
TaGRF3-2A Improves Some Agronomically Valuable Traits in Semi-Dwarf Spring Triticale

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
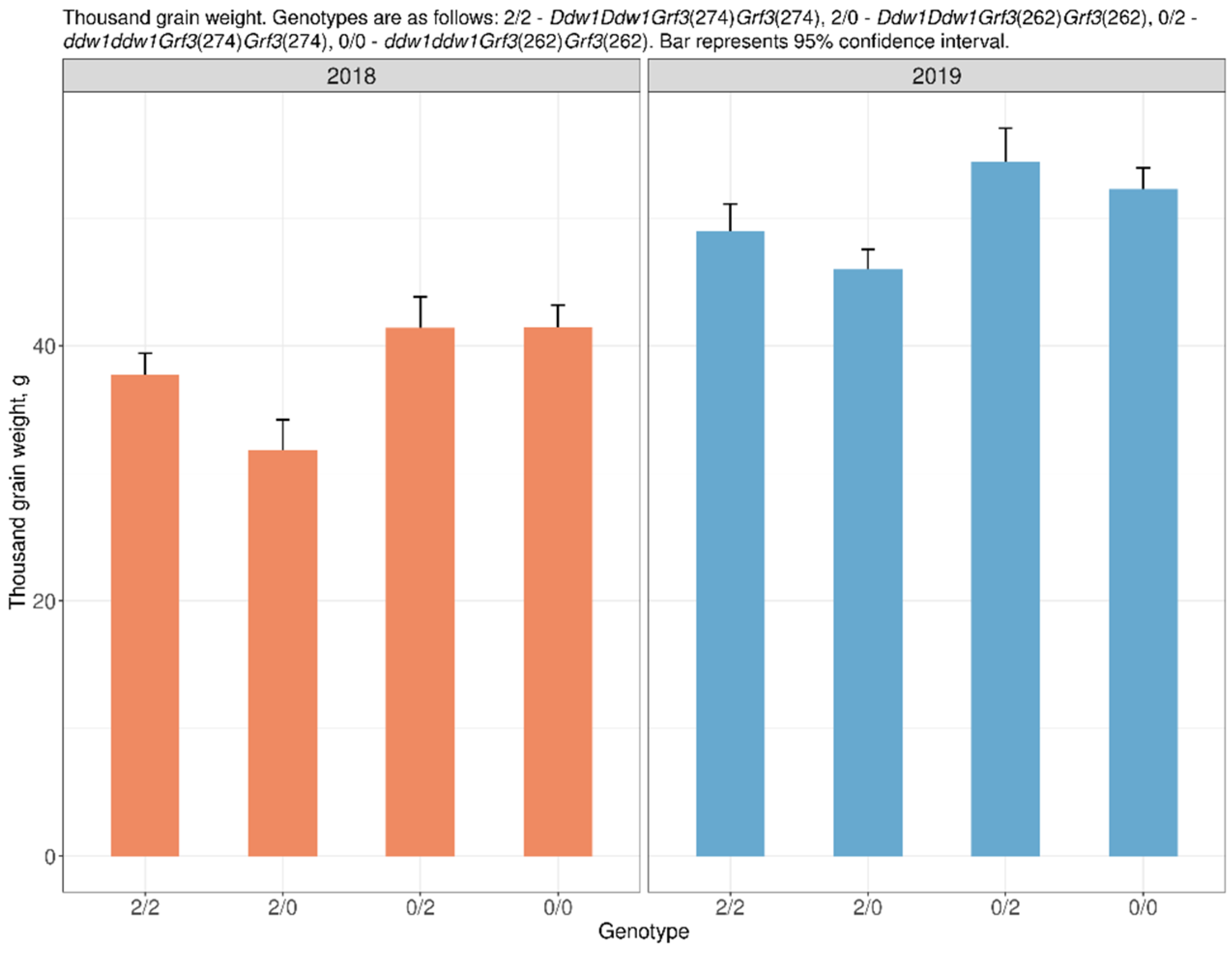
2.1. Spike Architecture and Productivity
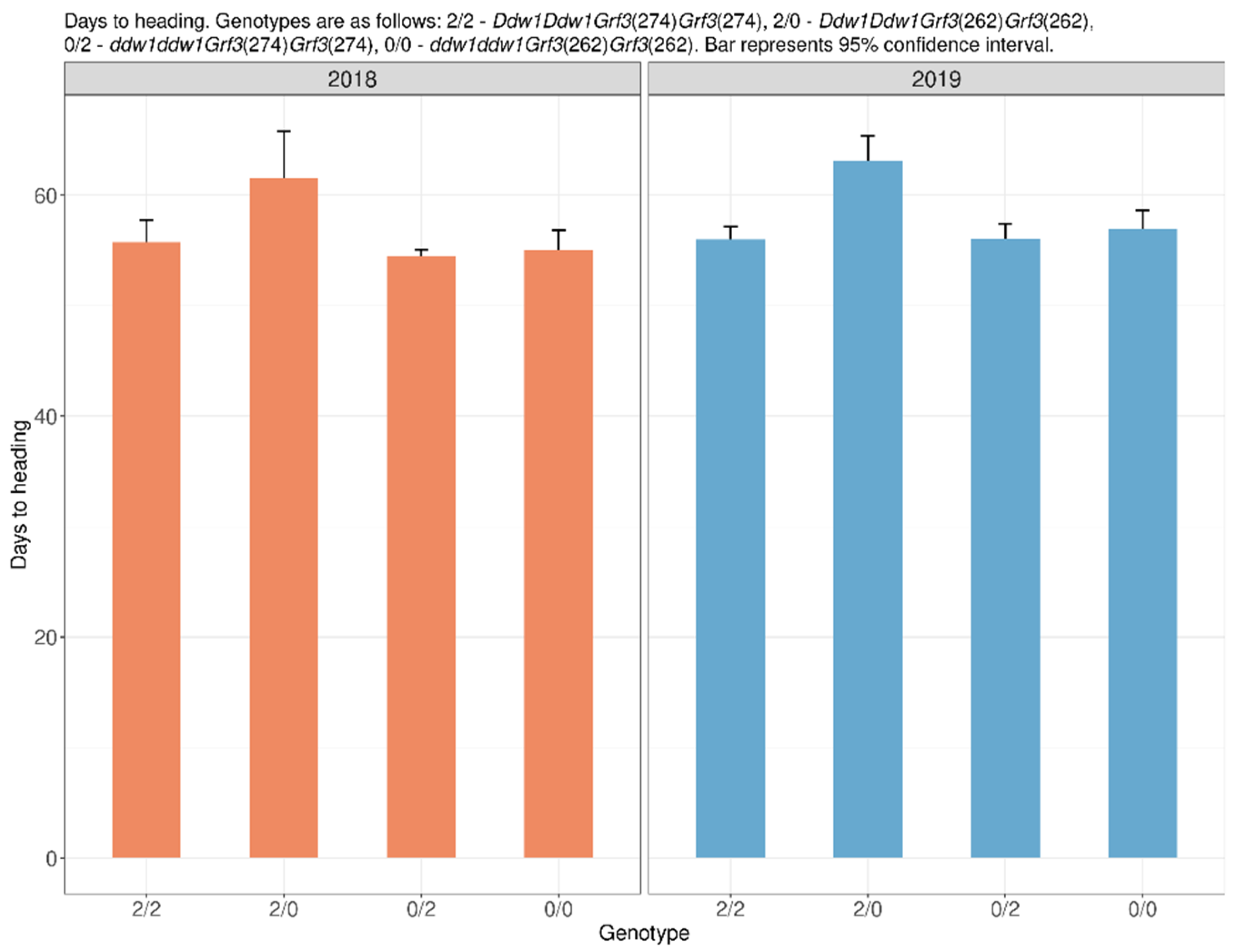
2.2. Flowering and Heading
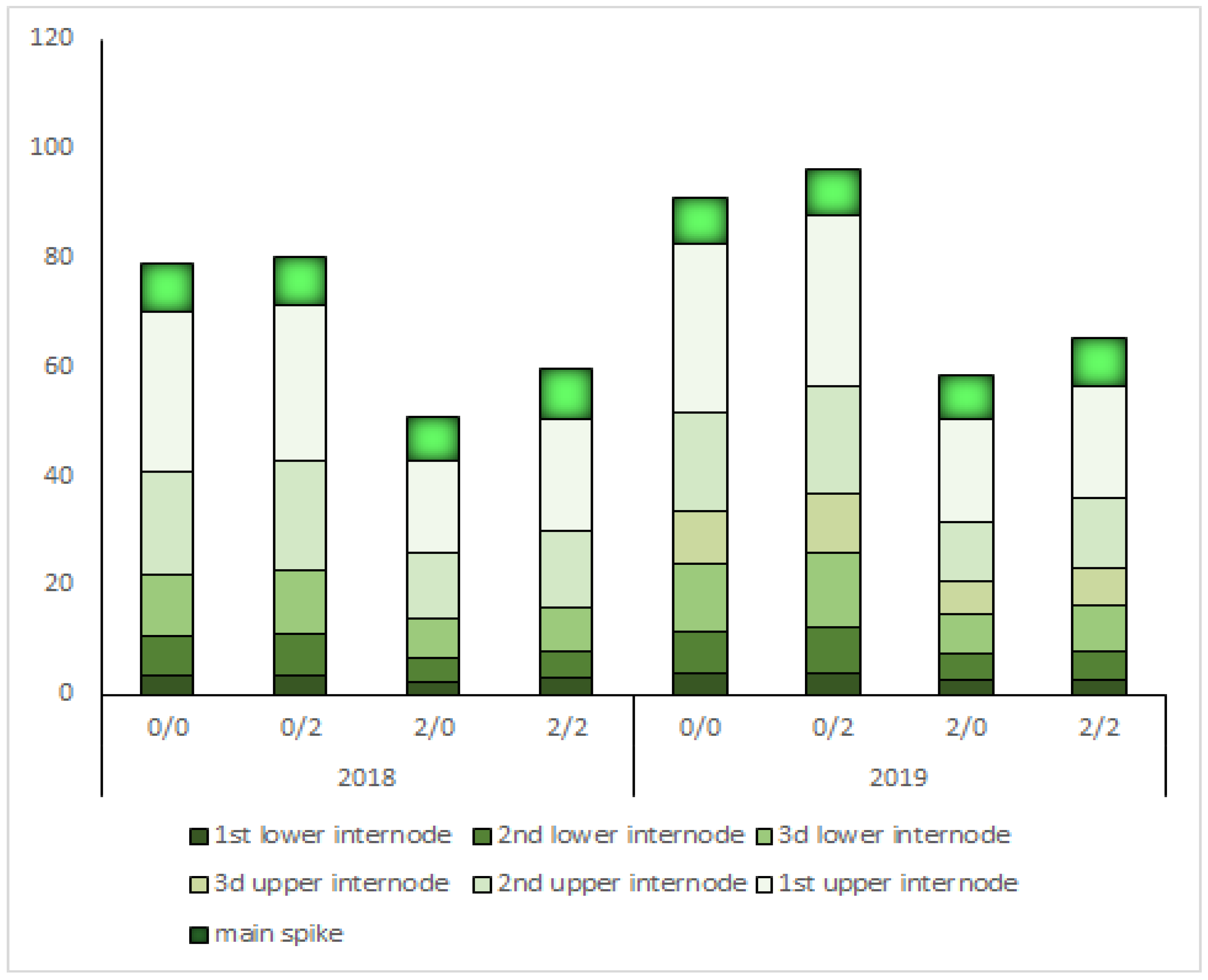
2.3. Plant Height, Internodes, and Main Spike Length
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Field Experiment
4.3. Plant Structural Analysis
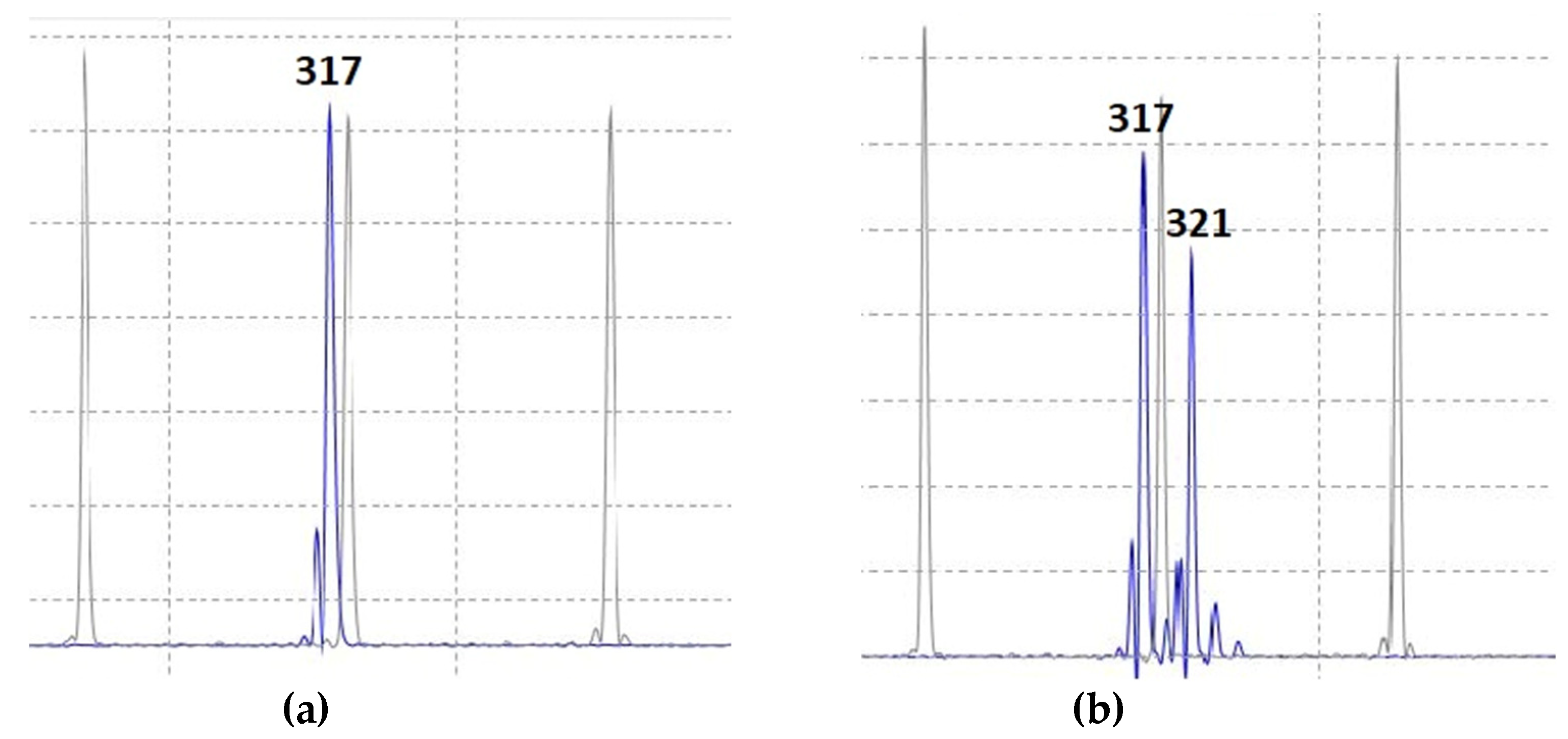
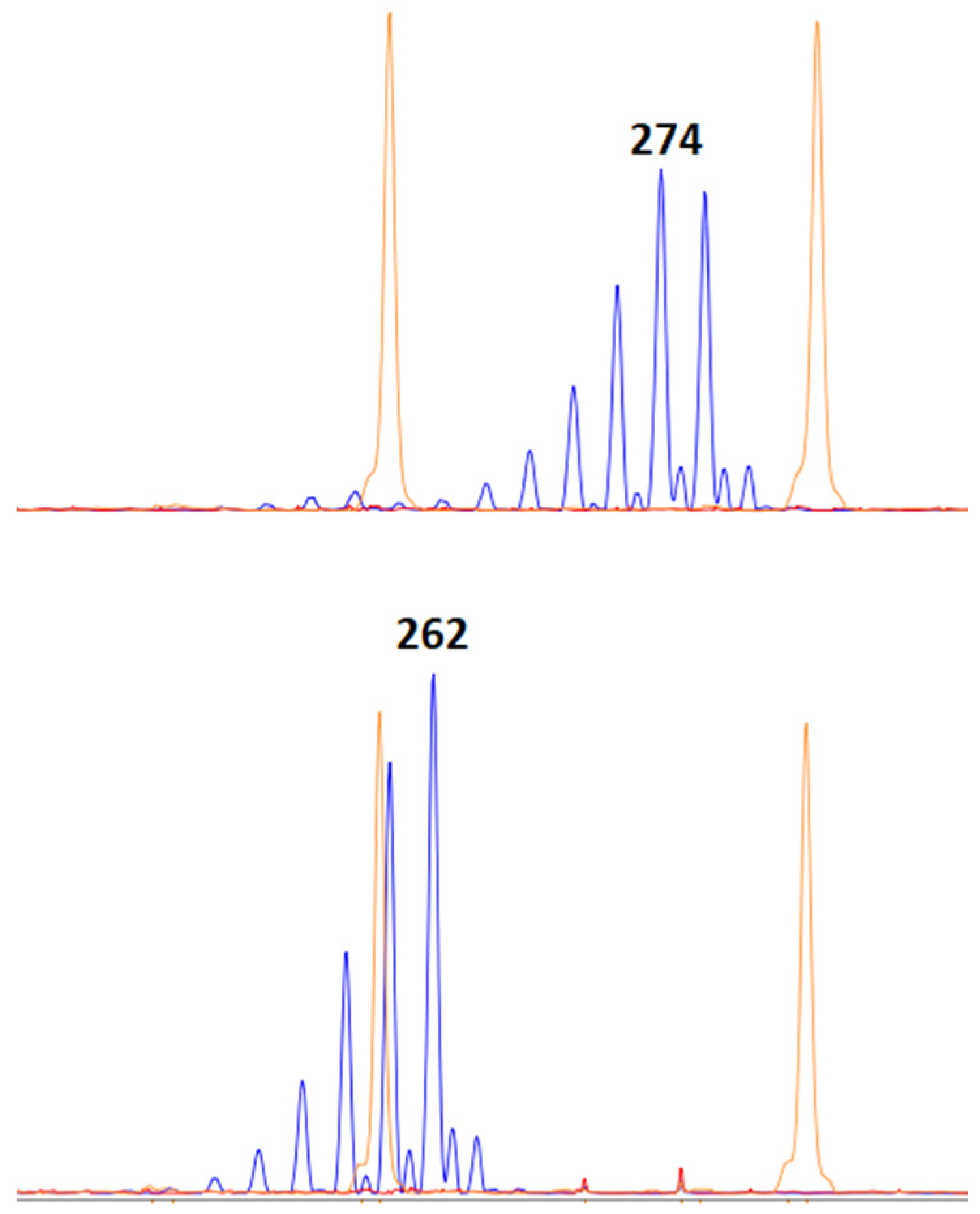
4.4. Plant Genotyping
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Sherman, J.F.; Young, E.O.; Cavadini, J. Tillage and Liquid Dairy Manure Effects on Overland Flow Nitrogen and Phosphorus Loss Potential in an Upper Midwest Corn Silage-Winter Triticale Cropping System. Agronomy 2021, 11, 1775. [Google Scholar] [CrossRef]
- Bartosiak, S.F.; Arseniuk, E.; Szechyńska-Hebda, M.; Bartosiak, E. Monitoring of Natural Occurrence and Severity of Leaf and Glume Blotch Diseases of Winter Wheat and Winter Triticale Incited by Necrotrophic Fungi Parastagonospora spp. and Zymoseptoria tritici. Agronomy 2021, 11, 967. [Google Scholar] [CrossRef]
- Góral, T.; Wiśniewska, H.; Ochodzki, P.; Twardawska, A.; Walentyn-Góral, D. Resistance to Fusarium Head Blight, Kernel Damage, and Concentration of Fusarium Mycotoxins in Grain of Winter Triticale (x Triticosecale Wittmack) Lines. Agronomy 2021, 11, 16. [Google Scholar] [CrossRef]
- Schillinger, W.F.; Archer, D.W. Winter Triticale: A Long-Term Cropping Systems Experiment in a Dry Mediterranean Climate. Agronomy 2020, 10, 1777. [Google Scholar] [CrossRef]
- Yerzhebayeva, R.S.; Bazylova, T.A.; Babissekova, D.I.; Amangeldiyeva, A.A.; Tajibayev, D.G.; Ydyrys, A. Studying a Spring Triticale Collection for Resistance to Leaf and Stem Rusts using Allele-Specific Markers. Cytol. Genet. 2020, 54, 546–554. [Google Scholar] [CrossRef]
