
A New MiRNA MiRm0002 in Eggplant Participates in the Regulation of Defense Responses to Verticillium Wilt


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
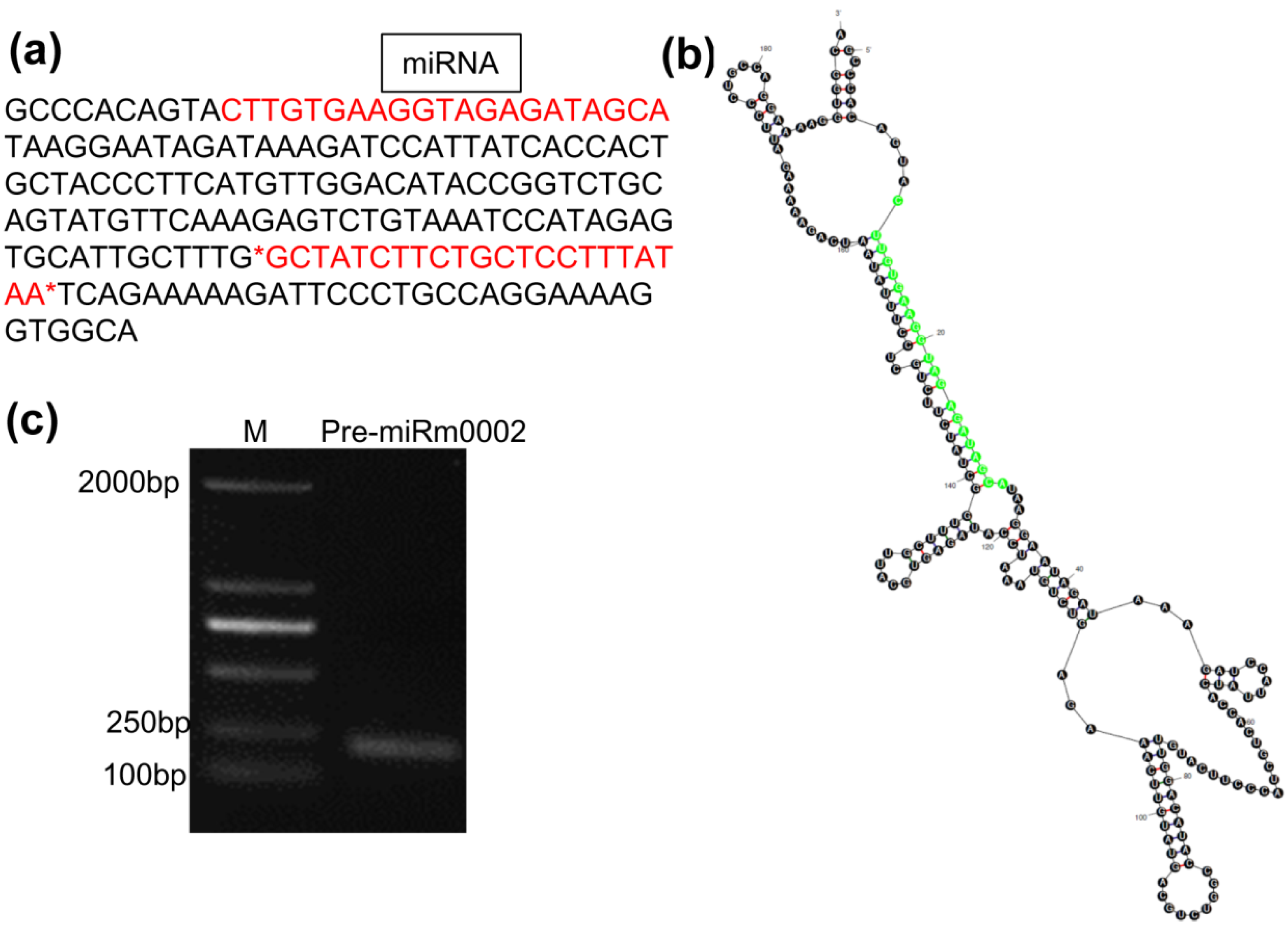
2.1. The Sequence, Stem Loop Structure, and Gene Clone of the MiRm0002 Precursor
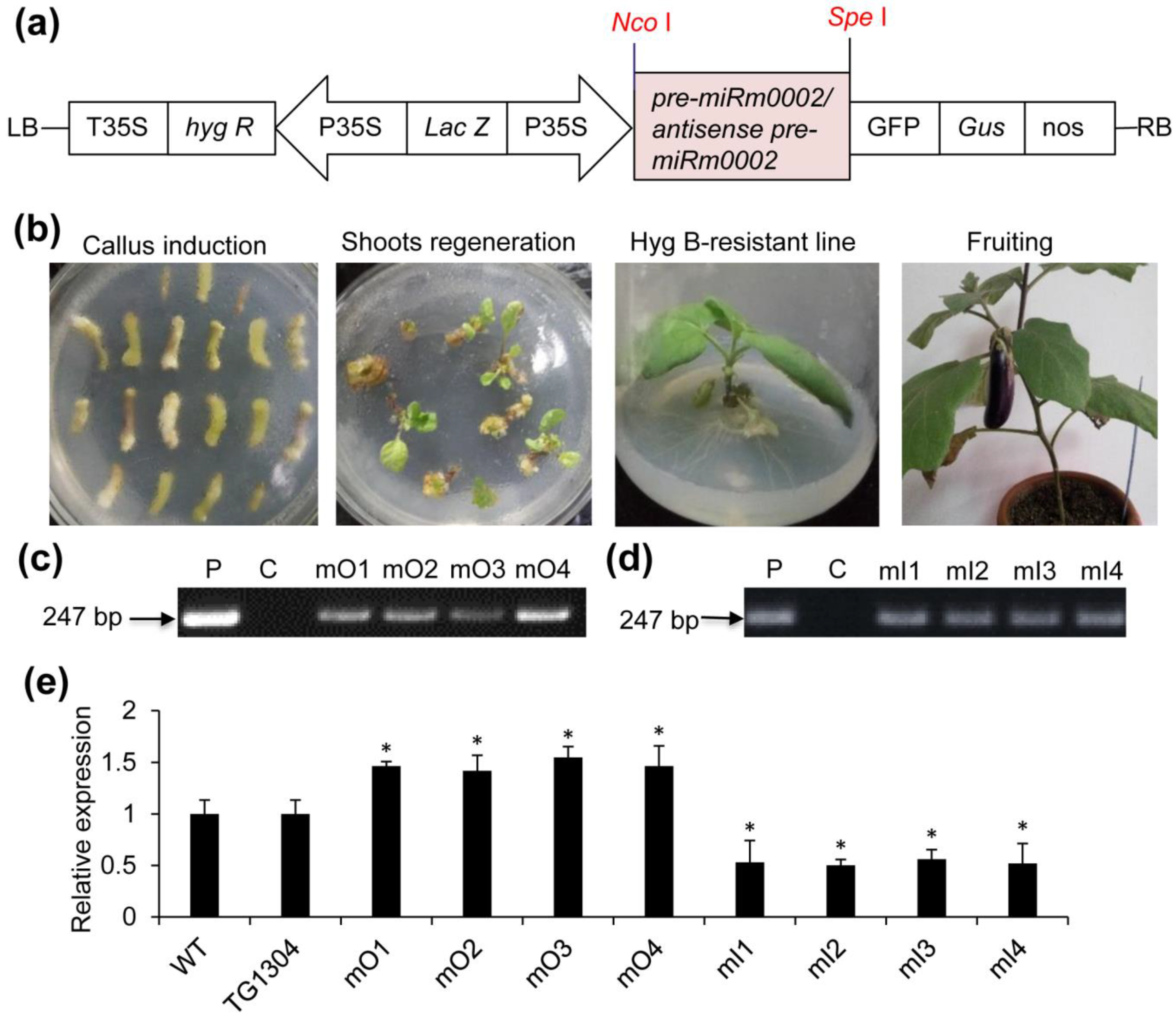
2.2. Production of Transgenic Eggplant
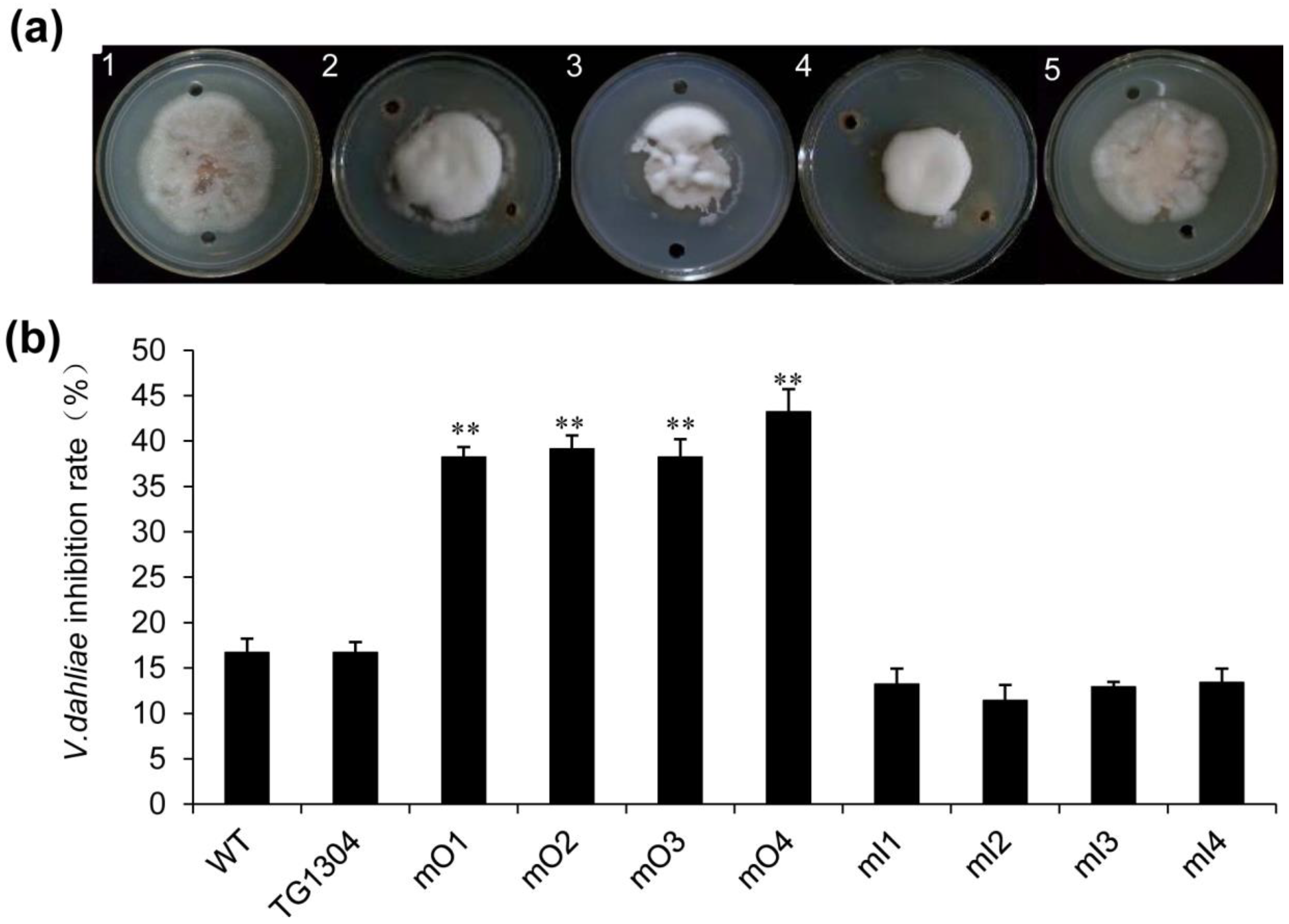
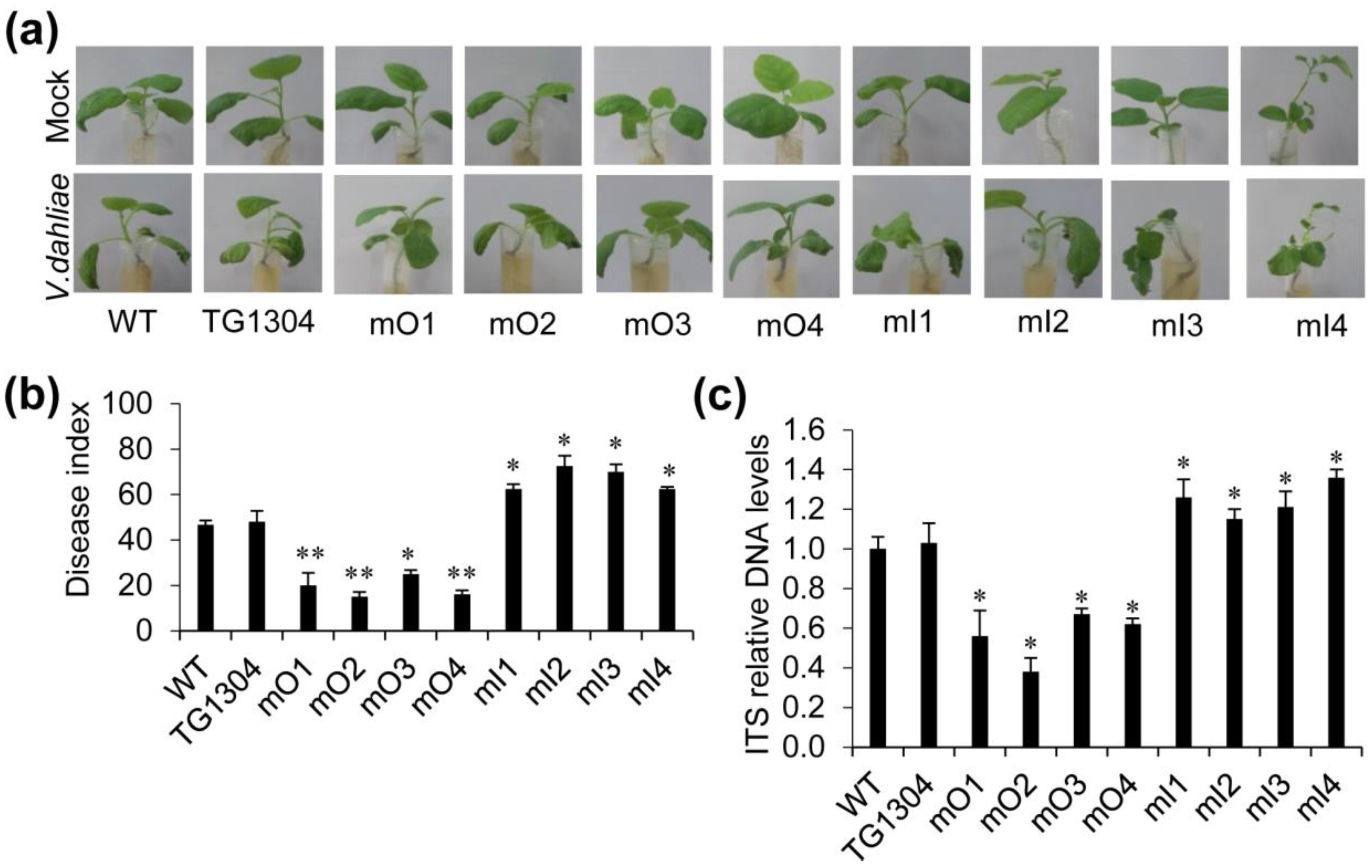
2.3. Disease Resistance of Transgenic Eggplant to V. Wilt
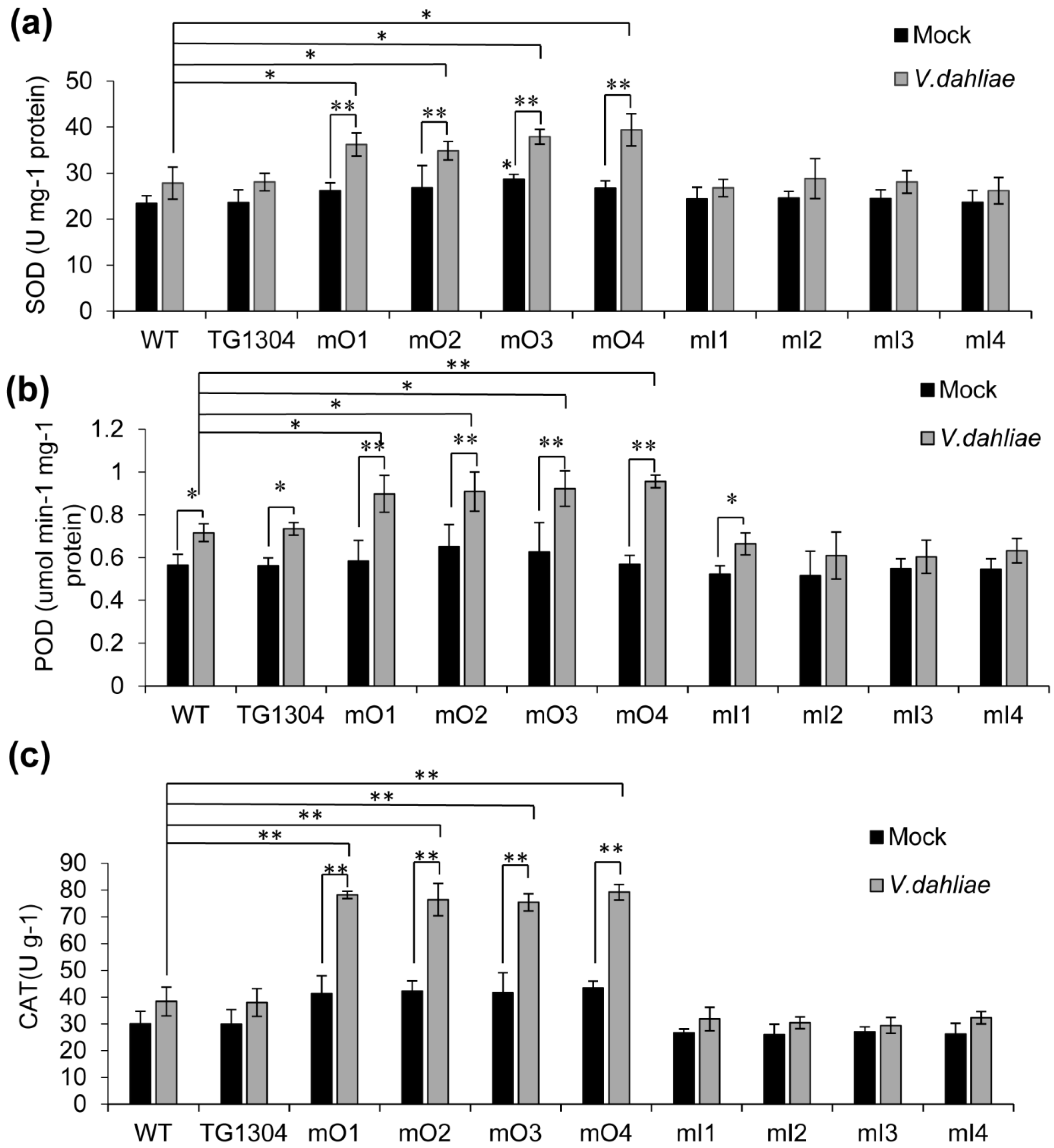
2.4. Antioxidant Activity in Transgenic Eggplants
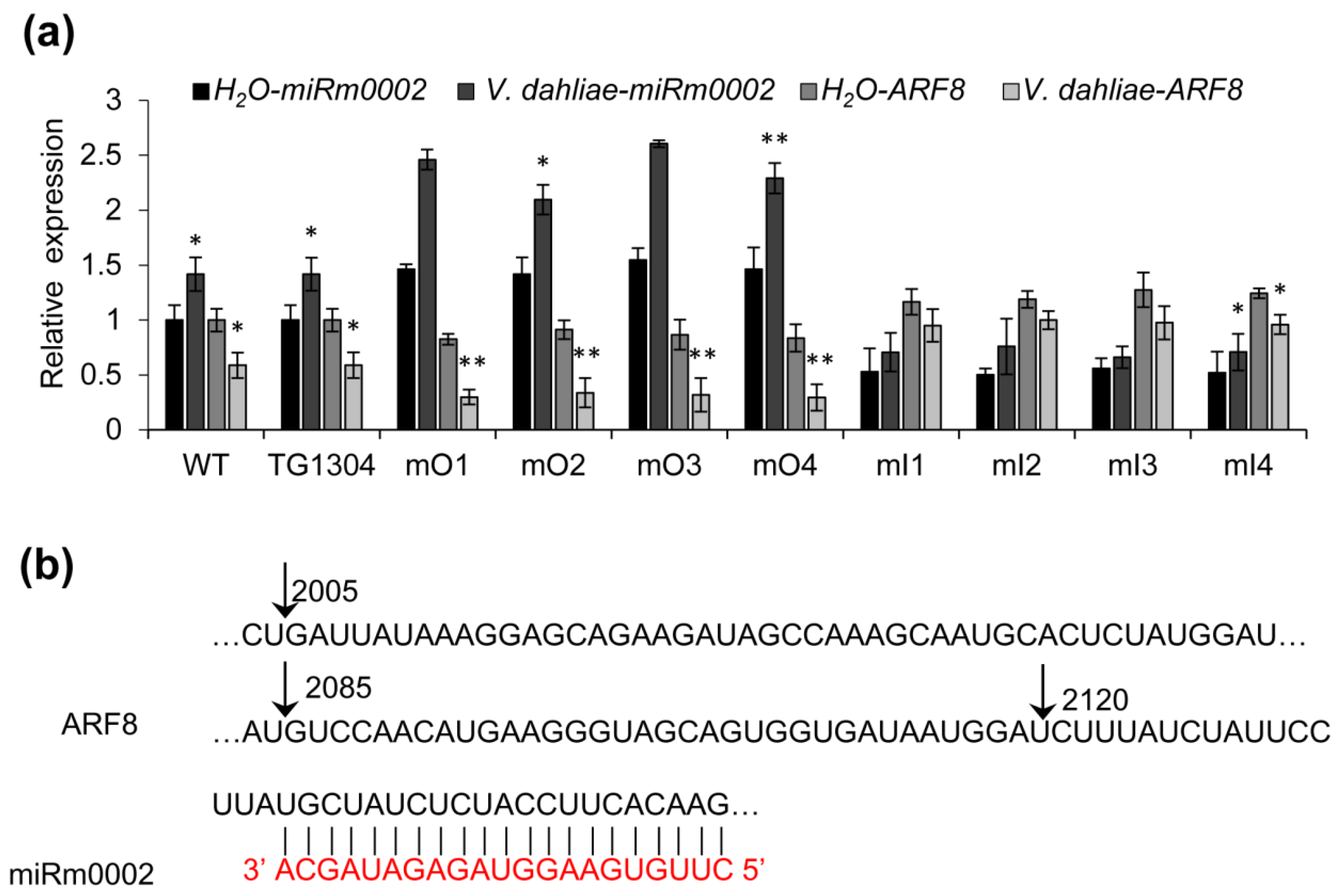
2.5. Analysis on Predicted Targets of MiRm0002
3. Discussion
4. Materials and Methods
4.1. Plants Material and Growth Conditions
4.2. Pathogen Culture and Infection
4.3. RNA Extraction and Expression Analysis
4.4. RACE Mapping of miRNA Target Cleavage Sites
4.5. Vector Construction and Plant Transformation
4.6. Transgenic Plant Identification
4.7. Evaluation of Plant Disease Resistance
4.8. Quantitative Analysis of V. dahliae in Tissues
4.9. In Vitro Anti-Fungal Assay
4.10. Antioxidant Enzyme Assay
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Marois, J.J.; Johnston, S.A.; Dunn, M.T.; Papavizas, G.C. Biological control of Verticillium Wilt of eggplant in the field. Plant. Dis. 1982, 66, 1166–1168. [Google Scholar] [CrossRef]
