
Studies on Reproductive Development and Breeding Habit of the Commercially Important Bamboo Bambusa tulda Roxb


 and
and
Abstract
:1. Introduction
2. Results
2.1. Observations on Recurrent, Sporadic Flowering Cycle of B. tulda for Seven Years
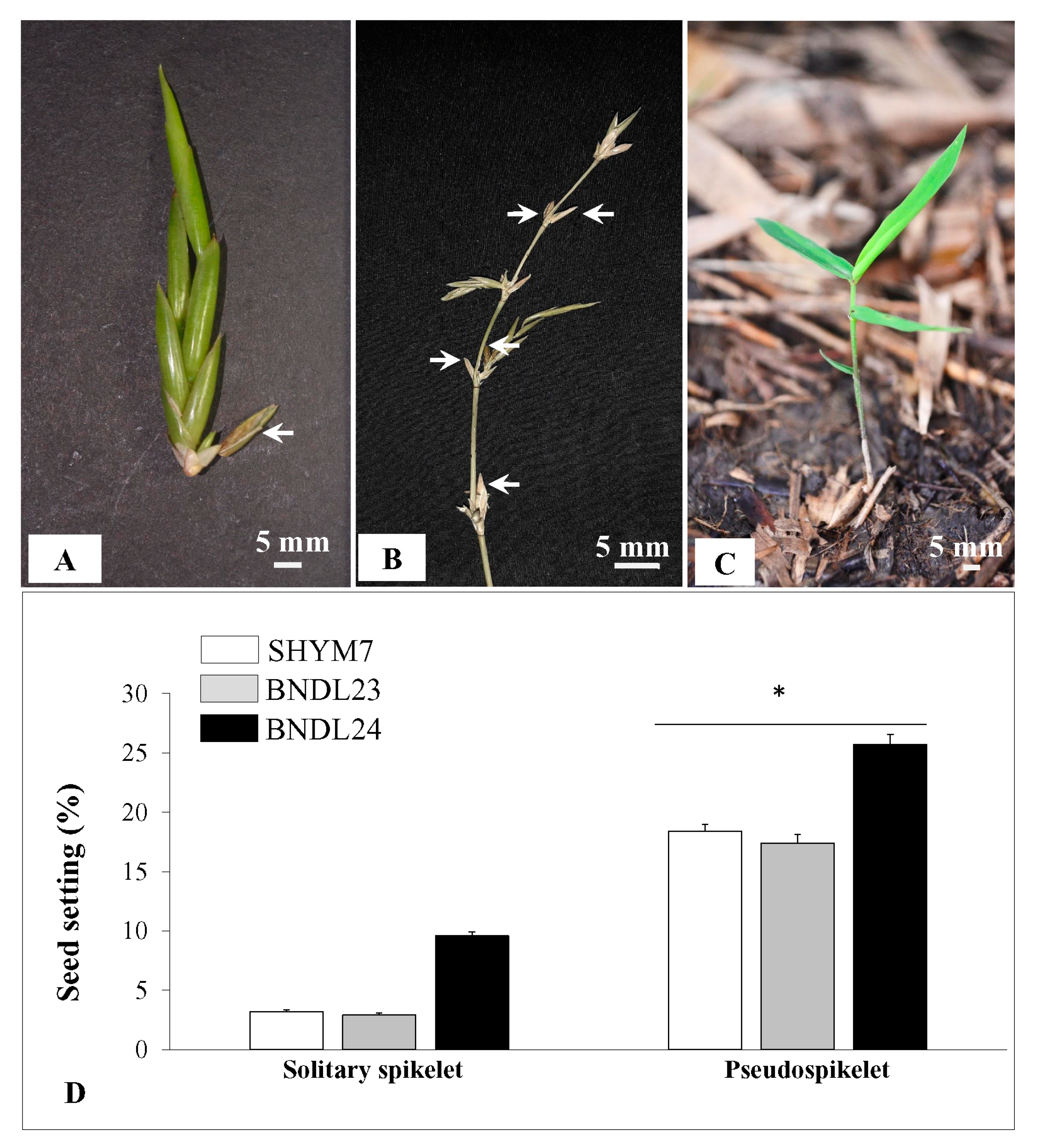
2.2. Macro- and Micro-Morphology of Solitary Spikelet and Pseudospikelet
2.3. Morphology of Florets and Micro Morphology of Floral Bracts
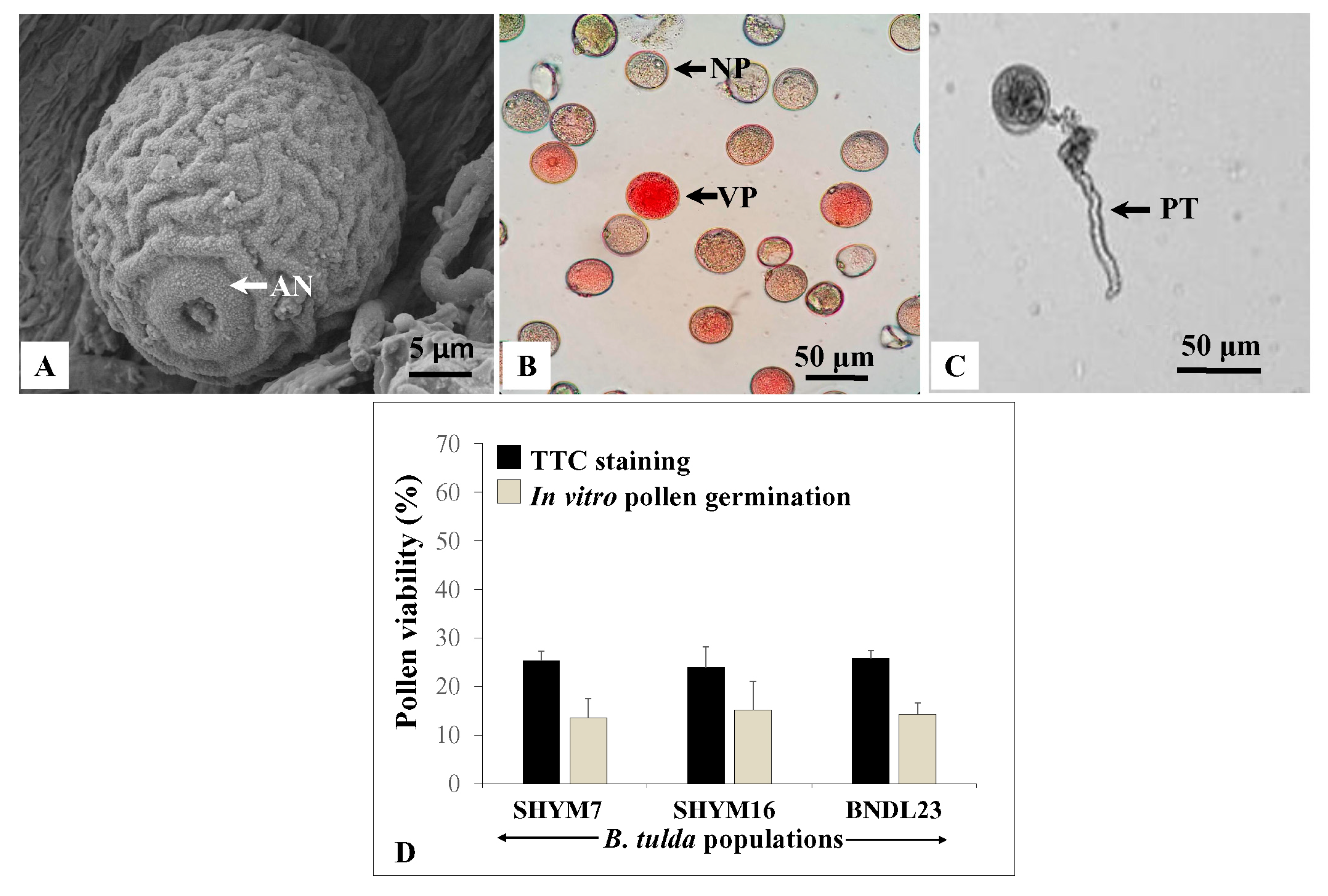
2.4. Morphology, Cytology and Germination of Pollen Grains
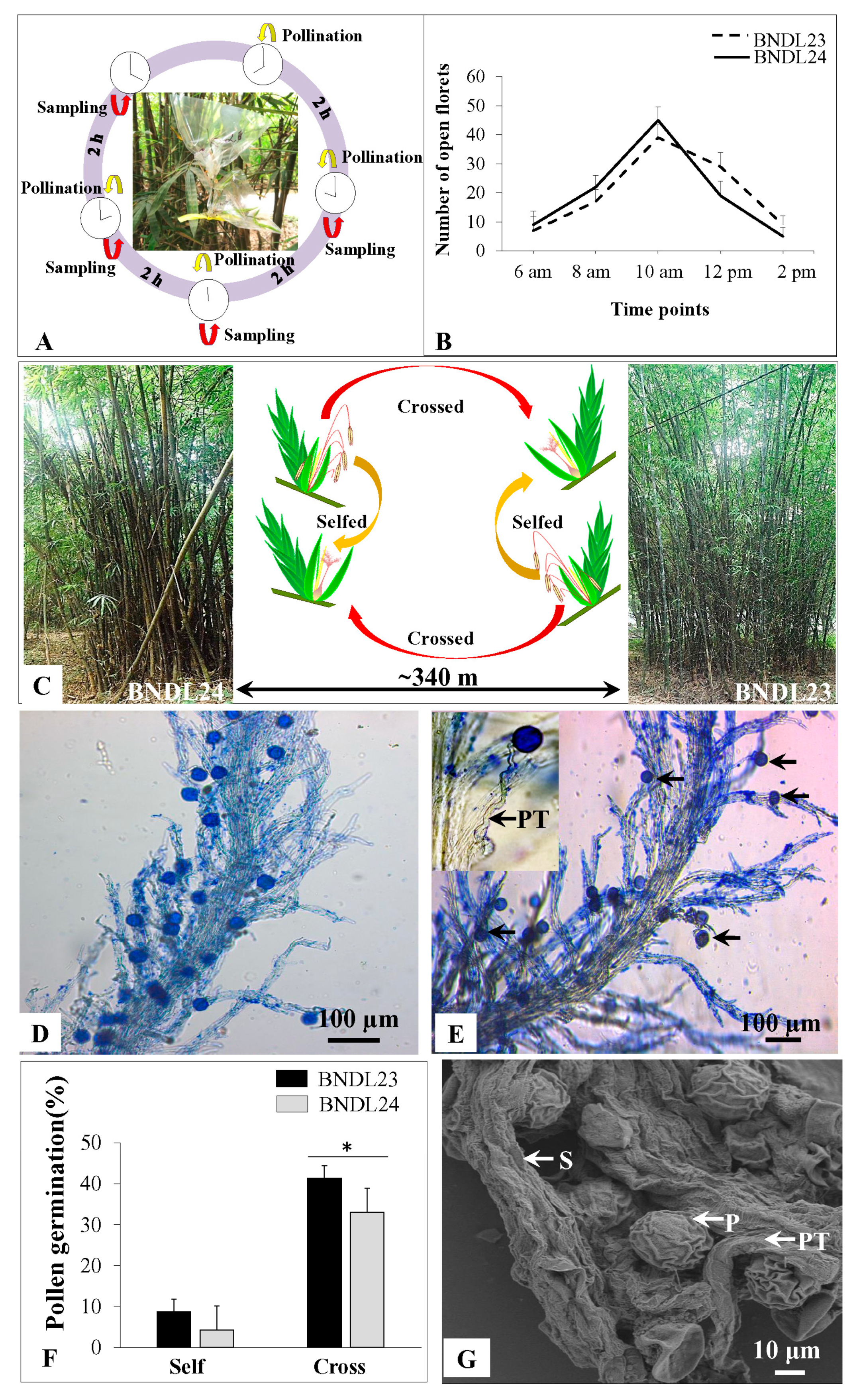
2.5. Self-Incompatibility in B. tulda
2.6. Higher Rate of Seed Setting in Pseudospikelet Compared to Solitary Spikelet Inflorescences
3. Discussion
3.1. Flowering Time, Cycle and Inflorescence Types: Why So Much Diversity?
3.2. Fragmented Flowering Population, Low Pollen Viability and Dispersal Lead to Reduced Seed Setting
3.3. Protandry and High Rate of Genetic Incompatibility to Ensure Genetic Variability
3.4. Semelparous Gregarious Flowering vs. Iteroparous Sporadic Flowering: Ecological Benefits and Costs
4. Materials and Methods
4.1. Population of B. tulda Studied
4.2. Studies on Inflorescence and Floral Morphology and Pollen Cytology of B. tulda
4.3. Scanning Electron Microscopy (SEM) Analyses of Inflorescence Buds, Floral Bracts and Pollen Grains
4.4. Test of Pollen Viability by Tetrazolium Staining and In Vitro Pollen Germination Assay
4.5. Artificial Pollination Conducted in Situ
4.6. Seed Numbers Obtained from Solitary Spikelet vs. Pseudospikelet
4.7. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Soreng, R.; Peterson, P.; Romaschenko, K.; Davidse, G.; Teisher, J.; Clark, L.; Barbera, P.; Gillespie, L.; Zuloaga, F. A worldwide phylogenetic classification of the Poaceae (Gramineae) II: An update and a comparison of two 2015 classifications. J. Syst. Evol. 2017, 55, 259–290. [Google Scholar] [CrossRef] [Green Version]