- Skowrońska, R.; Tomkowiak, A.; Nawracała, J.; Kwiatek, M. Molecular identification of slow rusting resistance Lr46/Yr29 gene locus in selected triticale (× Triticosecale Wittmack) cultivars. J. Appl. Genet. 2020, 61, 359–366. [Google Scholar] [CrossRef]
- Rajičić, V.; Popović, V.; Perišić, V.; Biberdžić, M.; Jovović, Z.; Gudžić, N.; Mihailović, V.; Čolić, V.; Đurić, N.; Terzić, D. Impact of Nitrogen and Phosphorus on Grain Yield in Winter Triticale Grown on Degraded Vertisol. Agronomy 2020, 10, 757. [Google Scholar] [CrossRef]
- Bielski, S.; Romaneckas, K.; Šarauskis, E. Impact of Nitrogen and Boron Fertilization on Winter Triticale Productivity Parameters. Agronomy 2020, 10, 279. [Google Scholar] [CrossRef] [Green Version]
- Wójcik-Gront, E.; Studnicki, M. Long-Term Yield Variability of Triticale (×Triticosecale Wittmack) Tested Using a CART Model. Agriculture 2021, 11, 92. [Google Scholar] [CrossRef]
- Klikocka, H.; Kasztelan, A.; Zakrzewska, A.; Wyłupek, T.; Szostak, B.; Skwaryło-Bednarz, B. The Energy Efficiency of the Production and Conversion of Spring Triticale Grain into Bioethanol. Agronomy 2019, 9, 423. [Google Scholar] [CrossRef] [Green Version]
- Sestili, F.; Margiotta, B.; Vaccino, P.; Moscaritolo, S.; Giorgi, D.; Lucretti, S.; Palombieri, S.; Masci, S.; Lafiandra, D. A Cross between Bread Wheat and a 2D(2R) Disomic Substitution Triticale Line Leads to the Formation of a Novel Disomic Addition Line and Provides Information of the Role of Rye Secalins on Breadmaking Characteristics. Int. J. Mol. Sci. 2020, 21, 8450. [Google Scholar] [CrossRef]
- Dreccer, M.F.; Condon, A.G.; Macdonald, B.; Rebetzke, G.J.; Awasi, M.-A.; Borgognone, M.G.; Peake, A.; Piñera-Chavez, F.J.; Hundt, A.; Jackway, P.; et al. Genotypic variation for lodging tolerance in spring wheat: Wider and deeper root plates, a feature of low lodging, high yielding germplasmmm. Field Crop Res. 2020, 258, 107942. [Google Scholar] [CrossRef]
