
Mixtures of Biological Control Agents and Organic Additives Improve Physiological Behavior in Cape Gooseberry Plants under Vascular Wilt Disease

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Estimation of Vascular Wilt Development by AUDPC, Disease Index and Vascular Browning
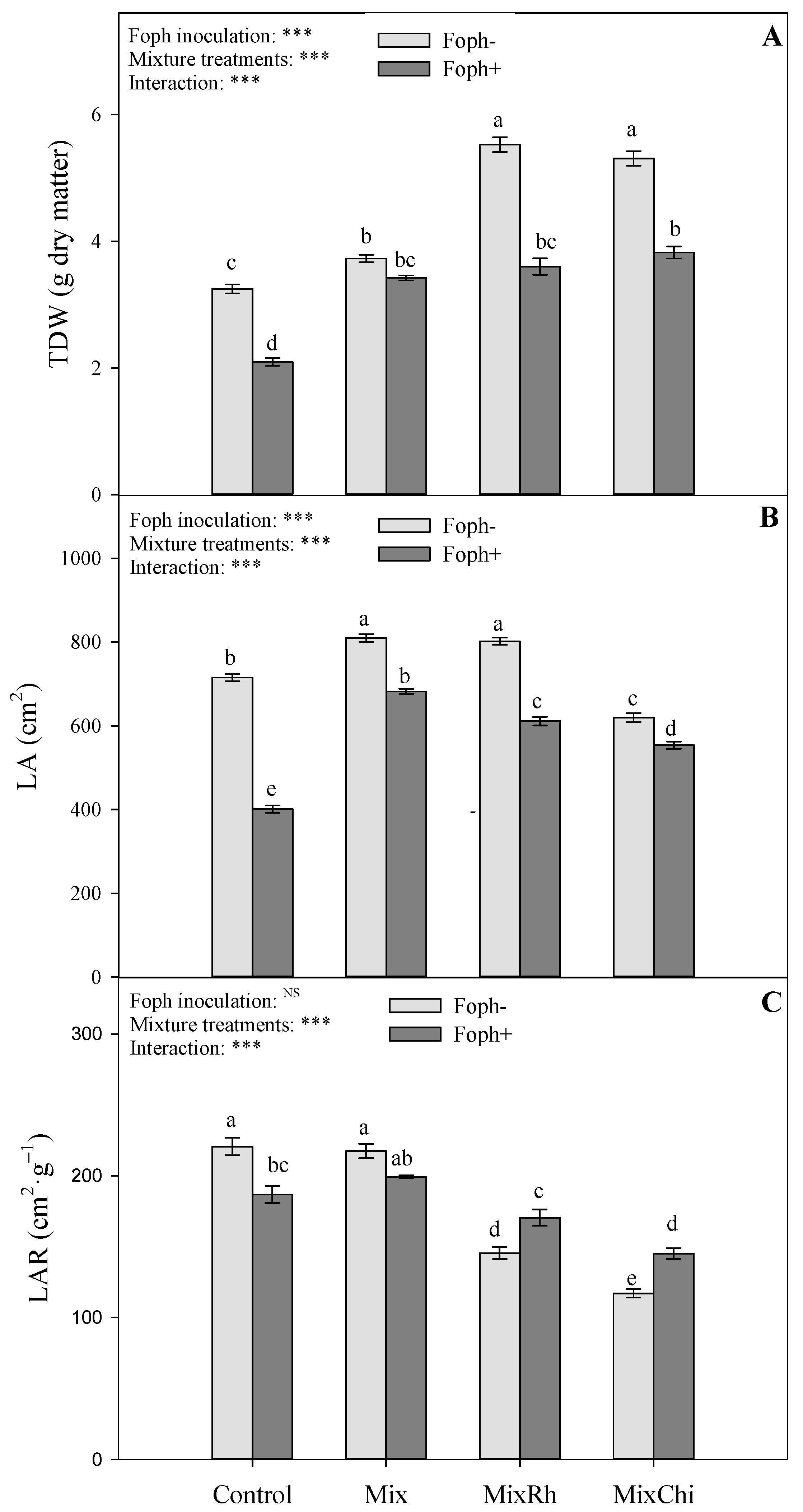
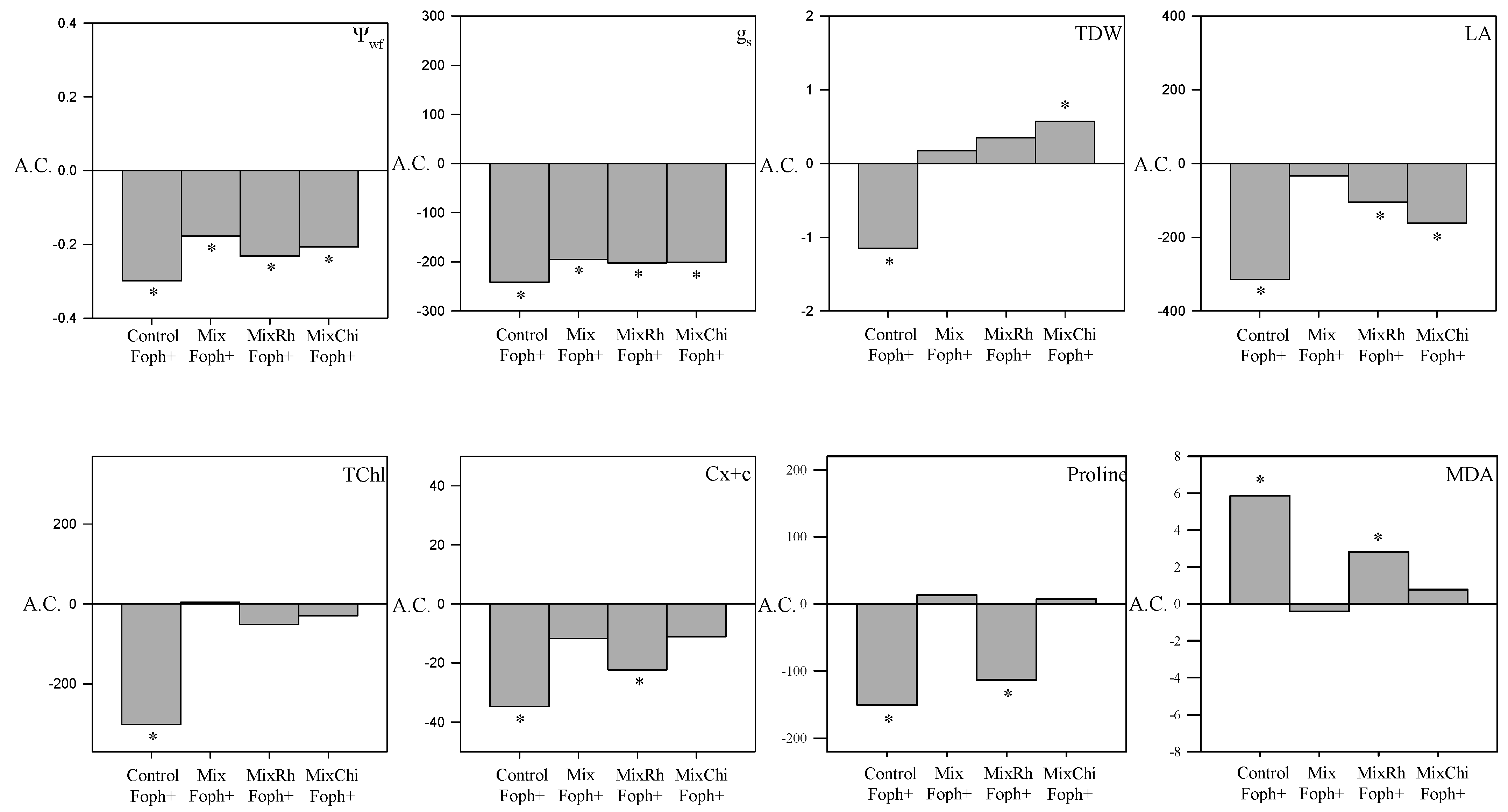
2.2. Growth Parameters
2.3. Stomatal Conductance and Leaf Water Potential
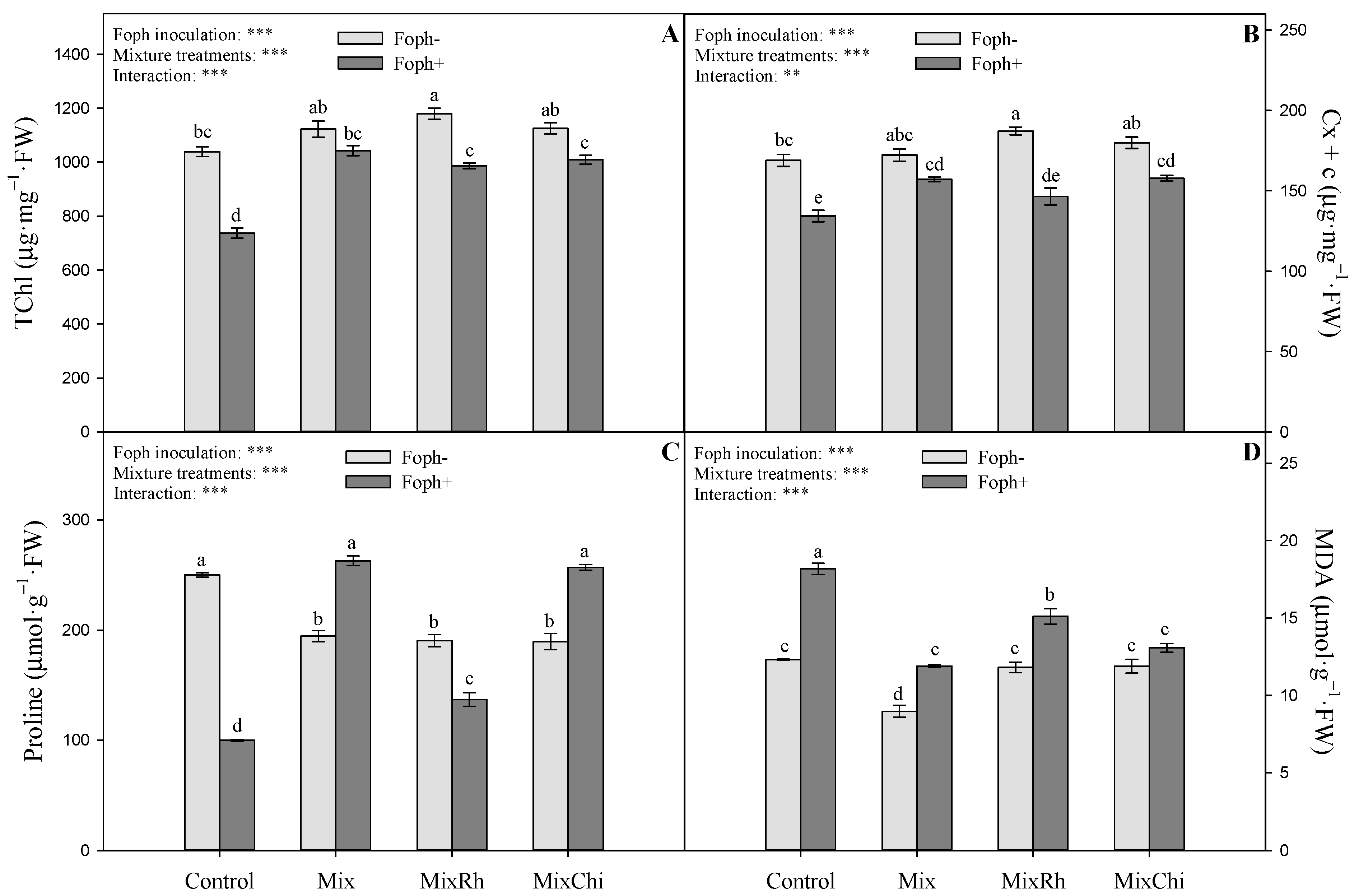
2.4. Chlorophyll and Carotenoid Content
2.5. Malondialdehyde and Proline Content
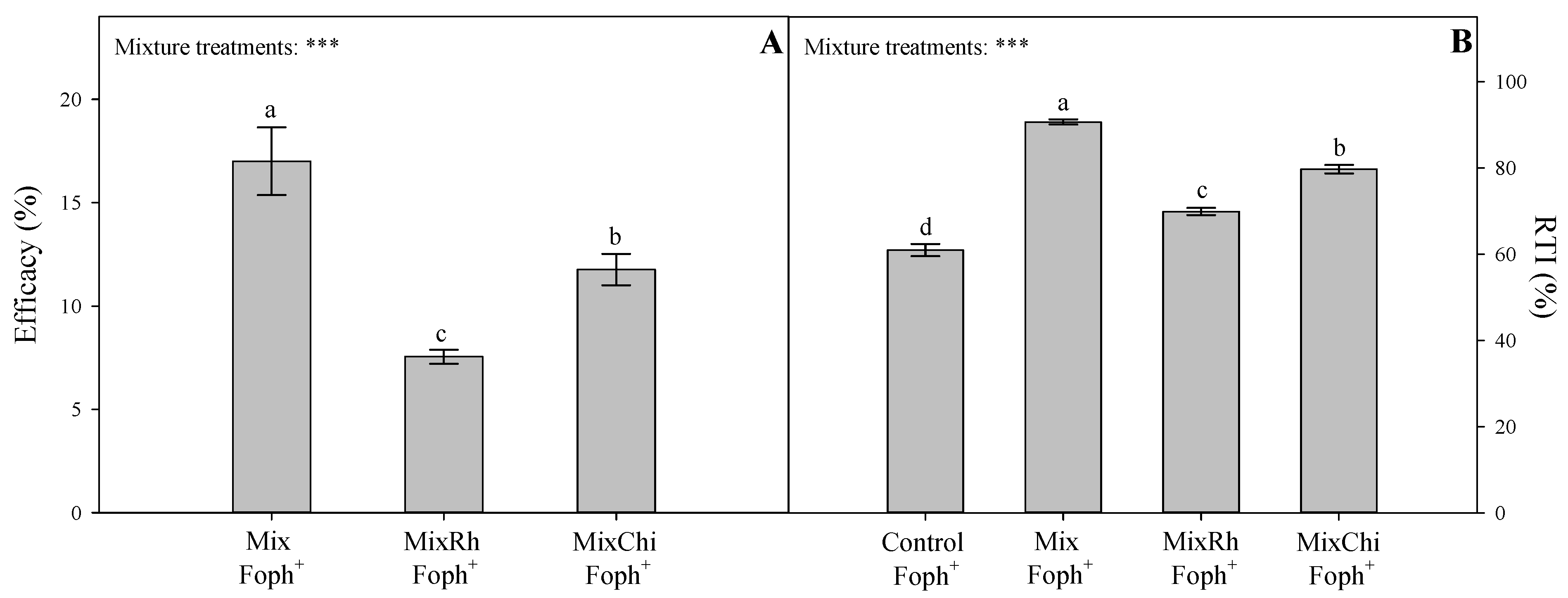
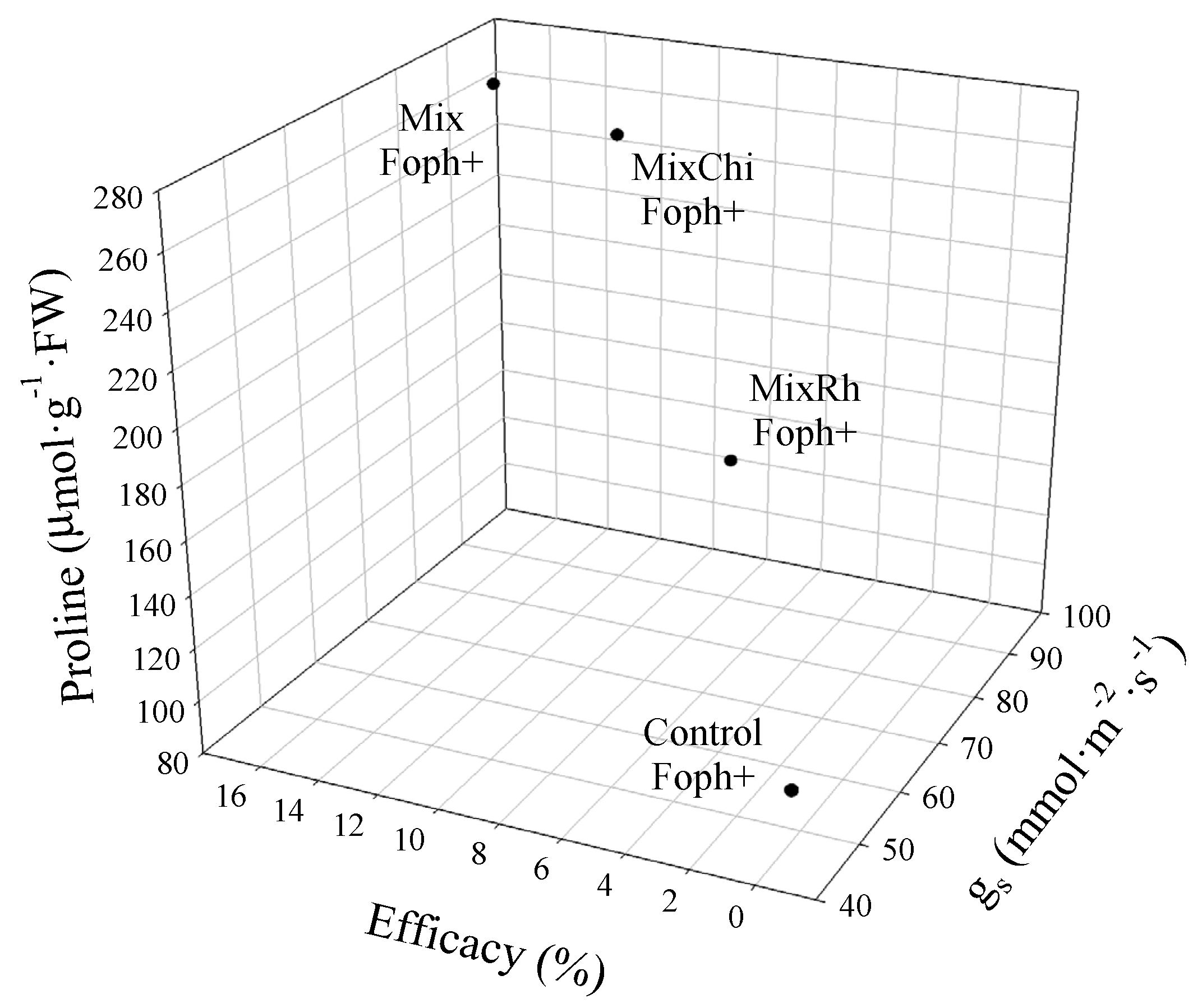
2.6. Efficacy of Mixtures of Biological Control Agents (BCAs) and Organic Additives and Relative Tolerance Index (RTI)
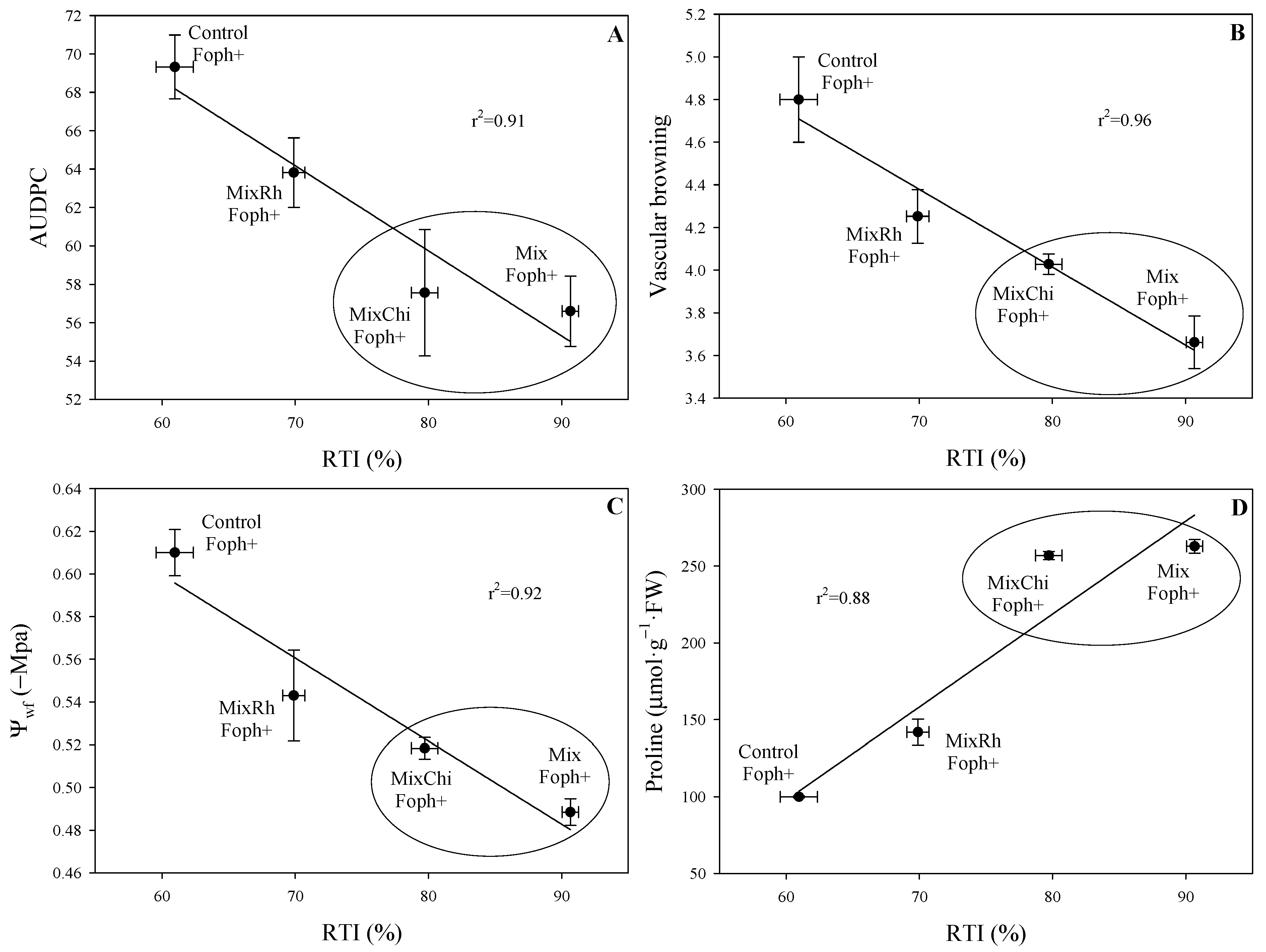
2.7. Correlation between Physiological Parameters, Disease Monitoring, and RTI
2.8. Comparative Analysis of Vascular Wilt Mitigation by the Application of Mixtures of Biological Control Agents (BCAs) and Organic Additives
3. Discussion
4. Materials and Methods
4.1. Microorganisms and Culture Conditions
4.2. Plant Material and Growth Conditions
4.3. Treatments with or without Foph Inoculation and the Addition of Mixtures of Biological Control Agents (BCAs) and Organic Additives
4.4. Disease Severity Analysis
4.5. Stomatal Conductance and Leaf Water Potential
4.6. Growth Parameters
4.7. Chlorophyll and Carotenoid Content
4.8. Malondialdehyde and Proline Content
4.9. Experimental Design and Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fischer, G.; Almanza-Merchán, P.J.; Miranda, D. Importancia y cultivo de la uchuva (Physalis peruviana L.). Rev. Bras. Frutic. 2014, 36, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Puente, L.A.; Pinto-Muñoz, C.A.; Castro, E.S.; Cortés, M. Physalis peruviana Linnaeus, the multiple properties of a highly functional fruit: A review. Food Res. Int. 2011, 44, 1733–1740. [Google Scholar] [CrossRef]
- Bravo, K.; Osorio, E. Characterization of polyphenol oxidase from Cape gooseberry (Physalis peruviana L.) fruit. Food Chem. 2016, 197, 185–190. [Google Scholar] [CrossRef]
- Agronet. Available online: http://www.agronet.gov.co/estadistica/Paginas/default.aspx (accessed on 1 August 2021).
- Osorio-Guarín, J.A.; Enciso-Rodríguez, F.E.; González, C.; Fernández-Pozo, N.; Mueller, L.A.; Barrero, L. Association analysis for disease resistance to Fusarium oxysporum in cape gooseberry (Physalis peruviana L). BMC Genom. 2016, 17, 248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okungbowa, F.I.; Shittu, H.O. Fusarium wilts: An overview. Environ. Res. J. 2012, 6, 83–102. [Google Scholar]
- Ochoa, J.; Clements, C.; Barrera, V.; Dominguez, J.M.; Ellis, M.A.; Alwang, J. IPM Packages for Naranjilla: Sustainable Production in an Environmentally Fragile Region. In Integrated Pest Management of Tropical Vegetable Crops; Muniappan, R., Heinrichs, E., Eds.; Springer: Dordrecht, The Netherlands, 2016; pp. 209–221. [Google Scholar]
- Simbaqueba, J.; Catanzariti, A.; Gonzalez, C.; Jones, D. Evidence for horizontal gene transfer and separation of effector recognition form effector function revealed by analysis of effector genes shared between cape-gooseberry and tomate-infecting formae speciales of Fusarium oxysporum. Mol. Plant Pathol. 2018, 9, 2302–2318. [Google Scholar] [CrossRef] [Green Version]