- Das, M.; Bhattacharya, S.; Singh, P.; Filgueiras, T.S.; Pal, A. Bamboo taxonomy and diversity in the era of molecular markers. Adv. Bot. Res. 2008, 47, 225–268. [Google Scholar]
- Bystriakova, N.; Kapos, V.; Lysenko, I.; Stapleton, C. Distribution and conservation status of forest bamboo biodiversity in the Asia-Pacific region. Biodivers. Conserv. 2003, 12, 1833–1841. [Google Scholar] [CrossRef]
- Liu, W.; Hui, C.; Wang, F.; Wang, M.; Liu, G. Review of the Resources and Utilization of Bamboo in China; IntechOpen: London, UK, 2018; pp. 1–10. [Google Scholar] [CrossRef] [Green Version]
- Patel, A.B. Traditional bamboo uses by the tribes of Gujarat. Indian J. Tradit. Knowl. 2005, 4, 179–184. [Google Scholar]
- Ju, Y.; Zhuo, J.; Liu, B.; Long, C. Eating from the wild: Diversity of wild edible plants used by Tibetans in Shangri-la region, Yunnan, China. J. Ethnobiol. 2013, 9, 28. [Google Scholar] [CrossRef] [Green Version]
- Dhyani, S.; Dhyani, D. Significance of provisioning ecosystem services from moist temperate forest ecosystems: Lessons from upper Kedarnath valley, Garhwal, India. Energ. Ecol. Environ. 2016, 1, 109–121. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, S.; Basak, M.; Dutta, S.; Chanda, C.; Dey, A.; Das, M. Ethnobamboology: Traditional uses of bamboos and opportunities to exploit genomic resources for better exploitation. In Biotechnological Advances in Bamboo: The “Green Gold” on the Earth; Ahmad, Z., Ding, Y., Shahzad, A., Eds.; Springer Nature: Singapore, 2021. [Google Scholar]
- Lin, S.Y.; Mao, G.X. The habit and regeneration of bamboo flowering. Forest Sci. Technol. 2007, 32, 23–25. [Google Scholar]