- Hura, T.; Dziurka, M.; Hura, K.; Ostrowska, A.; Dziurka, K.; Gadzinowska, J. Wheat and rye genome confer specific phytohormone profile features and interplay under water stress in two phenotypes of triticale. Plant Physiol. Biochem. 2017, 118, 494–509. [Google Scholar] [CrossRef]
- Trini, J.; Maurer, H.P.; Neuweiler, J.E.; Würschum, T. Identification and Fine-Mapping of Quantitative Trait Loci Controlling Plant Height in Central European Winter Triticale (×Triticosecale Wittmack). Plants 2021, 10, 1592. [Google Scholar] [CrossRef] [PubMed]
- Jatayev, S.; Sukhikh, I.; Vavilova, V.; Smolenskaya, S.E.; Goncharov, N.P.; Kurishbayev, A.; Zotova, L.; Absattarova, A.; Serikbay, D.; Hu, Y.; et al. Green revolution ‘stumbles’ in a dry environment: Dwarf wheat with Rht genes fails to produce higher grain yield than taller plants under drought. Plant Cell Environ. 2020, 43, 2355–2364. [Google Scholar] [CrossRef] [PubMed]
- Sukhikh, I.; Vavilova, V.; Blinov, A.; Goncharov, N. Diversity and Phenotypical Effect of the Allele Variants of Dwarfing Rht Genes in Wheat. Russ J. Genet. 2021, 57, 127–138. [Google Scholar] [CrossRef]
- Dowla, M.; Islam, S.; Stefanova, K.; Hara, G.O.; Ma, W.; Edwards, I. Phenology and Dwarfing Gene Interaction Effects on the Adaptation of Selected Wheat (Triticum aestivum L.) Advanced Lines across Diverse Water-Limited Environments of Western Australia. Agriculture 2020, 10, 470. [Google Scholar] [CrossRef]
- van der Knaap, E.; Kim, J.H.; Kende, H. A novel gibberellin-induced gene from rice and its potential regulatory role in stem growth. Plant Physiol. 2000, 122, 695–704. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Tian, Y.; Wu, K.; Ye, Y.; Yu, J.; Zhang, J.; Liu, Q.; Hu, M.; Li, H.; Tong, Y.; et al. Modulating plant growth–metabolism coordination for sustainable agriculture. Nature 2018, 560, 595–600. [Google Scholar] [CrossRef]
- Che, R.; Tong, H.; Shi, B.; Liu, Y.; Fang, S.; Liu, D.; Xiao, Y.; Hu, B.; Liu, L.; Wang, H.; et al. Control of grain size and rice yield by GL2-mediated brassinosteroid responses. Nat. Plants 2016, 2, 15195. [Google Scholar] [CrossRef]