- Ciccarese, F.; Amenduni, M.; Schiavone, D.; Cirulli, M. Effect of Verticillium wilt on the yield of susceptible and "slow-wilting" resistant eggplants in the field. Phytopathol. Mediterr. 1994, 212–216. [Google Scholar]
- Liu, S.P.; Zhu, Y.P.; Xie, C.; Jue, D.W.; Hong, Y.B.; Chen, M.; Hubdar, A.K.; Yang, Q. Transgenic potato plants expressing StoVe1 exhibit enhanced resistance to Verticillium dahliae. Plant. Mol. Biol. Rep. 2012, 30, 1032–1039. [Google Scholar] [CrossRef]
- Yang, L.; Xie, C.; Liu, W.; Zhang, R.J.; Jue, D.W.; Yang, Q. Expression of a wild eggplant ribosomal protein L13a in potato enhances resistance to Verticillium dahliae. Plant. Cell Tiss. Org. 2013, 115, 329–340. [Google Scholar] [CrossRef]
- Jue, D.W.; Yang, L.; Shi, C.; Chen, M.; Yang, Q. Cloning and characterization of a Solanum torvum NPR1 gene involved in regulating plant resistance to Verticillium dahliae. Acta. Physiol. Plant. 2014, 36, 2999–3011. [Google Scholar]
- Yang, L.; Shi, C.; Mu, X.Y.; Liu, C.; Shi, K.; Zhu, W.J.; Yang, Q. Cloning and expression of a wild eggplant cytochrome P450 gene, StoCYP77A2, involved in plant resistance to Verticillium dahliae. Plant Biotechnol. Rep. 2015, 9, 167–177. [Google Scholar] [CrossRef]