- Enciso-Rodríguez, F.E.; González, C.; Rodríguez, E.A.; López, C.E.; Landsman, D.; Barrero, L.S.; Mariño-Ramírez, L. Identification of immunity related genes to study the Physalis peruviana–Fusarium oxysporum pathosystem. PLoS ONE 2013, 8, e68500. [Google Scholar] [CrossRef] [Green Version]
- Joshi, R. A review of Fusarium oxysporum on its plant interaction and industrial use. J. Med. Plant Res. 2018, 6, 112–115. [Google Scholar] [CrossRef]
- Moreno-Velandia, C.A.; Izquierdo-García, L.F.; Ongena, M.; Kloepper, J.W.; Cotes, A.M. Soil sterilization, pathogen and antagonist concentration affect biological control of Fusarium wilt of cape gooseberry by Bacillus velezensis Bs006. Plant. Soil 2018, 435, 39–55. [Google Scholar] [CrossRef] [Green Version]
- Nogués, S.; Cotxarrera, L.; Alegre, L.; Trillas, M.I. Limitations to photosynthesis in tomato leaves induced by Fusarium wilt. New Phytol. 2002, 154, 461–470. [Google Scholar] [CrossRef]
- Dong, X.; Ling, N.; Wang, M.; Shen, Q.; Guo, S. Fusaric acid is a crucial factor in the disturbance of leaf water imbalance in Fusarium-infected banana plants. Plant Physiol. Biochem. 2012, 60, 171–179. [Google Scholar] [CrossRef]
- Wang, M.; Sun, Y.; Sun, G.; Liu, X.; Zhai, L.; Shen, Q.; Guo, S. Water balance altered in cucumber plants infected with Fusarium oxysporum f. sp. cucumerinum. Sci. Rep. 2015, 5, 7722. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Wang, M.; Li, Y.; Gu, Z.; Ling, N.; Shen, Q.; Guo, S. Wilted cucumber plants infected by Fusarium oxysporum f. sp. Cucumerinum do not suffer from water shortage. Ann. Bot. 2017, 120, 427–436. [Google Scholar] [CrossRef]
- Morkunas, I.; Bednarski, W. Fusarium oxysporum induced oxidative stress and antioxidative defenses of yellow lupine embryo axes with different level of sugars. J. Plant Physiol. 2008, 165, 262–277. [Google Scholar] [CrossRef]
- De Menezes, H.D.; Tonani, L.; Bachmann, L.; Wainwright, M.; Braga, G.Ú.L.; von Zeska Kress, M.R. Photodynamic treatment with phenothiazinium photosensitizers kills both ungerminated and germinated microconidia of the pathogenic fungi Fusarium oxysporum, Fusarium moniliforme and Fusarium solani. J. Photochem. Photobiol. B Biol. 2016, 164, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Gordon, T.R. Fusarium oxysporum and the Fusarium wilt syndrome. Annu. Rev. Phytopathol. 2017, 55, 23–39. [Google Scholar] [CrossRef] [PubMed]
- Chávez-Arias, C.C.; Gómez-Caro, S.; Restrepo-Díaz, H. Physiological, Biochemical and Chlorophyll Fluorescence Parameters of Physalis Peruviana L. Seedlings Exposed to Different Short-Term Waterlogging Periods and Fusarium Wilt Infection. Agronomy 2019, 9, 213. [Google Scholar] [CrossRef] [Green Version]
- McGovern, R.J. Management of tomato diseases caused by Fusarium oxysporum. Crop. Prot. 2015, 73, 78–92. [Google Scholar] [CrossRef]
- Pal, K.K.; Mc Spadden Gardener, B. Biological Control of Plant Pathogens. Plant Health Instr. 2006, 1–25. [Google Scholar] [CrossRef] [Green Version]
- Köhl, J.; Kolnaar, R.; Ravensberg, W.J. Mode of action of microbial biological control agents against plant diseases: Relevance beyond efficacy. Front. Plant Sci. 2019, 10, 845. [Google Scholar] [CrossRef] [Green Version]