- Zheng, X.; Lin, S.; Fu, H.; Wan, Y.; Ding, Y. The bamboo flowering cycle sheds light on flowering diversity. Front. Plant Sci. 2020, 11, 381. [Google Scholar] [CrossRef] [PubMed]
- Jeeva, S.; Kiruba, S. Flowering of bamboos in two biodiversity hotspots of India. Indian For. 2010, 136, 137–140. [Google Scholar]
- Naithani, H.B. Contribution to the Taxonomic Studies of Indian Bamboos. Ph.D. Thesis, Garhwal University, Srinagar, India, 1993. [Google Scholar]
- Bhattacharya, S.; Das, M.; Bar, R.; Pal, A. Morphological and molecular characterization of Bambusa tulda with a note on flowering. Ann. Bot. 2006, 98, 529–535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thapliyal, M. Flowering in Dendrocalamus Asper; FRI: Dehradun, Indian, 2011; Unpublished. [Google Scholar]
- Sharma, H.R.; Yadav, S.; Deka, B.; Meena, R.K.; Bisht, N.S. Sporadic flowering of Dendrocalamus longispathus (Kurz) Kurz in Mizoram, India. Tropic. Plant Res. 2014, 1, 26–27. [Google Scholar]
- Bahadur, K.N. A note on the flowering of Bambusa nutans. Indian For. 1980, 106, 314–316. [Google Scholar]
- Troup, R.S. The Silviculture of Indian Trees; Clarendon Press: Oxford, UK, 1921. [Google Scholar]
- Perry, N.E. On the flowering of bamboos. J. Bombay Nat. Hist. Soc. 1931, 34, 1099–1101. [Google Scholar]
- Mohan Ram, H.Y.; Gopal, B.H. Some observation on the flowering of bamboos in Mizoram. Curr. Sci. 1981, 50, 708–710. [Google Scholar]
- Rawat, B.S. Flowering of bamboos. Indian For. 1987, 113, 760–761. [Google Scholar]
- Sarma, H.; Sarma, A.M.; Sarma, A.; Borah, S. A case of gregarious flowering in bamboo, dominated lowland forest of Assam, India: Phenology, regeneration, impact on rural economy, and conservation. J. Forest. Res. 2010, 21, 409. [Google Scholar] [CrossRef]
- Naithani, H.B.; Deka, A.K.; Banerjee, G. Gregarious flowering of bamboos in Arunachal Pradesh. Indian For. 2013, 139, 746. [Google Scholar]