- Zhou, X.; Bao, S.; Liu, J.; Yong, Z. De novo sequencing and analysis of the transcriptome of the wild eggplant species Solanum aculeatissimum in response to Verticillium dahliae. Plant Mol. Biol. Rep. 2016, 34, 1–11. [Google Scholar] [CrossRef]
- Navarro, L.; Dunoyer, P.; Jay, F.; Arnold, B.; Dharmasiri, N.; Estelle, M.; Voinnet, O.; Jones, J.D. A plant miRNA contributes to antibacterial resistance by repressing auxin signaling. Science 2006, 312, 436–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Zhang, Q.Q.; Zhang, J.G.; Wu, L.; Qi, Y.J.; Zhou, J.M. Identification of microRNAs involved in pathogen-associated molecular pattern-triggered plant innate immunity. Plant Physiol. 2010, 152, 2222–2231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, F.; Pignatta, D.; Bendix, C.; Brunkard, J.O.; Cohn, M.M.; Tung, J.; Sun, H.Y.; Kumar, P.; Baker, B. MicroRNA regulation of plant innate immune receptors. Proc. Natl. Acad. Sci. USA 2012, 109, 1790–1795. [Google Scholar] [CrossRef] [Green Version]
- Shivaprasad, P.V.; Chen, H.M.; Patel, K.; Bond, D.M.; Santos, B.; Baulcombe, D.C. A MicroRNA Superfamily Regulates Nucleotide Binding Site–Leucine-Rich Repeats and Other mRNAs. Plant Cell 2012, 24, 859–874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Q.H.; Fan, L.; Yang, L.; Hao, X.; Llewellyn, D.; Wilson, I. miR482 