- Ab Rahman, S.F.S.; Singh, E.; Pieterse, C.M.; Schenk, P.M. Emerging microbial biocontrol strategies for plant pathogens. Plant Sci. 2018, 267, 102–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Brien, P.A. Biological control of plant diseases. Australas. Plant Pathol. 2017, 46, 293–304. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.M.; Jeffries, P.; Pautasso, M.; Jeger, M.J. Combined use of biological control agents to management plant diseases in theory and practice. Phytopathology 2011, 101, 1024–1031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Izquierdo-García, L.F.; González-Almario, A.; Cotes, A.M.; Moreno-Velandia, C.A. Trichoderma virens Gl006 and Bacillus velezensis Bs006: A compatible interaction controlling Fusarium wilt of cape gooseberry. Sci. Rep. 2020, 10, 6857. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Robinson, J.; Jeger, M.; Jeffries, P. Using combinations of biocontrol agents to control Botrytis cinerea on strawberry leaves under fluctuating temperatures. Biocontrol Sci. Technol. 2010, 20, 359–373. [Google Scholar] [CrossRef]
- Howell, C.R. Mechanisms employed by Trichoderma species in the biological control of plant diseases: The history and evolution of current concepts. Plant Dis. 2003, 87, 4–10. [Google Scholar] [CrossRef] [Green Version]
- Cawoy, H.; Bettiol, W.; Fickers, P.; Onge, M. Bacillus-Based Biological Control of Plant Diseases. In Pesticides in the Modern World Pesticides Use and Management; IntechOpen: Rijeka, Croatia, 2011; pp. 273–302. [Google Scholar]
- López-Bucio, J.; Pelagio-Flores, R.; Herrera-Estrella, A. Trichoderma as biostimulant: Exploiting the multilevel properties of a plant beneficial fungus. Sci. Hortic. 2015, 196, 109–123. [Google Scholar] [CrossRef]
- Waghunde, R.R.; Shelake, R.M.; Sabalpara, A.N. Trichoderma: A significant fungus for agriculture and environment. Afr. J. Agric. Res. 2016, 11, 1952–1965. [Google Scholar] [CrossRef] [Green Version]
- Shafi, O.; Tian, H.; Ji, M. Bacillus species as versatile weapons for plant pathogens: A review. Biotechnol. Biotechnol. Equip. 2017, 31, 446–459. [Google Scholar] [CrossRef] [Green Version]
- Pedraza, L.A.; López, C.E.; Uribe-Velez, D. Mechanisms of action of Bacillus spp. (Bacillaceae) against phytopathogenic microorganisms during their interaction with plants. Acta Biol. Colomb. 2020, 25, 112–125. [Google Scholar] [CrossRef] [Green Version]
- Howell, C.R. Understanding the mechanisms employed by Trichoderma virens to effect biological control of cotton diseases. Phytopathology 2006, 96, 178–180. [Google Scholar] [CrossRef] [Green Version]
- Jaimes Suárez, Y.Y.; Moreno Velandia, C.A.; Cotes Prado, A.M. Induced Systemic Resistance Against Fusarium oxysporum In Tomato by Trichoderma koningiopsis Th003. Acta Biol. Colomb. 2009, 14, 111–120. [Google Scholar]
- Wang, B.B.; Yuan, J.; Zhang, J.; Shen, Z.Z.; Zhang, M.X.; Li, R.; Ruan, Y.Z.; Shen, Q.R. Effects of novel bioorganic fertilizer produced by Bacillus amyloliquefaciens W19 on antagonism of Fusarium wilt of banana. Biol. Fertil. Soils 2013, 49, 435–446. [Google Scholar] [CrossRef]