- Bhattacharya, S.; Ghosh, J.S.; Das, M.; Pal, A. Morphological and molecular characterization of Thamnocalamus spathiflorus subsp. spathiflorus at population level. Plant Syst. Evol. 2009, 282, 13–20. [Google Scholar] [CrossRef]
- Verma, P.K.; Rawat, K.K.; Das, N.; Yadav, A.; Kaushik, P.K. A note on recent flowering in Thamnocalamus aristatus (Gamble) E.G Camus (Poaceae: Bambusoideae) in East Sikkim. Research 2013, 5, 19–21. [Google Scholar]
- Janzen, D.H. Why bamboos wait so long to flower. Ann. Rev. Ecol. Syst. 1976, 7, 347–391. [Google Scholar] [CrossRef]
- Wang, W.; Franklin, S.B.; Lu, Z.; Rude, B.J. Delayed flowering in bamboo: Evidence from Fargesia qinlingensis in the Qinling Mountains of China. Front. Plant Sci. 2016, 7, 151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basak, M.; Dutta, S.; Biswas, S.; Chakraborty, S.; Sarkar, A.; Rahaman, T.; Dey, S.; Biswas, P.; Das, M. Genomic insights into growth and development of bamboos: What have we learnt and what more to discover? Trees 2021, in press. [Google Scholar] [CrossRef]
- Biswas, P.; Chakraborty, S.; Dutta, S.; Pal, A.; Das, M. Bamboo flowering from the perspective of comparative genomics and transcriptomics. Front. Plant Sci. 2016, 7, 1900. [Google Scholar] [CrossRef] [Green Version]
- Dutta, S.; Biswas, P.; Chakraborty, S.; Mitra, D.; Pal, A.; Das, M. Identification, characterization and gene expression analyses of important flowering genes related to photoperiodic pathway in bamboo. BMC Genom. 2018, 19, 190. [Google Scholar] [CrossRef] [Green Version]
