.8. Self-Incompatibility Phenotype Assay
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nasrallah, J.B. Self-incompatibility in the Brassicaceae: Regulation and mechanism of self-recognition. Curr. Top. Dev. Biol. 2019, 131, 435–452. [Google Scholar]
- Nasrallah, J.B.; Nasrallah, M.E. Pollen-stigma signaling in the sporophytic self-incompatibility response. Plant Cell. 1993, 5, 1325–1335. [Google Scholar] [CrossRef] [PubMed]
- Casselman, A.L.; Vrebalov, J.; Conner, J.A.; Singhal, A.; Giovannoni, J.; Nasrallah, M.E.; Nasrallah, J.B. Determining the physical limits of the Brassica S locus by recombinational analysis. Plant Cell. 2000, 12, 23–33. [Google Scholar] [CrossRef] [Green Version]
- Takayama, S.; Isogai, A.; Tsukamoto, C.; Ueda, Y.; Hinata, K.; Okazaki, K.; Suzuki, A. Sequences of S-glycoproteins, products of the Brassica campestris self-incompatibility locus. Nature 1987, 326, 102–105. [Google Scholar] [CrossRef]
- Nasrallah, J.B. Cell–cell signaling in the self-incompatibility response. Curr. Opin. Plant Biol. 2000, 3, 368–373. [Google Scholar] [CrossRef]
- Shiba, H.; Kimura, N.; Takayama, S.; Hinata, K.; Suzuki, A.; Isogai, A. Alteration of the self-incompatibility phenotype in Brassica by transformation of the antisense SLG gene. Biosci. Biotech. Bioch. 2000, 64, 1016–1024. [Google Scholar] [CrossRef] [Green Version]
- Takayama, S.; Shiba, H.; Iwano, M.; Asano, K.; Hara, M.; Che, F.S.; Watanabe, M.; Hinata, K.; Isogai, A. Isolation and characterization of pollen coat proteins of Brassica campestris that interact with S locus-related glycoprotein1 involved in pollen-stigma adhesion. Proc. Natl. Acad. Sci. USA 2000, 97, 3765–3770. [Google Scholar] [CrossRef] [PubMed]
- Goring, D.R.; Rothstein, S.J. The S-locus receptor kinase gene in a self-incompatible Brassica napus line encodes a functional serine/threonine kinase. Plant Cell. 1992, 4, 1273–1281. [Google Scholar] [PubMed] [Green Version]
- Silva, N.F.; Stone, S.L.; Christie, L.N.; Sulaman, W.; Nazarian, K.A.; Burnett, L.A.; Arnoldo, M.A.; Rothstein, S.J.; Goring, D.R. Expression of the S receptor kinase in self-compatible Brassica napus cv. Westar leads to the allele-specific rejection of self-incompatible Brassica napus pollen. Mol. Genet. Genom. 2001, 265, 552–559. [Google Scholar] [CrossRef]
- Stein, J.C.; Howlett, B.; Boyes, D.C.; Nasrallah, M.E.; Nasrallah, J.B. Molecular cloning of a putative receptor protein kinase gene encoded at the self-incompatibility locus of Brassica oleracea. Proc. Natl. Acad. Sci. USA 1991, 88, 8816–8820. [Google Scholar] [CrossRef] [Green Version]
- Takasaki, T.; Hatakeyama, K.; Suzuki, G.; Watanabe, M.; Isogai, A.; Hinata, K. The S receptor kinase determines self-incompatibility in Brassica stigma. Nature 2000, 403, 913–916. [Google Scholar] [CrossRef]
- Suzuki, G.; Kai, N.; Hirose, T.; Fukui, K.; Nishio, T.; Takayama, S.; Isogai, A.; Watanabe, M.; Hinata, K. Genomic Organization of the S locus: Identification and characterization of genes in SLG/SRK region of S9 haplotype of Brassica campestris (syn. rapa). Genetics 1999, 151, 391–400. [Google Scholar] [CrossRef]
- Shiba, H.; Takayama, S.; Iwano, M.; Shimosato, H.; Funato, M.; Nakagawa, T.; Che, F.S.; Suzuki, G.; Watanabe, M.; Hinata, K.; et al. A pollen coat protein, SP11/SCR, determines the pollen S-specificity in the self-incompatibility of Brassica species. Plant Physiol. 2001, 125, 2095–2103. [Google Scholar] [CrossRef] [Green Version]