- Yuan, J.; Ruan, Y.; Wang, B.; Zhang, J.; Waseem, R.; Huang, Q.; Shen, Q. Plant growth-promoting rhizobacteria strain Bacillus amyloliquefaciens NJN-6-enriched bio-organic fertilizer suppressed Fusarium wilt and promoted the growth of banana plants. J. Agric. Food Chem. 2013, 61, 3774–3780. [Google Scholar] [CrossRef]
- Chaves-Gómez, J.L.; Chavez-Arias, C.C.; Cotes Prado, A.M.; Gómez-Caro, S.; Restrepo-Díaz, H. Physiological response of cape gooseberry seedlings to three biological control agents under Fusarium oxysporum f. sp. physali Infection. Plant Dis. 2019, 104, 388–397. [Google Scholar] [CrossRef]
- El Hadrami, A.; Adam, L.R.; El Hadrami, I.; Daayf, F. Chitosan in Plant Protection. Mar. Drugs 2010, 8, 968–987. [Google Scholar] [CrossRef]
- Hassan, O.; Chang, T. Chitosan for eco-friendly control of plant disease. Asian J. Plant Pathol. 2017, 11, 53–70. [Google Scholar] [CrossRef]
- Ways, T.M.M.; Lau, W.M.; Khutoryanskiy, V.V. Chitosan and Its Derivatives for Application in Mucoadhesive Drug Delivery Systems. Polymers 2018, 10, 267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakraborty, M.; Hasanuzzaman, M.; Rahman, M.; Khan, M.A.R.; Bhowmik, P.; Mahmud, N.U.; Tanveer, M.; Islam, T. Mechanism of Plant Growth Promotion and Disease Suppression by Chitosan Biopolymer. Agriculture 2020, 10, 624. [Google Scholar] [CrossRef]
- Al-Hetar, M.Y.; Zainal Abidin, M.A.; Sariah, M.; Wong, M.Y. Antifungal activity of chitosan against Fusarium oxysporum f. sp. cubense. J. Appl. Polym. Sci. 2011, 120, 2434–2439. [Google Scholar] [CrossRef]
- Berger, L.R.R.; Stamford, N.P.; Willadino, L.G.; Laranjeira, D.; de Lima, M.A.B.; Malheiros, S.M.M.; de Oliveira, W.J.; Stamford, T.C.M. Cowpea resistance induced against Fusarium oxysporum f. sp. tracheiphilum by crustaceous chitosan and by biomass and chitosan obtained from Cunninghamella elegans. Biol. Control 2016, 92, 45–54. [Google Scholar] [CrossRef]
- Chaves-Gómez, J.L.; Cotes-Prado, A.M.; Gómez-Caro, S.; Restrepo-Díaz, H. Physiological response of cape gooseberry seedlings to two organic additives and their mixture under inoculation with Fusarium oxysporum f. sp. physali. HortScience 2020, 55, 55–62. [Google Scholar] [CrossRef]
- Bonanomi, G.; Ippolito, F.; Scala, F. A black future for plant pathology? Biochar as a new soil amendment for controlling plant diseases. J. Plant Pathol. 2015, 97, 223–234. [Google Scholar]
- Eo, J.; Park, K.-C.; Kim, M.-H.; Kwon, S.-I.; Song, Y.-J. Effects of rice husk and rice husk biochar on root rot disease of ginseng (Panax ginseng) and on soil organisms. Biol. Agric. Hortic. 2018, 34, 27–39. [Google Scholar] [CrossRef]
- Somapala, K.; Weerahewa, D.; Thrikawala, S. Silicon rich rice hull amended soil enhances anthracnose resistance in tomato. Proced. Food Sci. 2016, 6, 190–193. [Google Scholar] [CrossRef] [Green Version]
- Ratnayake, R.M.R.N.K.; Ganehenege, M.; Ariyarathne, H.; Daundasekera, W. Soil application of rice husk as a natural silicon source to enhance some chemical defense responses against foliar fungal pathogens and growth performance of Bitter Gourd (Momordica charantia L.). Ceylon J. Sci. 2018, 47, 49–55. [Google Scholar] [CrossRef] [Green Version]