- Dutta, S.; Deb, A.; Biswas, P.; Chakraborty, S.; Guha, S.; Mitra, D.; Geist, B.; Schäffner, A.R.; Das, M. Identification and functional characterization of two bamboo FD gene homologs having contrasting effects on shoot growth and flowering. Sci. Rep. 2021, 11, 7849. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.H.; Ma, P.F.; Yang, G.Q.; Hu, J.Y.; Liu, Y.L.; Xia, E.H.; Zhong, M.C.; Zhao, L.; Sun, G.L.; Xu, Y.X.; et al. Genome sequences provide insights into the reticulate origin and unique traits of woody bamboos. Mol. Plant 2019, 12, 1353–1365. [Google Scholar] [CrossRef]
- Mizuki, I.; Sato, A.; Matsuo, A.; Suyama, Y.; Suzuki, J.I.; Makita, A. Clonal structure, seed set, and self-oollination rate in mass-Flowering bamboo species during off-year flowering events. PLoS ONE 2014, 9, e105051. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.N.; Cui, Y.Z.; Wong, K.M.; Li, D.Z.; Yang, H.Q. Breeding system and pollination of two closely related bamboo species. AoB Plants 2017, 9, plx021. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Sanchez, E.; Peredo, L.C.; Santacruz, J.B.; Ayala-Barajas, R. Bamboo flowers visited by insects: Do insects play a role in the pollination of bamboo flowers? Plant Syst. Evol. 2017, 303, 51–59. [Google Scholar] [CrossRef]
- Xie, N.; Chen, L.N.; Dong, Y.R.; Yang, H.Q. Mixed mating system and variable mating patterns in tropical woody bamboos. BMC Plant Biol. 2019, 19, 418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, F.; Xue, J.R.; Yang, Y.M.; Hui, C.M.; Wang, J. Study on flowering phenomenon and its type of bamboo in Yunnan in past fifteen years. Sci. Silvae Sin. 2000, 36, 57–68. [Google Scholar]
- Xie, N.; Chen, L.N.; Wong, K.M.; Cui, Y.Z.; Yang, H.Q. Seed set and natural regeneration of Dendrocalamus membranaceus Munro after mass and sporadic flowering in Yunnan, China. PLoS ONE 2016, 11, e0153845. [Google Scholar] [CrossRef]
- Dhar, A. When Flowering Spells Famine. The Hindu. 2003. Available online: http://www.hinduonnet.com/2003/10/26/stories/2003102604461000.htm (accessed on 26 October 2003).