regulation of NBS-LRR defense genes during fungal pathogen infection in cotton. PLoS ONE 2013, 8, e84390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.; Jue, D.; Li, W.; Zhang, R.; Chen, M.; Yang, Q. Identification of miRNA from eggplant (Solanum melongena L.) by small RNA deep sequencing and their response to Verticillium dahliae infection. PLoS ONE 2013, 8, e72840. [Google Scholar] [CrossRef]
- Mu, X.Y.; Liu, X.R.; Cai, J.H.; Zhu, W.J.; Wang, Z.; Yang, Q.; You, X. MiR395 overexpression increases eggplant sensibility to Verticillium dahliae infection. Russ. J. Plant Physiol. 2018, 65, 203–210. [Google Scholar] [CrossRef]
- Yang, L.; Mu, X.; Liu, C.; Cai, J.; Shi, K.; Zhu, W.; Yang, Q. Overexpression of potato miR482e enhanced plant sensitivity to Verticillium dahliae infection. J. Integr. Plant Biol. 2015, 57, 1078–1088. [Google Scholar] [CrossRef]
- Kazan, K.; Manners, J.M. Linking development to defense: Auxin in plant-pathogen interactions. Trends Plant Sci. 2009, 14, 373–382. [Google Scholar] [CrossRef]
- Fu, J.; Wang, S. Insights into Auxin Signaling in Plant–Pathogen Interactions. Front. Plant Sci. 2011, 2, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robert-Seilaniantz, A.; MacLean, D.; Jikumaru, Y.; Hill, L.; Yamaguchi, S.; Kamiya, Y.; Jones, J.D.G. The microRNA miR393 re-directs secondary metabolite biosynthesis away from camalexin and towards glucosinolates. Plant J. 2011, 67, 218–231. [Google Scholar] [CrossRef]