- Takayama, S.; Shiba, H.; Iwano, M.; Shimosato, H.; Che, F.S.; Kai, N.; Watanabe, M.; Suzuki, G.; Hinata, K.; Isogai, A. The pollen determinant of self-incompatibility in Brassica campestris. Proc. Natl. Acad. Sci. USA 2000, 97, 1920–1925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takayama, S.; Shimosato, H.; Shiba, H.; Funato, M.; Che, F.S.; Watanabe, M.; Iwano, M.; Isogai, A. Direct ligand-receptor complex interaction controls Brassica self-incompatibility. Nature 2001, 413, 534–538. [Google Scholar] [CrossRef] [PubMed]
- Kachroo, A.; Schopfer, C.R.; Nasrallah, M.E.; Nasrallah, J.B. Allele-specific receptor-ligand interactions in Brassica self-incompatibility. Science 2001, 293, 1824–1826. [Google Scholar] [CrossRef]
- Nasrallah, J.B.; Nishio, T.; Nasrallah, M.E. The self-incompatibility genes of Brassica: Expression and use in genetic ablation of floral tissues. Annu. Rev. Plant Biol. 1991, 42, 393–422. [Google Scholar] [CrossRef]
- Shiba, H.; Iwano, M.; Entani, T.; Ishimoto, K.; Shimosato, H.; Che, F.S.; Satta, Y.; Ito, A.; Takada, Y.; Watanabe, M.; et al. The dominance of alleles controlling self-incompatibility in Brassica pollen is regulated at the RNA level. Plant Cell. 2002, 14, 491–504. [Google Scholar] [CrossRef] [Green Version]
- Kakizaki, T.; Takada, Y.; Ito, A.; Suzuki, G.; Shiba, H.; Takayama, S.; Isogai, A.; Watanabe, M. Linear dominance relationship among four class-II S haplotypes in pollen is determined by the expression of SP11 in Brassica self-incompatibility. Plant Cell Physiol. 2003, 44, 70–75. [Google Scholar] [CrossRef] [Green Version]
- Tarutani, Y.; Shiba, H.; Iwano, M.; Kakizaki, T.; Suzuki, G.; Watanabe, M.; Isogai, A.; Takayama, S. Trans-acting small RNA determines dominance relationships in Brassica self-incompatibility. Nature 2010, 466, 983–986. [Google Scholar] [CrossRef]
- Yasuda, S.; Wada, Y.; Kakizaki, T.; Tarutani, Y.; Miura-Uno, E.; Murase, K.; Fujii, S.; Hioki, T.; Shimoda, T.; Takada, Y.; et al. A complex dominance hierarchy is controlled by polymorphism of small RNAs and their targets. Nat. Plants 2016, 3, 16206. [Google Scholar] [CrossRef] [PubMed]
- Shiba, H.; Kakizaki, T.; Iwano, M.; Tarutani, Y.; Watanabe, M.; Isogai, A.; Takayama, S. Dominance relationships between self-incompatibility alleles controlled by DNA methylation. Nat. Genet. 2006, 38, 297–299. [Google Scholar] [CrossRef] [PubMed]
- Hatakeyama, K.; Watanabe, M.; Takasaki, T.; Ojima, K.; Hinata, K. Dominance relationships between S-alleles in self-incompatible Brassica campestris L. Heredity 1998, 80, 241–247. [Google Scholar] [CrossRef]
- Hatakeyama, K.; Takasaki, T.; Suzuki, G.; Nishio, T.; Watanabe, M.; Isogai, A.; Hinata, K. The S receptor kinase gene determines dominance relationships in stigma expression of self-incompatibility in Brassica. Plant J. 2001, 26, 69–76. [Google Scholar] [CrossRef] [Green Version]
- Nou, S.; Watanabe, M.; Isogai, A.; Hinata, K. Comparison of S-alleles and S-glycoproteins between two wild populations of Brassica campestris in Turkey and Japan. Sex. Plant Reprod. 1993, 6, 79–86. [Google Scholar] [CrossRef]
- Ockendon, D. The S-allele collection of Brassica oleracea. Acta Hortic. 2000, 539, 25–30. [Google Scholar] [CrossRef]
- Okamoto, S.; Odashima, M.; Fujimoto, R.; Sato, Y.; Kitashiba, H.; Nishio, T. Self-compatibility in Brassica napus is caused by independent mutations in S-locus genes. Plant J. 2007, 50, 391–400. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.; Ma, C.; Zhang, X.; Li, F.; Zhang, J.; Zhai, W.; Wang, Y.; Tu, J.; Shen, J.; Fu, T. The genetic characterization of self-incompatibility in a Brassica napus line with promising breeding potential. Mol. Breed. 2013, 31, 485–493. [Google Scholar] [CrossRef]