- Nath, A.J.; Das, A.K. Gregarious flowering of a long-livedtropical semelparous bamboo Schizostachyum dullooa in Assam. Curr. Sci. 2010, 99, 154–155. [Google Scholar]
- Abe, Y.; Shibata, S. Spatial and temporal flowering patterns of the monocarpic dwarf bamboo Sasa veitchii var. hirsuta. Ecol. Res. 2012, 27, 625–632. [Google Scholar] [CrossRef]
- Soderstrom, T.R. Some evolutionary trends in the bambusoideae (poaceae). Ann. Missouri Bot. Gar. 1981, 68, 15–47. [Google Scholar] [CrossRef]
- Wysocki, W.P.; Ruiz-Sanchez, E.; Yin, Y.; Duvall, M.R. The floral transcriptomes of four bamboo species (Bambusoideae; Poaceae): Support for common ancestry among woody bamboos. BMC Genom. 2016, 17, 384. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.Y.; Li, D.Z. Dendrocalamus menghanensis (Poaceae, Bambusoideae), a new woody bamboo from Yunnan, China. In: Cai J, Yu WB, Zhang T, Li DZ, eds. Revealing of the plant diversity in China’s biodiversity hotspots. PhytoKeys 2019, 130, 143–150. [Google Scholar] [CrossRef]
- Koshy, K.C.; Jee, G. Studies on the Absence of Seed Set in Bambusa Vulgaris. Curr. Sci. 2001, 81, 375–378. [Google Scholar]
- Pacini, E.; Dolferus, R. Pollen Developmental Arrest: Maintaining pollen fertility in a world with a changing climate. Front. Plant Sci. 2019, 10, 679. [Google Scholar] [CrossRef] [PubMed]
- De Souza, P.F.; Ribas Dos Santos, C.M.; Ree, J.; Guerra, M.P.; Pescador, R. Flowering and morphological characterization of Dendrocalamus asper androecium and pollen grains. Grana 2020, 60, 20–34. [Google Scholar] [CrossRef]
- Tushabe, D.; Rosbakh, S. A compendium of in vitro germination media for pollen research. Front. Plant Sci. 2021, 12, 709945. [Google Scholar] [CrossRef] [PubMed]
- Ramanayake, S.M.S.D.; Weerawardene, T.E. Flowering in a bamboo, Melocanna baccifera (Bambusoideae: Poaceae). Bot. J. Linn. Soc. 2003, 143, 287–291. [Google Scholar] [CrossRef]
- Bannert, M.; Stamp, P. Cross-pollination of maize at long distance. Eur. J. Agron. 2007, 27, 44–51. [Google Scholar] [CrossRef]
- Gu, Z.J.; Yang, H.Q.; Sun, M.S.; Yang, Y.M. Distribution characteristics, flowering and seeding of Dendrocalamus sinicus in Yunnan, China. Forest Res. 2012, 25, 1–5. [Google Scholar]
- Gagnon, P.R.; Platt, W.J. Reproductive and seedling ecology of a semelparous native bamboo (Arundinaria gigantea, Poaceae). J. Torrey Bot. Soc. 2008, 135, 309–316. [Google Scholar] [CrossRef]
- Venkatesh, C.S. Dichogamy and breeding system in a tropical bamboo Ochlandra travancorica. Biotropica 1984, 16, 309–312. [Google Scholar] [CrossRef]
- Cheung, Y.Y.; Copper, S.G.; Hansken, T.J.; Cheung, Y.C. Research on the raising of Phyllostachys pubescens seedling. In Recent Research on Bamboos, Proceedings of the International Workshop Recent Research on Bamboos, Hangzhou, China 06-14 October; IDRC: Ottawa, ON, Canada, 1985. [Google Scholar]
- Nadgauda, R.S.; John, C.K.; Mascarenhas, A.F. Floral biology and breeding behavior in the bamboo Dendrocalamus strictus Nees. Tree Physiol. 1993, 13, 401–408. [Google Scholar] [CrossRef]
- Huang, S.Q.; Yang, C.F.; Lu, B.; Takahashi, Y. Honeybee-assisted wind pollination in bamboo Phyllostachys nidularia (Bambusoideae: Poaceae)? Bot. J. Linn. Soc. 2002, 138, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Romero-Morte, J.; Rojo, J.; Rivero, R.; Fernández-González, F.; Pérez-Badia, R. Standardised index for measuring atmospheric grass-pollen emission. Sci. Total Environ. 2018, 612, 180–191. [Google Scholar] [CrossRef]