- Wu, Y.F.; Reed, G.W.; Tian, C.Q. Arabidopsis microRNA167 controls patterns of ARF6 and ARF8 expression, and regulates both female and male reproduction. Development 2006, 133, 4211–4218. [Google Scholar] [CrossRef] [Green Version]
- Mallory, A.C.; Bartel, D.P.; Bartel, B. MicroRNA-directed regulation of Arabidopsis AUXIN RESPONSE FACTOR17 is essential for proper development and modulates expression of early auxin response genes. Plant Cell 2005, 17, 1360–1375. [Google Scholar] [CrossRef] [Green Version]
- Du, L.; Bao, C.; Hu, T.; Zhu, Q.; Hu, H.; He, Q.; Mao, W. SmARF8, a transcription factor involved in parthenocarpy in eggplant. Mol. Genet. Genom. 2016, 291, 93–105. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Tao, N.; Lin, Y.; Chen, M.; Yang, Q. Identification of miRNA targets in eggplant in response to Verticillium dahliae by degradome sequencing. Eur. J. Plant Pathol. 2021, 160. [Google Scholar] [CrossRef]
- Lu, S.; Sun, Y.H.; Shi, R.; Clark, C.; Chiang, V.L. Novel and mechanical stress-responsive microRNAs in Populus trichocarpa that are absent from Arabidopsis. Plant Cell 2005, 17, 2186–2203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, F.; Frazier, T.P.; Zhang, B. Identification and characterization of microRNAs and their targets in the bioenergy plant switchgrass (Panicum virgatum). Planta 2010, 232, 417–434. [Google Scholar] [CrossRef]
- Debernardi, J.M.; Rodriguez, R.E.; Mecchia, M.A.; Palatnik, J.F. Functional specialization of the plant miR396 regulatory network through distinct microRNA–target interactions. PLoS Genet. 2012, 8, e1002419. [Google Scholar] [CrossRef]
- Yang, L.; Wu, G.; Poethig, R.S. Mutations in the GW-repeat protein SUO reveal a developmental function for microRNA-mediated translational repression in Arabidopsis. Proc. Natl. Acad. Sci. USA 2012, 109, 315–320. [Google Scholar] [CrossRef] [Green Version]
- Fei, Y.; Wang, R.; Li, H.; Liu, S.; Zhang, H.; Huang, J. DPMIND: Degradome-based plant miRNA–target interaction and network database. Bioinformatics 2018, 34, 1618–1620. [Google Scholar] [CrossRef] [Green Version]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plantarum 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Hoyos, G.P.; Lauer, F.I.; Anderson, N.A. Early detection of Verticillium wilt resistance in a potato breeding program. Am. Potato J. 1993, 70, 535–541. [Google Scholar] [CrossRef]