- Sato, Y.; Sato, K.; Nishio, T. Interspecific pairs of class II S haplotypes having different recognition specificities between Brassica oleracea and Brassica rapa. Plant Cell Physiol. 2006, 47, 340–345. [Google Scholar] [CrossRef] [Green Version]
- Fukai, E.; Fujimoto, R.; Nishio, T. Genomic organization of the S core region and the S flanking regions of a class-II S haplotype in Brassica rapa. Mol. Genet. Genom. 2003, 269, 361–369. [Google Scholar] [CrossRef]
- Kakizaki, T.; Takada, Y.; Fujioka, T.; Suzuki, G.; Satta, Y.; Shiba, H.; Isogai, A.; Takayama, S.; Watanabe, M. Comparative analysis of the S-intergenic region in class-II S haplotypes of self-incompatible Brassica rapa (syn. campestris). Genes Genet. Syst. 2006, 81, 63–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Cai, X.; Wu, J.; Liu, M.; Grob, S.; Cheng, F.; Liang, J.; Cai, C.; Liu, Z.; Liu, B.; et al. Improved Brassica rapa reference genome by single-molecule sequencing and chromosome conformation capture technologies. Hortic. Res. 2018, 5, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belser, C.; Istace, B.; Denis, E.; Dubarry, M.; Baurens, F.-C.; Falentin, C.; Genete, M.; Berrabah, W.; Chèvre, A.-M.; Delourme, R.; et al. Chromosome-scale assemblies of plant genomes using nanopore long reads and optical maps. Nat. Plants 2018, 4, 879–887. [Google Scholar] [CrossRef]
- Xiao, Z.; Han, F.; Hu, Y.; Xue, Y.; Fang, Z.; Yang, L.; Zhang, Y.; Liu, Y.; Li, Z.; Wang, Y. Overcoming cabbage crossing incompatibility by the development and application of self-compatibility-QTL-specific markers and genome-wide background analysis. Front. Plant Sci. 2019, 10, 189. [Google Scholar] [CrossRef] [Green Version]
- Osaka, M.; Nabemoto, M.; Maeda, S.; Sakazono, S.; Masuko-Suzuki, H.; Ito, K.; Takada, Y.; Kobayashi, I.; Lim, Y.P.; Nakazono, M.; et al. Genetic and tissue-specific RNA-sequencing analysis of self-compatible mutant TSC28 in Brassica rapa L. toward identification of a novel self-incompatibility factor. Genes Genet. Syst. 2019, 94, 167–176. [Google Scholar] [CrossRef] [Green Version]
- Murase, K.; Shiba, H.; Iwano, M.; Che, F.S.; Watanabe, M.; Isogai, A.; Takayama, S. A membrane-anchored protein kinase involved in Brassica self-incompatibility signaling. Science 2004, 303, 1516–1519. [Google Scholar] [CrossRef] [PubMed]
- Hatakeyama, K.; Horisaki, A.; Niikura, S.; Narusaka, Y.; Abe, H.; Yoshiaki, H.; Ishida, M.; Fukuoka, H.; Matsumoto, S. Mapping of quantitative trait loci for high level of self-incompatibility in Brassica rapa L. Genome 2010, 53, 257–265. [Google Scholar] [CrossRef]
- Isokawa, S.; Osaka, M.; Shirasawa, A.; Kikuta, R.; Komatsu, S.; Horisaki, A.; Niikura, S.; Takada, Y.; Shiba, H.; Isogai, A. Novel self-compatible lines of Brassica rapa L. isolated from the Japanese bulk-populations. Genes Genet. Syst. 2010, 85, 87–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tochigi, T.; Udagawa, H.; Li, F.; Kitashiba, H.; Nishio, T. The self-compatibility mechanism in Brassica napus L. is applicable to F1 hybrid breeding. Theor. Appl. Genet. 2011, 123, 475–482. [Google Scholar] [CrossRef]
- Gao, C.; Zhou, G.; Ma, C.; Zhai, W.; Zhang, T.; Liu, Z.; Yang, Y.; Wu, M.; Yue, Y.; Duan, Z.; et al. Helitron-like transposons contributed to the mating system transition from out-crossing to self-fertilizing in polyploid Brassica napus L. Sci. Rep. 2016, 6, 33785. [Google Scholar] [CrossRef] [Green Version]