- García-Mozo, H. Poaceae pollen as the leading aeroallergen worldwide: A review. Allergy 2018, 72, 1849–1858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, K.M. Flowering fruiting and germination of the bamboo Schizostachyum zollingeri in Perlis. Malays. For. 1982, 44, 453–463. [Google Scholar]
- Das, M.C.; Gopakumar, B.; Shameena, M.K.; Jero-Mathu, A.; Nath, A.J.; Das, A.K. floral biology and pollen sterility in relation to seed set in bambusa balcooa. J. Trop. For. Sci. 2017, 29, 504–508. [Google Scholar]
- Jijeesh, C.M.; Seethalakshmi, K.K.; Raveendran, V.P. Flowering, reproductive biology and post flowering behaviour of Dendrocalamus sikkimensis Gamble, in Kerala, India. J. Am. Bamboo Soc. 2012, 25, 36–42. [Google Scholar]
- Kitamura, K.; Kawahara, T. Estimation of outcrossing rates at small-scale flowering sites of the dwarf bamboo species, Sasa cernua. J. Plant Res. 2011, 124, 683–688. [Google Scholar] [CrossRef]
- Liu, Z.; Lin, Y.; Zhou, M.; Tang, D. Estimation of outcrossing rate in Phyllostachys edulis by microsatellite polymorphism. Sci. Silvae Sin. 2014, 50, 76–81. [Google Scholar]
- Matsuo, A.; Tomimatsu, H.; Suzuki, J.I.; Saitoh, T.; Shibata, S.; Makita, A.; Suyama, Y. Female and male fitness consequences of clonal growth in a dwarf bamboo population with a high degree of clonal intermingling. Ann. Bot. 2014, 114, 1035–1041. [Google Scholar] [CrossRef] [Green Version]
- Zhong, Y.B.; Yue, J.J.; Lou, C.; Yuan, J.L.; Gu, X.P. Floral organ and breeding system of Dendrocalamus latiflorus. Sci. Silvae Sin. 2017, 53, 1–10. [Google Scholar]
- Langridge, P.; Baumann, U. Self-Incompatibility in the Grasses. In Self-Incompatibility in Flowering Plants; Springer: Berlin/Heidelberg, Germany, 2008. [Google Scholar]
- Silva, N.F.; Goring, D.R. Mechanisms of self-incompatibility in flowering plants. Cell Mol. Life Sci. 2001, 58, 1988–2007. [Google Scholar] [CrossRef]
- Ferrer, M.M.; Good-Avila, S.V.; Montaña, C.; Domínguez, C.A.; Eguiarte, L.E. Effect of variation in self-incompatibility on pollen limitation and inbreeding depression in Flourensia cernua (Asteraceae) scrubs of contrasting density. Ann. Bot. 2009, 103, 1077–1089. [Google Scholar] [CrossRef] [Green Version]
- Rai, V.; Dey, N. Identification of programmed cell death related genes in bamboo. Gene 2012, 497, 243–248. [Google Scholar] [CrossRef]
- Marchesini, V.A.; Sala, O.E.; Austin, A.T. Ecological consequences of a massive flowering event of bamboo (Chusquea culeou) in a temperate forest of Patagonia, Argentina. J. Veg. Sci. 2009, 20, 424–432. [Google Scholar] [CrossRef]
- Sertse, D.; Disasa, T.; Bekele, K.; Alebachew, M.; Kebede, Y.; Eshete, N. Mass flowering and death of bamboo: A potential threat to biodiversity and livelihoods in Ethiopia. J. Bio. Environ. Sci. 2011, 1, 16–25. [Google Scholar]
- Austin, A.T.; Marchesini, V.A. Gregarious flowering and death of understorey bamboo slow litter decomposition and nitrogen turnover in a southern temperate forest in Patagonia, Argentina. Funct. Ecol. 2012, 26, 265–273. [Google Scholar] [CrossRef]
- Taylor, A.H.; Jinyan, H.; Shi-Qiang, Z. Canopy tree development and undergrowth bamboo dynamics in old-growth Abies-Betula forests in southwestern China: A 12 year study. For. Ecol. Manag. 2004, 200, 347–360. [Google Scholar] [CrossRef]
- Holz, C.A.; Veblen, T.T. Tree regeneration response to Chusquea montana bamboo die-off in a subalpine nothofagus forest in the southern Andes. J. Veg. Sci. 2006, 17, 19–28. [Google Scholar] [CrossRef]
- Takahashi, M.; Furusawa, H.; Limtong, P.; Sunanthapongsuk, V.; Marod, D.; Panuthai, S. Soil nutrient status after bamboo flowering and death in a seasonal tropical forest in western Thailand. Ecol. Res. 2007, 22, 160–164. [Google Scholar] [CrossRef]
