
The Development of New Species-Specific Molecular Markers Based on 5S rDNA in Elaeagnus L. Species


Abstract
:1. Introduction
2. Results
2.1. Alignments of 5S rDNA Non-Transcribed Spacers (NTSs) among the Elaeagnus Species and Development of Species-Specific Primers
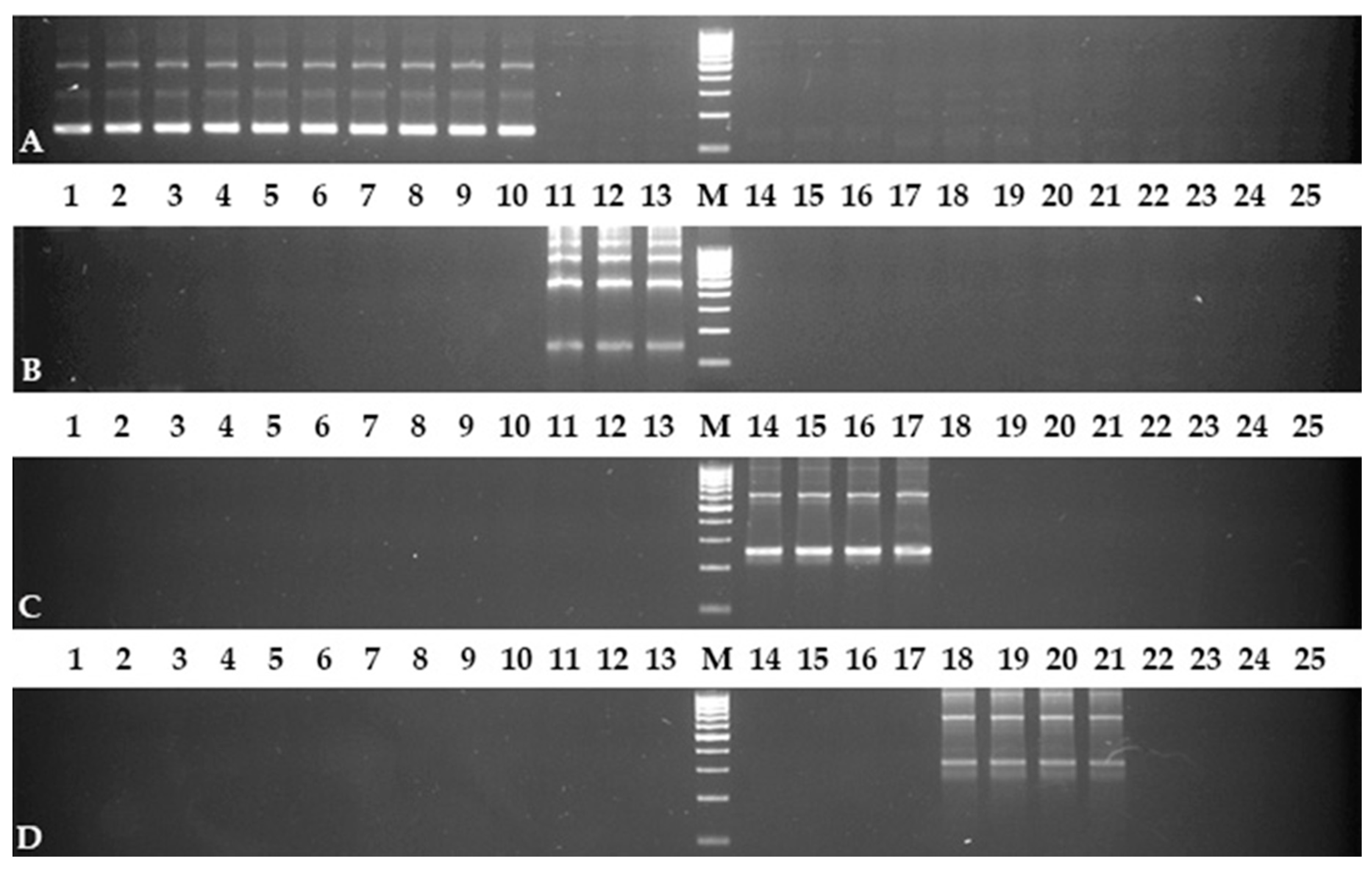
2.2. PCR Test with Developed Primers
3. Discussion
4. Materials and Methods
4.1. Plant Material and DNA Isolation
4.2. Analysis of Sequences and Primer Design
4.3. The PCR Experiments and Electrophoresis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bartish, I.V.; Swenson, U. Elaeagnaceae. In The Families and Genera of Vascular Plants; Kubitzki, K., Ed.; Springer: Berlin/Heidelberg, Germany, 2004; pp. 131–134. [Google Scholar]
- Raunkiaer, C. The Life Forms of Plants and Statistical Geography; The Claderon Press: Oxford, UK, 1934; p. 623. [Google Scholar]
- Gardner, I.C. Nitrigen fixation in Elaeagnus root nodules. Nature 1958, 181, 717–718. [Google Scholar] [CrossRef]
- Patel, S. Plant genus Elaeagnus: Underutilized lycopene and linoleic acid reserve with permaculture potential. Fruits 2015, 70, 191–199. [Google Scholar] [CrossRef] [Green Version]
- Mao, Y.; Hu, Q.; Zhang, M.; Yang, L.; Zhang, L.; Wang, Y.; Yin, Y.; Pang, H.; Liu, Y.; Su, X.; et al. Genome analysis and Hi-C assisted assembly of Elaeagnus angustifolia L., a deciduous tree belonging to Elaeagnaceae. iScience 2019. [Google Scholar] [CrossRef]
- Choi, K.S.; Son, O.; Park, S. The Chloroplast genome of Elaeagnus macrophylla and trnH duplication event in Elaeagnaceae. PLoS ONE 2015, 10, e0138727. [Google Scholar] [CrossRef]
- Liu, J.; Gong, L.D.; Qi, L.; Liu, Z.Y.; Niu, Y.F.; Shi, C. The complete chloroplast genome of Elaeagnus conferta Roxb (Elaeagnaceae). Mitochondrial DNA B Resour. 2019, 4, 2035–2036. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.P.; Fan, C.Z.; Zhu, J.; Li, X.J. Identification of original plants of uyghur medicinal materials fructus elaeagni using morphological characteristics and DNA barcode. J. Chin. Mater. Med. 2014, 39, 2216–2221. [Google Scholar]
- Tagane, S.; Van Son, D.; Rueangruea, S.; Suddee, S.; Chhang, P.; Toyama, H.; Yahara, T. Elaeagnus elongatus Tagane & V.S.Dang (Elaeagnaceae), a new species from Cambodia and Thailand. Thai Forest Bull. Bot. 2015, 43, 30–35. [Google Scholar]