- Varkonyi-Gasic, E.; Wu, R.; Wood, M.; Walton, E.F.; Hellens, R.P. Protocol: A highly sensitive RT-PCR method for detection and quantification of microRNAs. Plant Methods 2007, 3, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Hofgen, R.; Willmitzer, L. Storage of competent cells for Agrobacterium transformation. Nucleic Acids Res. 1988, 16, 9877. [Google Scholar] [CrossRef] [Green Version]
- Pantelides, I.S.; Tjamos, S.E.; Paplomatas, E.J. Ethylene perception via ETR1 is required in Arabidopsis infection by Verticillium dahliae. Mol. Plant Pathol. 2010, 11, 191–202. [Google Scholar] [CrossRef]
- Kaiser, C.; Merwe, R.; Bekker, T.F.; Labuschagne, N. In-vitro inhibition of mycelial growth of several phytopathogenic fungi, including Phytophthora cinnamomi by soluble silicon. S. Afr. Avocado Grow. Assoc. Yearb. 2005, 28, 70–74. [Google Scholar]
- Wan, F.; Pan, Y.; Li, J.; Chen, X.; Pan, Y.; Wang, Y.; Tian, S.; Zhang, X. Heterologous expression of Arabidopsis C-repeat binding factor 3 (AtCBF3) and cold-regulated 15A (AtCOR15A) enhanced chilling tolerance in transgenic eggplant (Solanum melongena L.). Plant Cell Rep. 2014, 33, 1951–1961. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Rao, K.V.M.; Sresty, T.V.S. Antioxidative parameters in the seedlings of pigeonpea ( Cajanus cajan (L.) Millspaugh) in response to Zn and Ni stresses. Plant Sci. 2000, 157, 113–128. [Google Scholar]