- Goring, D.R.; Glavin, T.L.; Schafer, U.; Rothstein, S.J. An S receptor kinase gene in self-compatible Brassica napus has a 1-bp deletion. Plant Cell. 1993, 5, 531–539. [Google Scholar] [PubMed]
- Hee-Jeong, J.; Nasar Uddin, A.; Jong-In, P.; Senthil Kumar, T.; Hye-Ran, K.; Yong-Gu, C.; Ill-Sup, N. Analysis of S-locus and expression of S-alleles of self-compatible rapid-cycling Brassica oleracea ‘TO1000DH3’. Mol. Biol. Rep. 2014, 41, 6441–6448. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Yang, Y.; Li, B.; Liu, Z.; Khan, F.; Zhang, T.; Zhou, G.; Tu, J.; Shen, J.; Yi, B.; et al. Functional analysis of M-locus protein kinase revealed a novel regulatory mechanism of self-incompatibility in Brassica napus L. Int. J. Mol. Sci. 2019, 20, 3303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kakita, M.; Murase, K.; Iwano, M.; Matsumoto, T.; Watanabe, M.; Shiba, H.; Isogai, A.; Takayama, S. Two distinct forms of M-locus protein kinase localize to the plasma membrane and interact directly with S-locus receptor kinase to transduce self-incompatibility signaling in Brassica rapa. Plant Cell. 2007, 19, 3961–3973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, T.; Mazzurco, M.; Sulaman, W.; Matias, D.D.; Goring, D.R. Binding of an arm repeat protein to the kinase domain of the S-locus receptor kinase. Proc. Natl. Acad. Sci. USA 1998, 95, 382–387. [Google Scholar] [CrossRef] [Green Version]
- Stone, S.L.; Arnoldo, M.; Goring, D.R. A breakdown of Brassica self-incompatibility in ARC1 antisense transgenic plants. Science 1999, 286, 1729–1731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitashiba, H.; Liu, P.; Nishio, T.; Nasrallah, J.B.; Nasrallah, M.E. Functional test of Brassica self-incompatibility modifiers in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2011, 108, 18173–18178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Indriolo, E.; Safavian, D.; Goring, D.R. The ARC1 E3 ligase promotes two different self-pollen avoidance traits in Arabidopsis. Plant Cell. 2014, 26, 1525–1543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, Z.; Dou, S.; Liu, Z.; Li, B.; Yi, B.; Shen, J.; Tu, J.; Fu, T.; Dai, C.; Ma, C. Comparative phosphoproteomic analysis of compatible and incompatible pollination in Brassica napus L. Acta Bioch. Bioph. Sin. 2020, 52, 446–456. [Google Scholar] [CrossRef]
- Xu, Q.; Guo, S.R.; Li, L.; An, Y.H.; Shu, S.; Sun, J. Proteomics analysis of compatibility and incompatibility in grafted cucumber seedlings. Plant Physiol. Biochem. 2016, 105, 21–28. [Google Scholar] [CrossRef]
- Sankaranarayanan, S.; Jamshed, M.; Samuel, M.A. Proteomics approaches advance our understanding of plant self-incompatibility response. J. Proteome Res. 2013, 12, 4717–4726. [Google Scholar] [CrossRef] [PubMed]
- Kenney, P.; Sankaranarayanan, S.; Balogh, M.; Indriolo, E. Expression of Brassica napus GLO1 is sufficient to breakdown artificial self-incompatibility in Arabidopsis thaliana. Plant Reprod. 2020, 33, 159–171. [Google Scholar] [CrossRef]
- Samuel, M.A.; Chong, Y.T.; Haasen, K.E.; Aldea-Brydges, M.G.; Stone, S.L.; Goring, D.R. Cellular pathways regulating responses to compatible and self-incompatible pollen in Brassica and Arabidopsis stigmas intersect at Exo70A1, a putative component of the exocyst complex. Plant Cell. 2009, 21, 2655–2671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sankaranarayanan, S.; Jamshed, M.; Samuel, M.A. Degradation of glyoxalase I in Brassica napus stigma leads to self-incompatibility response. Nat. Plants 2015, 1, 15185. [Google Scholar] [CrossRef]