- Burgess, K.S.; Fazekas, A.J.; Kesanakurti, P.R.; Graham, S.W.; Husband, B.C.; Newmaster, S.G.; Percy, D.M.; Hajibabaei, M.; Barrett, S.C.H. Discriminating plant species in a local temperate flora using the rbcL + matK DNA barcode. Methods Ecol. Evol. 2011, 2, 333–340. [Google Scholar] [CrossRef]
- Son, O.; Yoon, C.Y.; Park, S.J. Phylogenetic relationships in Korean Elaeagnus L. based on nrDNA ITS sequences. Korean J. Plant. Res. 2014, 27, 671–679. [Google Scholar] [CrossRef] [Green Version]
- Asadiar, L.S.; Rahmani, F.; Siami, A. Assessment of genetic variation in Russian olive (Elaeagnus angustifolia) based on morphological traits and random amplified polymorphic DNA (RAPD) genetic markers. J. Med. Plant. Res. 2012, 6, 1652–1661. [Google Scholar]
- Asadiar, L.S.; Rahmani, F.; Siami, A. Assessment of genetic diversity in the Russian olive (Elaeagnus angustifolia) based on ISSR genetic markers. Rev. Ciênc. Agron. 2013, 44, 310–316. [Google Scholar] [CrossRef] [Green Version]
- Zirak, R.; Soleimani, A.; Zeinolabedini, M.; Maleki, H.H.; Kheiri, A. Morphological and AFLP-based genetic diversity assessment of Elaeagnus angustifolia L. J. Plant. Genet. Res. 2018, 5, 41–54. [Google Scholar]
- Uzun, A.; Yigit, M.A.; Pinar, H.; Yaman, M.; Arslan, S. Molecular characterization of Elaeagnus angustifolia L. genotypes collected from different parts of Turkey. Int. J. Agric. Nat. Sci. 2019, 1, 10–13. [Google Scholar]
- Wang, Y.; Ma, Y.; Jia, B.; Wu, Q.; Zang, D.; Yu, X. Analysis of the genetic diversity of the coastal and island endangered plant species Elaeagnus macrophylla via conserved DNA-derived polymorphism marker. PeerJ 2020, 8, e8498. [Google Scholar] [CrossRef] [Green Version]
- Alexandrov, O.S.; Razumova, O.V.; Karlov, G.I. A Comparative study of 5S rDNA non-transcribed spacers in Elaeagnaceae species. Plants 2021, 10, 4. [Google Scholar] [CrossRef] [PubMed]
- Martins, C.; Wasko, A.P. Organization and evolution of 5S ribosomal DNA in the fish genome. Chapter X. In Focus on Genome Research; Clyde, R., Ed.; Williams: Nova Science Publishers: New York, NY, USA, 2004; pp. 335–363. [Google Scholar]
- Gerbi, S.A. Evolution of ribosomal DNA. In Molecular Evolutionary Genetics; MacIntyre, R.J., Ed.; Plenum Publishing: New York, NY, USA, 1985; pp. 419–517. [Google Scholar]
- Avadhani, M.N.M.; Selvaraj, C.I.; Tharachand, C.; Rajasekharan, P.E. Molecular characterization of medicinal and aromatic plants by 5S rRNA NTS and PCR RFLP—A mini review. Res. Biotechnol. 2012, 3, 41–48. [Google Scholar]