- Zhang, J.; Zhou, Z.; Bai, J.; Tao, X.; Wang, L.; Zhang, H.; Zhu, J.K. Disruption of MIR396e and MIR396f improves rice yield under nitrogen-deficient conditions. Natl. Sci. Rev. 2020, 7, 102–112. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.; He, Y.; Yang, L.; Lu, C.; Zhu, Y.; Sun, C.; Ma, D.; Yin, J. Genome-wide analysis of growth-regulating factors (GRFs) in Triticum aestivum. PeerJ 2021, 9, e10701. [Google Scholar] [CrossRef] [PubMed]
- Kroupin, P.Y.; Chernook, A.; Bazhenov, M.; Karlov, G.I.; Goncharov, N.P.; Chikida, N.N.; Divashuk, M.G. Allele mining of TaGRF-2D gene 5′-UTR in Triticum aestivum and Aegilops tauschii genotypes. PLoS ONE 2020, 15, e0231704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, K.; Liu, Y.G. DELLA-GRF4-mediated coordination of growth and nitrogen metabolism paves the way for a new Green Revolution. Sci. China Life Sci. 2018, 61, 1130–1131. [Google Scholar] [CrossRef]
- Xue, H.; Zhang, Y.; Xiao, G. Neo-gibberellin signaling: Guiding the next generation of the green revolution. Trends Plant Sci. 2020, 25, 520–522. [Google Scholar] [CrossRef] [PubMed]
- Ben Amer, I.M.; Korzun, V.; Worland, A.J.; Börner, A. Genetic mapping of QTL controlling tissue culture response on chromosome 2B of wheat (Triticum aestivum L.) in relation to major genes and RFLP markers. Theor. Appl. Genet. 1997, 94, 1047–1052. [Google Scholar] [CrossRef]
- Börner, A.; Röder, M.; Korzun, V. Comparative molecular mapping of GA insensitive Rht loci on chromosomes 4B and 4D of wheat (Triticum aestivum L.). Theor. Appl. Genet. 1997, 95, 1133–1137. [Google Scholar] [CrossRef]
- Korzun, V.; Börner, A.; Worland, A.J.; Law, C.N.; Röder, M.S. Application of microsatellite markers to distinguish inter-varietal chromosome substitution lines of wheat (Triticum aestivum L.). Euphytica 1997, 95, 149–155. [Google Scholar] [CrossRef]