- Scandola, S.; Samuel, M.A. A flower-specific phospholipase D is a stigmatic compatibility factor targeted by the self-incompatibility response in Brassica napus. Curr. Biol. 2019, 29, 506–512.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, Z.; Zhang, Y.; Tu, J.; Shen, J.; Yi, B.; Fu, T.; Dai, C.; Ma, C. The Brassica napus GATA transcription factor BnA5.ZML1 is a stigma compatibility factor. J. Integr. Plant Biol. 2020, 62, 1112–1131. [Google Scholar] [CrossRef] [PubMed]
- Nou, I.S.; Watanabe, M.; Isogai, A.; Shiozawa, H.; Suzuki, A.; Hinata, K. Variation of S-alleles and S-glycoproteins in a naturalized population of self-incompatible Brassica campestris L. Jpn. J. Genet. 1991, 66, 227–239. [Google Scholar] [CrossRef] [Green Version]
- Peters, J.L.; Cnudde, F.; Gerats, T. Forward genetics and map-based cloning approaches. Trends Plant Sci. 2003, 8, 484–491. [Google Scholar] [CrossRef] [PubMed]
- Keller, B.; Feuillet, C.; Yahiaoui, N. Map-based isolation of disease resistance genes from bread wheat: Cloning in a supersize genome. Genet. Res. 2005, 85, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Sui, F.; Zhao, D.; Zhu, H.; Gong, Y.; Tang, Z.; Huang, X.Y.; Zhang, G.; Zhao, F.J. Map-based cloning of a new total loss-of-function allele of OsHMA3 causes high cadmium accumulation in rice grain. J. Exp. Bot. 2019, 70, 2857–2871. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Yao, F.J.; Lu, L.X.; Fang, M.; Zhang, Y.M.; Khan, A.A.; Kong, X.H.; Yu, J.; Jiang, W.Z.; Kitamoto, Y.; et al. Map-based cloning of genes encoding key enzymes for pigment synthesis in Auricularia cornea. Fungal Biol. 2019, 123, 843–853. [Google Scholar] [CrossRef]
- Liu, S.; Yeh, C.T.; Tang, H.M.; Nettleton, D.; Schnable, P.S. Gene mapping via bulked segregant RNA-Seq (BSR-Seq). PLoS ONE 2012, 7, e36406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein, H.; Xiao, Y.; Conklin, P.A.; Govindarajulu, R.; Kelly, J.A.; Scanlon, M.J.; Whipple, C.J.; Bartlett, M. Bulked-segregant analysis coupled to whole genome sequencing (BSA-Seq) for rapid gene cloning in Maize. G3 2018, 8, 3583–3592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, W.; Shi, J.; Qiu, C.; Wang, Y.; Rehman, S.; Yu, S.; Huang, S.; He, C.; Wang, W.; Chen, H.; et al. Identification of a genomic region controlling thermotolerance at flowering in maize using a combination of whole genomic re-sequencing and bulked segregant analysis. Theor. Appl. Genet. 2020, 133, 2797–2810. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Dai, G.; Zhou, W.; Liang, H.; Huang, J.; Qing, D.; Chen, W.; Wu, H.; Yang, X.; Li, D.; et al. Mapping and identifying a candidate gene Plr4, a recessive gene regulating purple leaf in Rice, by using bulked segregant and transcriptome analysis with next-generation sequencing. Int. J. Mol. Sci. 2019, 20, 4335. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Su, T.; Zhao, X.; Wang, W.; Zhang, D.; Yu, Y.; Bayer, P.E.; Edwards, D.; Yu, S.; Zhang, F. Assembly of the non-heading pak choi genome and comparison with the genomes of heading Chinese cabbage and the oilseed yellow sarson. Plant Biotechnol. J. 2021, 19, 966–976. [Google Scholar] [CrossRef]
- Jain, M.; Olsen, H.E.; Paten, B.; Akeson, M. The Oxford Nanopore MinION: Delivery of nanopore sequencing to the genomics community. Genome Biol. 2016, 17, 1–11. [Google Scholar]