- Sid Ahmed, A.; Ezziyyani, M.; Pérez Sánchez, C.; Candela, M.E. Effect of chitin on biological control activity of Bacillus spp. and Trichoderma harzianum against root rot disease in pepper (Capsicum annuum) plants. Eur. J. Plant Pathol. 2003, 109, 633–637. [Google Scholar] [CrossRef]
- Nitu, N.J.; Masum, M.I.; Jannat, R.; Sultana, S.; Bhuiyan, K.A. Application of chitosan and Trichoderma against soil-borne pathogens and their effect on yield of tomato (Solanum lycopersicum L.). Int. J. Biosci. 2016, 9, 10–24. [Google Scholar]
- Benhamou, N.; Kloepper, J.W.; Tuzun, S. Induction of resistance against Fusarium wilt of tomato by combination of chitosan with an endophytic bacterial strain: Ultrastructure and cytochemistry of the host response. Planta 1998, 204, 153–168. [Google Scholar] [CrossRef]
- Hashem, A.; Tabassum, B.; Abd_Allah, E.F. Bacillus subtilis: A plant-growth promoting rhizobacterium that also impacts biotic stress. Saudi J. Biol. Sci. 2019, 26, 1291–1297. [Google Scholar] [CrossRef]
- Mendoza-Vargas, L.A.; Villamarín-Romero, W.P.; Cotrino-Tierradentro, A.S.; Ramírez-Gil, J.G.; Chávez-Arias, C.C.; Restrepo-Díaz, H.; Gómez-Caro, S. Physiological Response of Cape Gooseberry Plants to Fusarium oxysporum f. sp. physali, Fusaric Acid, and Water Deficit in a Hydrophonic System. Front. Plant Sci. 2021, 12, 702842. [Google Scholar] [CrossRef]
- Christopher, D.J.; Raj, T.S.; Rani, S.U.; Udhayakumar, R. Role of defense enzymes activity in tomato as induced by Trichoderma virens against Fusarium wilt caused by Fusarium oxysporum f.sp. lycopersici. J. Biopestic. 2010, 3, 158–162. [Google Scholar]
- Bell, A.A.; Hubbard, J.C.; Liu, L.; Davis, R.M.; Subbarao, K.V. Effects of chitin and chitosan on the incidence and severity of Fusarium yellows in celery. Plant Dis. 1998, 82, 322–328. [Google Scholar] [CrossRef] [Green Version]
- Casals, C.; Elmer, P.A.G.; Viñas, I.; Teixidó, N.; Sisquella, M.; Usall, J. The combination of curing with either chitosan or Bacillus subtilis CPA-8 to control brown rot infections caused by Monilinia fructicola. Postharvest Biol. Technol. 2012, 64, 126–132. [Google Scholar] [CrossRef]
- El-Mohamedy, R.S.R.; Abdel-Kareem, F.; Jabnoun-Khiareddine, H.; Daami-Remadi, M. Chitosan and Trichoderma harzianum as fungicide alternatives for controlling Fusarium crown and root rot of tomato. Tunis. J. Plant Prot. 2014, 9, 31–43. [Google Scholar]
- Bakeer, A.R.T.; El-Mohamedy, R.S.; Saied, N.M.; Abd-El-Kareem, F. Field Suppression of Fusarium Soil Borne Diseases of Tomato Plants by the Combined Application of Bio Agents and Chitosan. Br. Biotechnol. J. 2016, 13, 1–10. [Google Scholar] [CrossRef]
- Ruano-Rosa, D.; Mercado-Blanco, J. Combining biocontrol agents and organics amendments to manage soil-borne phytopathogens. In Organic Amendments and Soil Suppressiveness in Plant Disease Management; Soil Biology; Meghvansi, M.K., Varma, A., Eds.; Springer International Publishing: Cham, Switzerland, 2015; pp. 457–478. [Google Scholar]
- Okorski, A.; Olszewski, J.; Głowacka, K.; Okorska, S.; Pszczółkowska, A. The effect of the application of the biological control agent EM1 on gas exchange parameters and productivity of Pisum sativum L. infected with Fusarium oxysporum Schlecht. ACTA Agrobot. 2012, 63, 105–115. [Google Scholar] [CrossRef]