- Korzun, V.; Röder, M.; Worland, A.J.; Börner, A. Intrachromosomal mapping of the dwarfing (Rht12) and vernalisation response (Vrn1) genes 33 in wheat by using RFLP and microsatellite markers. Plant Breed. 1997, 116, 227–232. [Google Scholar] [CrossRef]
- Braun, E.M.; Tsvetkova, N.; Rotter, B.; Siekmann, D.; Schwefel, K.; Krezdorn, N.; Plieske, J.; Winter, P.; Melz, G.; Voylokov, A.; et al. Gene Expression Profiling and Fine Mapping Identifies a Gibberellin 2-Oxidase Gene Co-segregating With the Dominant Dwarfing Gene Ddw1 in Rye (Secale cereale L.). Front. Plant Sci. 2019, 10, 857. [Google Scholar] [CrossRef]
- Divashuk, M.G.; Litvinov, D.Y.; Chernook, A.G.; Nazarova, L.A.; Karlov, G.I.; Kroupin, P.Y.; Bazhenov, M.S. Effect of allelic forms of GRFs genes on the development of common wheat under different conditions of nitrogen supplementation. Plant Genet. Genom. Bioinform. Biotechnol. 2021, 10, 53. [Google Scholar] [CrossRef]
- Kroupin, P.; Chernook, A.; Karlov, G.; Soloviev, A.; Divashuk, M. Effect of Dwarfing Gene Ddw1 on Height and Agronomic Traits in Spring Triticale in Greenhouse and Field Experiments in a Non-Black Earth Region of Russia. Plants 2019, 8, 131. [Google Scholar] [CrossRef] [Green Version]
- Kroupin, P.; Chernook, A.; Karlov, G.I.; Soloviev, A.; Korshunova, A.D.; Divashuk, M.G. Effects of Dwarfing Wheat (Triticum aestivum L.) and rye (Secale cereale L.) genes in spring triticale segregating population asstudied in pot trials. Sel’skokhozyaistvennaya Biol. 2019, 54, 920–933. [Google Scholar]
- Kroupin, P.Y.; Karlov, G.I.; Bespalova, L.A.; Salina, E.A.; Chernook, A.G.; Watanabe, N.; Bazhenov, M.S.; Panchenko, V.V.; Nazarova, L.A.; Kovtunenko, V.Y.; et al. Effects of Rht17 in combination with Vrn-B1 and Ppd-D1 alleles on agronomic traits in wheat in black earth and non-black earth regions. BMC Plant Biol. 2020, 20, 304. [Google Scholar] [CrossRef] [PubMed]