- Azibi, T.; Hadj-Arab, H.; Lode, M.; Ferreira de Carvalho, J.; Trotoux, G.; Negre, S.; Gilet, M.M.; Boutte, J.; Lucas, J.; Vekemans, X.; et al. Impact of whole genome triplication on the evolutionary history and the functional dynamics of regulatory genes involved in Brassica self-incompatibility signalling pathway. Plant Reprod. 2020, 33, 43–58. [Google Scholar] [CrossRef]
- Guo, Y.L.; Zhao, X.; Lanz, C.; Weigel, D. Evolution of the S-locus region in Arabidopsis relatives. Plant Physiol. 2011, 157, 937–946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dou, S.; Zhang, T.; Tu, J.; Shen, J.; Yi, B.; Wen, J.; Fu, T.; Dai, C.; Ma, C. Generation of novel self-incompatible Brassica napus by CRISPR/Cas9. Plant Biotechnol. J. 2021, 19, 875–877. [Google Scholar] [CrossRef] [PubMed]
- Jung, H.J.; Jung, H.J.; Ahmed, N.U.; Park, J.I.; Kang, K.K.; Hur, Y.; Lim, Y.P.; Nou, I.S. Development of self-compatible B. rapa by RNAi-mediated S locus gene silencing. PLoS ONE 2012, 7, e49497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; Van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, Y.; Fujimoto, R.; Toriyama, K.; Nishio, T. Commonality of self-recognition specificity of S haplotypes between Brassica oleracea and Brassica rapa. Plant Mol. Biol. 2003, 52, 617–626. [Google Scholar] [CrossRef]
- Ma, R.; Han, Z.; Hu, Z.; Lin, G.; Gong, X.; Zhang, H.; Nasrallah, J.B.; Chai, J. Structural basis for specific self-incompatibility response in Brassica. Cell Res. 2016, 26, 1320–1329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murasec, K.; Moriwaki, Y.; Mori, T.; Liu, X.; Masaka, C.; Takada, Y.; Maesaki, R.; Mishima, M.; Fujii, S.; Hirano, Y.; et al. Mechanism of self/nonself-discrimination in Brassica self-incompatibility. Nat. Commun. 2020, 11, 4916. [Google Scholar] [CrossRef] [PubMed]
- Nishio, T.; Hinata, K. Stigma proteins in self-incompatible Brassica campestris L. and self-compatible relatives, with special reference to S-allele specificity. Genes Genet. Syst. 1978, 53, 27–33. [Google Scholar] [CrossRef] [Green Version]
- Walker, J.C.; Zhang, R. Relationship of a putative receptor protein kinase from maize to the S-locus glycoproteins of Brassica. Nature 1990, 345, 743–746. [Google Scholar] [CrossRef] [PubMed]
- Schopfer, C.R.; Nasrallah, M.E.; Nasrallah, J.B. The male determinant of self-incompatibility in Brassica. Science 1999, 286, 1697–1700. [Google Scholar] [CrossRef]
- The Brassica rapa Genome Sequencing Project Consortium; Wang, X.; Wang, H.; Wang, J.; Sun, R.; Wu, J.; Liu, S.; Bai, Y.; Mun, J.H.; Bancroft, I.; et al. The genome of the mesopolyploid crop species Brassica rapa. Nat. Genet. 2011, 43, 1035–1039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, F.; Sun, R.; Hou, X.; Zheng, H.; Zhang, F.; Zhang, Y.; Liu, B.; Liang, J.; Zhuang, M.; Liu, Y.; et al. Subgenome parallel selection is associated with morphotype diversification and convergent crop domestication in Brassica rapa and Brassica oleracea. Nat. Genet. 2016, 48, 1218–1224. [Google Scholar] [CrossRef] [PubMed]
- Murray, M.; Thompson, W.F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Res. 1980, 8, 4321–4326. [Google Scholar] [CrossRef] [Green Version]
- Nishio, T.; Kusaba, M.; Watanabe, M.; Hinata, K. Registration of S alleles in Brassica campestris L. by the restriction fragment sizes of SLGs. Theor. Appl. Genet. 1996, 92, 388–394. [Google Scholar] [CrossRef] [PubMed]
- Nishio, T.; Kusaba, M.; Sakamoto, K.; Ockendon, D. Polymorphism of the kinase domain of the S-locus receptor kinase gene (SRK) in Brassica oleracea L. Theor. Appl. Genet. 1997, 95, 335–342. [Google Scholar] [CrossRef]