- Fernández-Pérez, J.; Nantón, A.; Méndez, J. Sequence characterization of the 5S ribosomal DNA and the internal transcribed spacer (ITS) region in four European Donax species (Bivalvia: Donacidae). BMC Genet. 2018, 19, 97. [Google Scholar] [CrossRef]
- Alexandrov, O.S.; Divashuk, M.G.; Karlov, G.I. Development of the St/J/V-genome specific molecular marker on basis of 5S-rDNA polymorphism. Mosc. Univ. Biol. Sci. Bull. 2018, 73, 18–23. [Google Scholar] [CrossRef]
- Alexandrov, O.S. Creating of CAPS-marker system for identification of J, V and St subgenomes among species of Triticae tribe. In Proceedings of the International scientific and practical conference “Vavilovskie chteniya—2018”, Dedicated to the 131st Anniversary of the Birth of N.I. Vavilov, Saratov, Russsia, 28–29 November 2018; Kuznetsov, N.I., Vorotnikov, I.L., Sh’urova, N.A., Eds.; Amirit: Saratov, Russia, 2018. (In Russian). [Google Scholar]
- Alexandrov, O.S.; Karlov, G.I. Development of 5S rDNA-based molecular markers for the identification of Populus deltoides Bartr. ex Marshall, Populus nigra L., and their hybrids. Forests 2018, 9, 604. [Google Scholar] [CrossRef] [Green Version]
- Alexandrov, O.S.; Karlov, G.I. The development of Populus alba L. and Populus tremula L. species specific molecular markers based on 5S rDNA non-transcribed spacer polymorphism. Forests 2019, 10, 1092. [Google Scholar] [CrossRef] [Green Version]
- Malhotra, K.; Foltz, L.; Mahoney, W.C.; Schueler, P.A. Interaction and effect of annealing temperature on primers used in differential display RT-PCR. Nucleic Acids Res. 1998, 26, 854–856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorenz, T.C. Polymerase chain reaction: Basic protocol plus troubleshooting and optimization strategies. J. Vis. Exp. 2012, 63, e3998. [Google Scholar] [CrossRef] [PubMed]
- Handell-Mazzetti, H.R.E. Elaeagnus umbellata var. siphonantha (Nakai) Hand.-Mazz. Symb. Sin. 1933, 7, 540. [Google Scholar]
- Doyle, J.J.; Doyle, J.L. Isolation of plant DNA from fresh tissue. Focus 1990, 12, 13–15. [Google Scholar]
- Alexandrov, O.S. Study of the upstream ricin gene sequences in different castor (Ricinus communis) varieties as a preliminary step in CRISPR/Cas9 editing. Res. Crops 2020, 21, 344–348. [Google Scholar]
- GeneDoc: Analysis and Visualization of Genetic Variation. Available online: http://www.nrbsc.org/gfx/genedoc/ebinet.htm (accessed on 1 November 2021).