- Chernook, A.; Kroupin, P.; Karlov, G.; Soloviev, A.; Korshunova, A.; Rubets, V.; Igonin, V.; Divashuk, M. Effects of Rht-B1b and Ddw1 Dwarfing Genes in Two Connecting Populations of Spring Triticale under Greenhouse Experiment Conditions. Agriculture 2019, 9, 119. [Google Scholar] [CrossRef] [Green Version]
- Tikhenko, N.; Tsvetkova, N.; Voylokov, A. The Effect of Parental Genotypes of Rye Lines on the Development of Quantitative Traits in Primary Octoploid Triticale: Plant Height. Russ. J. Genet. 2003, 39, 52–56. [Google Scholar] [CrossRef]
- Khlestkina, E.; Shvachko, N.; Zavarzin, A.; Börner, A. Vavilov’s Series of the “Green Revolution” Genes. Russian J. Genet. 2020, 56, 1371–1380. [Google Scholar] [CrossRef]
- Banaszak, Z. Breeding of Triticale in DANKO. 61 Tagung der Vereinigung der Pflanzenzüchter und Saatgutkaufleute Österreichs; Raumberg-Gumpenstein: Raumberg, Austria, 2010; pp. 65–68. [Google Scholar]
- Ittu, G.; Saulescu, N.; Ittu, M.; Mustatea, P. Present and perspectives in Romanian triticale breeding program. Commun Agric Appl. Biol. Sci. 2014, 79, 185–191. [Google Scholar]
- Braun, E.M. Erstellung Einer Hochauflösenden Karte für das Dominante Verzwergungsgen Ddw1 im Winterroggen (Secale cereale L.): D.—Technische Universität München. Ph.D. Thesis, Technische Universität München, Munich, Germany, 2021. [Google Scholar]
- Zan, T.; Zhang, L.; Xie, T.; Li, L. Genome-Wide Identification and Analysis of the Growth-Regulating Factor (GRF) Gene Family and GRF-Interacting Factor Family in Triticum aestivum L. Biochem Genet. 2020, 58, 705–724. [Google Scholar] [CrossRef]
- Sun, L.; Yang, W.; Li, Y.; Shan, Q.; Ye, X.; Wang, D.; Yu, K.; Lu, W.; Xin, P.; Pei, Z.; et al. A wheat dominant dwarfing line with Rht12, which reduces stem cell length and affects gibberellic acid synthesis, is a 5AL terminal deletion line. Plant J. 2019, 97, 887–900. [Google Scholar] [CrossRef] [PubMed]