- Goodwin, S.; Gurtowski, J.; Ethe-Sayers, S.; Deshpande, P.; Schatz, M.C.; McCombie, W.R. Oxford Nanopore sequencing, hybrid error correction, and de novo assembly of a eukaryotic genome. Genome Res. 2015, 25, 1750–1756. [Google Scholar] [CrossRef] [Green Version]
- Jayakumar, V.; Sakakibara, Y. Comprehensive evaluation of non-hybrid genome assembly tools for third-generation PacBio long-read sequence data. Brief. Bioinform. 2019, 20, 866–876. [Google Scholar] [CrossRef] [PubMed]
- Phillippy, A.M. New advances in sequence assembly. Genome Res. 2017, 27, xi–xiii. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salzberg, S.L.; Phillippy, A.M.; Zimin, A.; Puiu, D.; Magoc, T.; Koren, S.; Treangen, T.J.; Schatz, M.C.; Delcher, A.L.; Roberts, M. GAGE: A critical evaluation of genome assemblies and assembly algorithms. Genome Res. 2012, 22, 557–567. [Google Scholar] [CrossRef] [Green Version]
- Li, H. Minimap2: Pairwise alignment for nucleotide sequences. Bioinformatics 2018, 34, 3094–3100. [Google Scholar] [CrossRef] [PubMed]
- Vaser, R.; Sović, I.; Nagarajan, N.; Šikić, M. Fast and accurate de novo genome assembly from long uncorrected reads. Genome Res. 2017, 27, 737–746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, B.J.; Abeel, T.; Shea, T.; Priest, M.; Abouelliel, A.; Sakthikumar, S.; Cuomo, C.A.; Zeng, Q.; Wortman, J.; Young, S. Pilon: An integrated tool for comprehensive microbial variant detection and genome assembly improvement. PLoS ONE 2014, 9, e112963. [Google Scholar] [CrossRef]
- Price, A.L.; Jones, N.C.; Pevzner, P.A. De novo identification of repeat families in large genomes. Bioinformatics 2005, 21 (Suppl. S1), i351–i358. [Google Scholar] [CrossRef] [Green Version]
- Jurka, J.; Kapitonov, V.V.; Pavlicek, A.; Klonowski, P.; Kohany, O.; Walichiewicz, J. Repbase Update, a database of eukaryotic repetitive elements. Cytogenet. Genome Res. 2005, 110, 462–467. [Google Scholar] [CrossRef]
- Chen, N. Using RepeatMasker to identify repetitive elements in genomic sequences. Curr. Protoc. Bioinform. 2004, 5, 4.10.1–4.10.14. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C.; Myers, E.W. PILER: Identification and classification of genomic repeats. Bioinformatics. 2005, 21 (Suppl. S1), i152–i158. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Wang, H. LTR_FINDER: An efficient tool for the prediction of full-length LTR retrotransposons. Nucleic Acids Res. 2007, 35 (Suppl. S2), W265–W268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kent, W.J. BLAT-the BLAST-like alignment tool. Genome Res. 2002, 12, 656–664. [Google Scholar]
- Stanke, M.; Keller, O.; Gunduz, I.; Hayes, A.; Waack, S.; Morgenstern, B. AUGUSTUS: Ab initio prediction of alternative transcripts. Nucleic Acids Res. 2006, 34 (Suppl. S2), W435–W439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holt, C.; Yandell, M. MAKER2: An annotation pipeline and genome-database management tool for second-generation genome projects. BMC Bioinform. 2011, 12, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Lowe, T.M.; Eddy, S.R. tRNAscan-SE: A program for improved detection of transfer RNA genes in genomic sequence. Nucleic Acids Res. 1997, 25, 955–964. [Google Scholar] [CrossRef]