- Pantelides, I.; Tjamos, S.; Pappa, S.; Kargakis, M.; Paplomatas, E.J. The ethylene receptor ETR1 is required for Fusarium oxysporum pathogenicity. Plant Pathol. 2013, 62, 1302–1309. [Google Scholar] [CrossRef]
- Zaim, S.; Bekkar, A.A.; Belabid, L. Efficacy of Bacillus subtilis and Trichoderma harzianum combination on chickpea Fusarium wilt caused by F. oxysporum f. sp. ciceris. J. Plant Prot. Res. 2018, 53, 177–183. [Google Scholar] [CrossRef]
- Ayala, A.; Muñoz, M.F.; Argüelles, S. Lipid peroxidation: Production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxid. Med. Cell Longev. 2014, 2014, 360438. [Google Scholar] [CrossRef]
- Dar, M.I.; Naikoo, M.I.; Rehman, F.; Naushin, F.; Khan, F.A. Proline accumulation in plants: Roles in stress tolerance and plant development. In Osmolytes and Plants Acclimation to Changing Environment: Emerging Omics Technologies; Springer: New Delhi, India, 2016; pp. 155–166. [Google Scholar]
- Pérez-Bueno, M.L.; Pineda, M.; Barón, M. Phenotyping plant responses to biotic stress by chlorophyll fluorescence imaging. Front. Plant Sci. 2019, 10, 1135. [Google Scholar] [CrossRef] [PubMed]
- Alamri, S.A. The synergistic effect of two formulated biofungicides in the biocontrol of root and bottom rot of lettuce. Biocontrol Sci. 2014, 19, 189–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, S.; Jagadeesh, K.S.; Krishnaraj, P.U.; Prem, S. Biocontrol of tomato leaf curl virus (ToLCV) in tomato with chitosan supplemented formulations of Pseudomonas sp. under field conditions. Aust. J. Crop Sci. 2014, 8, 347–355. [Google Scholar]
- Zhang, H.Y.; Ge, L.L.; Chen, K.P.; Zhao, L.N.; Zhang, X.Y. Enhanced biocontrol activity of Rhodotorula mucilaginosa cultured in media containing chitosan against postharvest diseases in strawberries: Possible mechanisms underlying the effect. J. Agric. Food Chem. 2014, 62, 4214–4224. [Google Scholar] [CrossRef]
- Mukherjee, M.; Mukherjee, P.K.; Horwitz, B.A.; Zachow, C.; Berg, G.; Zeilinger, S. Trichoderma–Plant–Pathogen Interactions: Advances in Genetics of Biological Control. Indian J. Microbiol. 2012, 52, 522–529. [Google Scholar] [CrossRef] [Green Version]
- Nusaibah, S.A.; Musa, H. A Review report on the mechanism of Trichoderma spp. as biological control agent of the Basal Stem Rot (BSR) disease of Elaeis guineensis. In Trichoderma—The Most Widely Used Fungicide, 1st ed.; Mohammad, M.S., Sharif, U., Buhari, T.R., Eds.; IntechOpen: London, UK, 2019; pp. 79–90. [Google Scholar]
- Fan, B.; Wang, C.; Song, X.F.; Ding, X.L.; Wu, L.M.; Wu, H.J.; Gao, X.W.; Borriss, R. Bacillus velezensis FZB42 in 2018: The gram-positive model strain for plant growth promotion and biocontrol. Front. Microbiol. 2018, 9, 2491. [Google Scholar] [CrossRef] [Green Version]
- Gowtham, H.G.; Murali, M.; Singh, S.B.; Lakshmeesha, T.R.; Murthy, K.N.; Amruthesh, K.N.; Niranjana, S.R. Plant growth promoting rhizobacteria Bacillus amyloliquefaciens improves plant growth and induces resistance in chilli against anthracnose disease. Biol. Control 2018, 126, 209–217. [Google Scholar] [CrossRef]
- Rabbee, M.F.; Ali, M.S.; Choi, J.; Hwang, B.S.; Jeong, S.C.; Baek, K.-h. Bacillus velezensis: A Valuable Member of Bioactive Molecules within Plant Microbiomes. Molecules 2019, 24, 1046. [Google Scholar] [CrossRef] [Green Version]