- Han, Y.; Zhang, J.; Chen, X.; Gao, Z.; Xuan, W.; Xu, S.; Ding, X.; Shen, W. Carbon monoxide alleviates cadmium-induced oxidative damage by modulating glutathione metabolism in the roots of Medicago sativa. New Phytol. 2008, 177, 155–166. [Google Scholar] [CrossRef]
- Luna, C.M.; Pastori, G.M.; Simon, D.; Karin, G.; Stephanie, B.; Foyer, C.H. Drought controls on H2O2 accumulation, catalase (CAT) activity and CAT gene expression in wheat. J. Exp. Bot. 2005, 56, 417–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, W.; Liu, X.; Chen, M.; Tao, N.; Tendu, A.; Yang, Q. A New MiRNA MiRm0002 in Eggplant Participates in the Regulation of Defense Responses to Verticillium Wilt. Plants 2021, 10, 2274. https://doi.org/10.3390/plants10112274
Zhu W, Liu X, Chen M, Tao N, Tendu A, Yang Q. A New MiRNA MiRm0002 in Eggplant Participates in the Regulation of Defense Responses to Verticillium Wilt. Plants. 2021; 10(11):2274. https://doi.org/10.3390/plants10112274
Chicago/Turabian StyleZhu, Wenjiao, Xinru Liu, Min Chen, Nianjiao Tao, Alexander Tendu, and Qing Yang. 2021. "A New MiRNA MiRm0002 in Eggplant Participates in the Regulation of Defense Responses to Verticillium Wilt" Plants 10, no. 11: 2274. https://doi.org/10.3390/plants10112274