- Griffiths-Jones, S.; Moxon, S.; Marshall, M.; Khanna, A.; Eddy, S.R.; Bateman, A. Rfam: Annotating non-coding RNAs in complete genomes. Nucleic Acids Res. 2005, 33 (Suppl. S1), D121–D124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, T.; Gao, C.; Yue, Y.; Liu, Z.; Ma, C.; Zhou, G.; Yang, Y.; Duan, Z.; Li, B.; Wen, J.; et al. Time-course transcriptome analysis of compatible and incompatible pollen-stigma interactions in Brassica napus L. Front. Plant Sci. 2017, 8, 682. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547. [Google Scholar] [CrossRef] [PubMed]
- Marçais, G.; Delcher, A.L.; Phillippy, A.M.; Coston, R.; Salzberg, S.L.; Zimin, A. MUMmer4: A fast and versatile genome alignment system. PLoS Comput. Biol. 2018, 14, e1005944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, H.; Zhang, X.; Miao, C.; Zhang, J.; Ming, R.; Schnable, J.C.; Schnable, P.S.; Lyons, E.; Lu, J. ALLMAPS: Robust scaffold ordering based on multiple maps. Genome Biol. 2015, 16, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Ma, C.; Tang, J.; Tang, W.; Tu, J.; Shen, J.; Fu, T. Distribution of S haplotypes and its relationship with restorer-maintainers of self-incompatibility in cultivated Brassica napus. Theor. Appl. Genet. 2008, 117, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Chen, C.; Zhou, G.; Geng, C.; Ma, C.; Tu, J.; Fu, T. Genetic analysis of four self-incompatible lines in Brassica napus. Plant Breed. 2001, 120, 57–61. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Material | Genotype | SI/SC | Expected | χ2 | ||
|---|---|---|---|---|---|---|
| I | I/II | II | Phenotype | Ratio | ||
| 325 | 0 | 0 | 12 | 0/12 | ||
| 326 | 12 | 0 | 0 | 12/0 | ||
| 325 × 326 | 0 | 12 | 0 | 12/0 | ||
| 326 × 325 | 0 | 12 | 0 | 12/0 | ||
| (325 × 326)F2 | 20 | 50 | 22 | 70/22 | 3:1 | 0.094 < 3.84 |
| 325 | ZYCX | Chiifu | Z1 | |
|---|---|---|---|---|
| Assembly feature | ||||
| Total assembly size (Mb) | 376.69 | 370.42 | 353.14 | 401.92 |
| Contig number | 314 | 1985 | 1498 | 1037 |
| Contig N50 (Mb) | 4.54 | 2.82 | 1.45 | 2.27 |
| Longest length (Mb) | 22.32 | 22.37 | 9.42 | 22.13 |
| Genome annotation | ||||
| Gene model | 42500 | 45363 | 46250 | 46721 |
| Percentage of anchored genes (%) | 0.9691 | 0.985 | 0.9858 | 0.9814 |
| Accession | Phenotype | S haplotype | Alignment Size (kb) | S locus Size (kb) | Distance 1 a (kb) | Distance 2 b (kb) | Distance 3 c (kb) | SCR Length (bp) | SRK Length (bp) | SLG Length (bp) | TE Type | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 325 | SC | BrS-325 | 145.991 | 67.777 | 41.173 | 19.706 | 15.384 | 391 | 5618 d | 6083 e | 4122 | DNA-TE |
| Z1 | SC | BrS-60 | 147.921 | 54.448 | 24.828 | 6.575 | 11.533 | 378 | - | 6720 | 1966 | LTR-TE |
| Chiifu | SI | BrS-60 | 155.427 | 73.764 | 41.492 | 6.582 | 26.943 | 378 | - | 7967 | 1459 | LTR-TE |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, B.; Zhang, X.; Liu, Z.; Wang, L.; Song, L.; Liang, X.; Dou, S.; Tu, J.; Shen, J.; Yi, B.; et al. Genetic and Molecular Characterization of a Self-Compatible Brassica rapa Line Possessing a New Class II S Haplotype. Plants 2021, 10, 2815. https://doi.org/10.3390/plants10122815
Li B, Zhang X, Liu Z, Wang L, Song L, Liang X, Dou S, Tu J, Shen J, Yi B, et al. Genetic and Molecular Characterization of a Self-Compatible Brassica rapa Line Possessing a New Class II S Haplotype. Plants. 2021; 10(12):2815. https://doi.org/10.3390/plants10122815
Chicago/Turabian StyleLi, Bing, Xueli Zhang, Zhiquan Liu, Lulin Wang, Liping Song, Xiaomei Liang, Shengwei Dou, Jinxing Tu, Jinxiong Shen, Bin Yi, and et al. 2021. "Genetic and Molecular Characterization of a Self-Compatible Brassica rapa Line Possessing a New Class II S Haplotype" Plants 10, no. 12: 2815. https://doi.org/10.3390/plants10122815