- Buss, W.; Ford, B.A.; Foo, E.; Schnippenkoetter, W.; Borrill, P.; Brooks, B.; Ashton, A.R.; Chandler, P.M.; Spielmeyer, W. Overgrowth mutants determine the causal role of gibberellin GA2oxidaseA13 in Rht12 dwarfism of wheat. J. Exp. Bot. 2020, 71, 7171–7178. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Liu, H.; Guo, S.; Wang, B.; Li, Z.; Chong, K.; Xu, Y. OsmiR396d Affects Gibberellin and Brassinosteroid Signaling to Regulate Plant Architecture in Rice. Plant Physiol. 2018, 176, 946–959. [Google Scholar] [CrossRef] [Green Version]
- Kalih, R.; Maurer, H.; Miedaner, T. Genetic Architecture of Fusarium Head Blight Resistance in Four Winter Triticale Populations. Phytopathology 2015, 105, 334–341. [Google Scholar] [CrossRef] [Green Version]
- Wolski, T.; Gryka, J. Semi-dwarf winter triticale. In Triticale—Today and Tomorrow; Developments in Plant Breeding; Guedes-Pinto, H., Darvey, N., Carnide, V.P., Eds.; Kluwer: Dordrecht, The Netherlands, 1996; Volume 5, pp. 581–587. ISBN 978-94-010-6634-1. [Google Scholar]
- Chen, L.; Phillips, A.L.; Condon, A.G.; Parry, M.A.J.; Hu, Y.-G. GA-Responsive Dwarfing Gene Rht12 Affects the Developmental and Agronomic Traits in Common Bread Wheat. PLoS ONE 2013, 8, e62285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Hao, L.; Condon, A.G.; Hu, Y.-G. Exogenous GA3 Application Can Compensate the Morphogenetic Effects of the GA-Responsive Dwarfing Gene Rht12 in Bread Wheat. PLoS ONE 2014, 9, e86431. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Du, Y.; Lu, Q.; Chen, H.; Meng, R.; Cui, C.; Lu, S.; Yang, Y.; Chai, Y.; Li, J.; et al. The Photoperiod-Insensitive Allele Ppd-D1a Promotes Earlier Flowering in Rht12 Dwarf Plants of Bread Wheat. Front. Plant Sci. 2018, 9, 1312. [Google Scholar] [CrossRef] [PubMed]
- Worland, A.J.; Sayers, E.J.; Börner, A. The Genetics and Breeding Potential of Rht12, a Dominant Dwarfing Gene in Wheat. Plant Breed. 1994, 113, 187–196. [Google Scholar] [CrossRef]
- Komyshev, E.; Genaev, M.; Afonnikov, D. Evaluation of the SeedCounter, A Mobile Application for Grain Phenotyping. Front. Plant Sci. 2017, 7, 1990. [Google Scholar] [CrossRef] [Green Version]