- Giordano, C.V.; Sánchez, R.A.; Austin, A.T. Gregarious bamboo flowering opens a window of opportunity for regeneration in a temperate forest of Patagonia. New Phytol. 2009, 181, 880–889. [Google Scholar] [CrossRef] [PubMed]
- Abe, Y.; Shibata, S. Flower and seed production in a series of flowerings from sporadic events before to after mass flowering of the dwarf bamboo Sasa veitchii var. hirsuta. J. For. Res. 2014, 19, 268–275. [Google Scholar] [CrossRef]
- Kitzberger, T.; Chaneton, E.J.; Caccia, F. Indirect effects of prey swamping: Differential seed predation during a bamboo masting event. Ecology 2007, 88, 2541–2554. [Google Scholar] [CrossRef] [PubMed]
- McClure, F.A. The Bamboos—A Fresh Perspective; Harvard University Press: Cambridge, UK, 1966; p. 347. [Google Scholar]
- Dafni, A.; Firmage, D. Pollen viability and longevity: Practical, ecological and evolutionary implications. Plant Syst. Evol. 2000, 222, 113–132. [Google Scholar] [CrossRef]
- Brewbaker, J.L.; Kwack, B.H. The essential role of calcium ions in pollen germination and pollen tube growth. Am. J. Bot. 1963, 50, 859–865. [Google Scholar] [CrossRef]
- Tuinstra, M.R.; Wedel, J. Estimation of pollen viability in sorghum. Crop Sci. 2000, 40, 968–970. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Population Number | Duration of Flowering | Clump and Culm | Year 1 | Year 2 | Year 3 | Year 4 | Year 5 | Year 6 | Year 7 |
|---|---|---|---|---|---|---|---|---|---|
| SHYM7 | 2013–16 | Number of flowering clumps | 1 | 1 | 2 | 1 | 0 | -- | -- |
| Total culm number in a flowering clump | 42 | 45 | 59 | 33 | -- | -- | -- | ||
| Number of flowering culms | 3 | 5 | 8 | 1 | -- | -- | -- | ||
| Number of withered culms | 0 | 1 | 2 | 7 | -- | -- | -- | ||
| SHYM16 | 2016–19 | Number of flowering clumps | 2 | 2 | 2 | 1 | 0 | -- | -- |
| Total culm number in a flowering clump | 38 | 41 | 40 | 24 | -- | -- | -- | ||
| Number of flowering culms | 7 | 8 | 5 | 2 | -- | -- | -- | ||
| Number of withered culms | 0 | 0 | 5 | 9 | -- | -- | -- | ||
| BNDL23 | 2013–19 | Number of flowering clumps | 4 | 4 | 3 | 3 | 3 | 3 | 1 |
| Total culm number in a flowering clump | 77 | 93 | 86 | 87 | 79 | 55 | 43 | ||
| Number of flowering culms | 11 | 14 | 17 | 15 | 6 | 5 | 1 | ||
| Number of withered culms | 2 | 5 | 9 | 6 | 11 | 12 | 8 | ||
| BNDL24 | 2017–to date | Number of flowering clumps | 1 | 1 | 2 | 1 | 2 | -- | -- |
| Total culm number in a flowering clump | 31 | 33 | 71 | 29 | 84 | -- | -- | ||
| Number of flowering culms | 6 | 10 | 9 | 8 | 11 | -- | -- | ||
| Number of withered culms | 1 | 0 | 5 | 2 | 1 | -- | -- |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chakraborty, S.; Biswas, P.; Dutta, S.; Basak, M.; Guha, S.; Chatterjee, U.; Das, M. Studies on Reproductive Development and Breeding Habit of the Commercially Important Bamboo Bambusa tulda Roxb. Plants 2021, 10, 2375. https://doi.org/10.3390/plants10112375
Chakraborty S, Biswas P, Dutta S, Basak M, Guha S, Chatterjee U, Das M. Studies on Reproductive Development and Breeding Habit of the Commercially Important Bamboo Bambusa tulda Roxb. Plants. 2021; 10(11):2375. https://doi.org/10.3390/plants10112375
Chicago/Turabian StyleChakraborty, Sukanya, Prasun Biswas, Smritikana Dutta, Mridushree Basak, Suman Guha, Uday Chatterjee, and Malay Das. 2021. "Studies on Reproductive Development and Breeding Habit of the Commercially Important Bamboo Bambusa tulda Roxb" Plants 10, no. 11: 2375. https://doi.org/10.3390/plants10112375