- Gül, A.; Kidoglu, F.; Tüzel, Y.; Tüzel, I.H. Effects of nutrition and “Bacillus amyloliquefaciens” on tomato (“Solanum lycopersicum L.”) growing in perlite. Span. J. Agric. Res. 2008, 6, 422–429. [Google Scholar] [CrossRef]
- Vieira, P.M.; Santos, M.P.; Andrade, C.M.; Souza-Neto, O.A.; Ulhoa, C.J.; Aragao, F.J.L. Overexpression of an aquaglyceroporin gene from Trichoderma harzianum improves water-use efficiency and drought tolerance in Nicotiana tabacum. Plant Physiol. Biochem. 2017, 121, 38–47. [Google Scholar] [CrossRef]
- Kumar, S.M.; Chowdappa, P.; Krishna, V. Development of seed coating formulation using consortium of Bacillus subtilis OTPB1 and Trichoderma harzianum OTPB3 for plant growth promotion and induction of systemic resistance in field and horticultural crops. Indian Phytopath. 2015, 68, 25–31. [Google Scholar]
- Zeilinger, S.; Gruber, S.; Bansal, R.; Mukherjee, P.K. Secondary metabolism in Trichoderma—Chemistry meets genomics. Fungal Biol. Rev. 2016, 30, 74–90. [Google Scholar] [CrossRef]
- Doni, F.; Zain, C.R.C.M.; Isahak, A.; Fathurrahman, F.; Sulaiman, N.; Uphoff, N.; Yusoff, W.M.W. Relationships observed between Trichoderma inoculation and characteristics of rice grown under System of Rice Intensification (SRI) vs. conventional methods of cultivation. Symbiosis 2017, 72, 45–59. [Google Scholar] [CrossRef]
- Zehra, A.; Meena, M.; Dubey, M.K.; Aamir, M.; Upadhyay, R.S. Synergistic effects of plant defense elicitors and Trichoderma harzianum on enhanced induction of antioxidant defense system in tomato against Fusarium wilt disease. Bot. Stud. 2017, 58, 44. [Google Scholar] [CrossRef] [PubMed]
- Karuppiah, V.; Li, T.; Vallikkannu, M.; Chen, J. Co-cultivation of Trichoderma asperellum GDFS1009 and Bacillus amyloliquefaciens 1841 causes differential gene expression and improvement in the wheat growth and biocontrol activity. Front. Microbiol. 2019, 10, 1068. [Google Scholar] [CrossRef] [PubMed]
- Kheiri, A.; Moosawi Jorfa, S.A.; Malihipourb, A.; Saremic, H.; Nikkhahda, M. Synthesis and characterization of chitosan nanoparticles and their effect on Fusarium head blight and oxidative activity in wheat. Int. J. Biol. Macromol. 2017, 102, 526–538. [Google Scholar] [CrossRef] [PubMed]
- Bistgani, Z.E.; Siadat, S.A.; Bakhshandeh, A.; Pirbalouti, A.G.; Hashemic, M. Interactive effects of drought stress and chitosan application on physiological characteristics and essential oil yield of Thymus daenensis Celak. Crop J. 2017, 5, 407–415. [Google Scholar] [CrossRef]
- Rahman, M.; Mukta, J.A.; Sabir, A.A.; Gupta, D.R.; Mohi-ud-din, M.; Hasanuzzaman, M.; Miah, M.G.; Rahman, M.; Islam, M.T. Chitosan biopolymer promotes yield and stimulates accumulation of antioxidants in strawberry fruit. PLoS ONE 2018, 13, e0203769. [Google Scholar] [CrossRef]
- Xu, C.; Mou, B. Chitosan as Soil Amendment Affects Lettuce Growth, Photochemical Efficiency, and Gas Exchange. HortTechnology 2018, 28, 476–480. [Google Scholar] [CrossRef] [Green Version]
- Muley, A.B.; Shingote, P.R.; Patil, A.P.; Dalvi, S.G.; Suprasanna, P. Gamma radiation degradation of chitosan for application in growth promotion and induction of stress tolerance in potato (Solanum tuberosum L.). Carbohydr. Polym. 2019, 210, 289–301. [Google Scholar] [CrossRef]