- Litvinov, D.Y.; Chernook, A.G.; Kroupin, P.Y.; Bazhenov, M.S.; Karlov, G.I.; Avdeev, S.M.; Divashuk, M.G. A Convenient Co-Dominant Marker for Height-Reducing Ddw1 Allele Useful for Marker-Assisted Selection. Agriculture 2020, 10, 110. [Google Scholar] [CrossRef] [Green Version]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 3rd ed.; Sage: Thousand Oaks, CA, USA, 2019; Errata and updates (as of 2020-05-17); Available online: https://socialsciences.mcmaster.ca/jfox/Books/Companion/ (accessed on 17 May 2020).
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Month | Sum of Precipitation (mm) | Average Temperature (°C) | ||
|---|---|---|---|---|
| 2018 | 2019 | 2018 | 2019 | |
| May | 44 | 58 | 16.1 | 16.2 |
| June | 54 | 55 | 17.2 | 19.6 |
| July | 85 | 64 | 20.3 | 16.7 |
| August | 20 | 48 | 19.8 | 16.4 |
| Genotype | F4:5(2018) | F5:6(2019) |
|---|---|---|
| Ddw1Ddw1 Grf3(274)Grf3(274) | 45 | 268 |
| ddw1 ddw1 Grf3(274)Grf3(274) | 216 | 304 |
| ddw1 ddw1 Grf3(262)Grf3(262) | 351 | 472 |
| Ddw1Ddw1 Grf3(262)Grf3(262) | 106 | 262 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Divashuk, M.; Chernook, A.; Kroupina, A.; Vukovic, M.; Karlov, G.; Ermolaev, A.; Shirnin, S.; Avdeev, S.; Igonin, V.; Pylnev, V.; et al. TaGRF3-2A Improves Some Agronomically Valuable Traits in Semi-Dwarf Spring Triticale. Plants 2021, 10, 2012. https://doi.org/10.3390/plants10102012
Divashuk M, Chernook A, Kroupina A, Vukovic M, Karlov G, Ermolaev A, Shirnin S, Avdeev S, Igonin V, Pylnev V, et al. TaGRF3-2A Improves Some Agronomically Valuable Traits in Semi-Dwarf Spring Triticale. Plants. 2021; 10(10):2012. https://doi.org/10.3390/plants10102012
Chicago/Turabian StyleDivashuk, Mikhail, Anastasiya Chernook, Aleksandra Kroupina, Milena Vukovic, Gennady Karlov, Aleksey Ermolaev, Sergey Shirnin, Sergey Avdeev, Vladimir Igonin, Vladimir Pylnev, and et al. 2021. "TaGRF3-2A Improves Some Agronomically Valuable Traits in Semi-Dwarf Spring Triticale" Plants 10, no. 10: 2012. https://doi.org/10.3390/plants10102012