{kind=link}
| Primer Name | Sequence | Annealing, t °C 1 | PCR Product Length, bp |
|---|---|---|---|
| Elang1-f | 5′-TCGATCAACCGAATCAAACA-3′ | 59.0 | 150 |
| Elang1-r | 5′-CGAAACTTGTTATTTTTGCGAAT-3′ | ||
| Elcom1-f | 5′-TCTAATCCGATAAACCGAATTGT-3′ | 62.0 | 151 |
| Elcom1-r | 5′-AAAGTATTTCATGCGTGCGTAA-3′ | ||
| Elcom2-f | 5′-GCTCTATTTTATTCTAATCCGAT-3′ | 62.0 | 165 |
| Elcom2-r | 5′-GATTTGTACGGGTAAAGTATTT-3′ | ||
| Elpung1-f | 5′-GTGTAAGTAGAAAGTTGGAAAC-3′ | 54.0 | 253 |
| Elpung1-r | 5′-AACTCTTCTTATGATTTGGTT-3′ | ||
| Elmult1-f | 5′-GGATGGGTGACCTCCGG-3′ | 55.0 | 328 |
| Elmul1-r | 5′-AACGTTCTACATGCATTCGT-3′ | ||
| Elmult1-f | 5′-GGATGGGTGACCTCCGG-3′ | 55.0 | 328 |
| Elmult2-r | 5′-GGCGACCCTGGGAAGTGT-3′ | ||
| Elmult1-f | 5′-GGATGGGTGACCTCCGG-3′ | 55.0 | 329 |
| Elmult3-r | 5′-TAACGTTCTACATGCATTCG-3′ | ||
| Elmult2-f | 5′-GCTACATCATCAGTCCAACA-3′ | 55.0 | 103 |
| Elmult1-r | 5′-AACGTTCTACATGCATTCGT-3′ | ||
| Elmult2-f | 5′-GCTACATCATCAGTCCAACA-3′ | 55.0 | 103 |
| Elmult2-r | 5′-GGCGACCCTGGGAAGTGT-3′ | ||
| Elmult2-f | 5′-GCTACATCATCAGTCCAACA-3′ | 55.0 | 104 |
| Elmult3-r | 5′-TAACGTTCTACATGCATTCG-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alexandrov, O.S.; Karlov, G.I. The Development of New Species-Specific Molecular Markers Based on 5S rDNA in Elaeagnus L. Species. Plants 2021, 10, 2713. https://doi.org/10.3390/plants10122713
Alexandrov OS, Karlov GI. The Development of New Species-Specific Molecular Markers Based on 5S rDNA in Elaeagnus L. Species. Plants. 2021; 10(12):2713. https://doi.org/10.3390/plants10122713
Chicago/Turabian StyleAlexandrov, Oleg S., and Gennady I. Karlov. 2021. "The Development of New Species-Specific Molecular Markers Based on 5S rDNA in Elaeagnus L. Species" Plants 10, no. 12: 2713. https://doi.org/10.3390/plants10122713