- Rabêlo, V.M.; Magalhães, P.C.; Bressanin, L.A.; Carvalho, D.T. The foliar application of a mixture of semisynthetic chitosan derivatives induces tolerance to water deficit in maize, improving the antioxidant system and increasing photosynthesis and grain yield. Sci. Rep. 2019, 9, 8164. [Google Scholar] [CrossRef] [PubMed]
- Araujo, A.S.D.; Blum, L.E.B.; Figueiredo, C.C.D. Biochar and Trichoderma harzianum for the Control of Macrophomina phaseolina. Braz. Arch. Biol. Technol. 2019, 62, e19180259. [Google Scholar] [CrossRef] [Green Version]
- Akhter, A.; Hage-Ahmed, K.; Soja, G.; Steinkellner, S. Potential of Fusarium wilt-inducing chlamydospores, in vitro behaviour in root exudates and physiology of tomato in biochar and compost amended soil. Plant Soil 2016, 406, 425–440. [Google Scholar] [CrossRef] [Green Version]
- Win, K.T.; Okazaki, K.; Ookawa, T.; Yokoyama, T.; Ohwaki, Y. Influence of rice-husk biochar and Bacillus pumilus strain TUAT-1 on yield, biomass production, and nutrient uptake in two forage rice genotypes. PLoS ONE 2019, 14, e0220236. [Google Scholar] [CrossRef] [Green Version]
- Olivares, B.O.; Araya-Alman, M.; Acevedo-Opazo, C.; Rey, J.C.; Cañete-Salinas, P.; Kurina, F.G.; Balzarini, M.; Lobo, D.; Navas-Cortés, J.A.; Landa, B.B.; et al. Relationship between soil properties and banana productivity in the two main cultivation areas in venezuela. J. Soil Sci. Plant 2020, 20, 2512–2524. [Google Scholar] [CrossRef]
- Olivares, B.O.; Rey, J.C.; Lobo, D.; Navas-Cortés, J.A.; Gómez, J.A.; Landa, B.B. Fusarium Wilt of Bananas: A Review of Agro-Environmental Factors in the Venezuelan Production System Affecting Its Development. Agronomy 2021, 11, 986. [Google Scholar] [CrossRef]
- Olivares Campo, B.; Paredes, F.; Rey, J.; Lobo, D.; Galvis-Causil, S. The relationship between the normalized difference vegetation index, rainfall, and potential evapotranspiration in banana plantation of Venezuela. Sains Tanah J. Soil Sci. Agroclimatol. 2021, 18, 58–64. [Google Scholar] [CrossRef]
- Park, D. Isolation of Fusarium oxysporum from soils. Trans. Br. Mycol. Soc. 1961, 44, 119–122. [Google Scholar] [CrossRef]
- Moreno-Velandia, C. Interactions between Bacillus amyloliquefaciens Bs006, Fusarium oxysporun Map5 and Cape Gooseberry (Physalis peruviana). Ph.D. Thesis, Universidad Nacional de Colombia, Bogotá, Colombia, 25 January 2018. [Google Scholar]
- Chiang, K.S.; Liu, H.I.; Tsai, J.W.; Tsai, J.R.; Bock, C.H. A discussion on disease severity index values. Part II: Using the disease severity index for null hypothesis testing. Ann. Appl. Biol. 2017, 171, 490–505. [Google Scholar] [CrossRef]
- Campbell, C.L.; Madden, L.V. Introduction to Plant Disease Epidemiology; Wiley-Interscience: New York, NY, USA, 1990; p. 532. [Google Scholar]
- Leslie, J.F.; Summerell, B.A. The Fusarium Laboratory Manual, 1st ed.; Blackwell Publishing: Ames, IA, USA, 2006; p. 382. [Google Scholar]
- Mandal, S.; Mallick, N.; Mitra, A. Salicylic acid-induced resistance to Fusarium oxysporum f. sp. lycopersici in tomato. Plant Physiol. Biochem. 2009, 47, 642–649. [Google Scholar] [CrossRef] [PubMed]
- Abbott, W. A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- Roussos, A.P.; Denaxa, N.K.; Damvakaris, T.; Stournaras, V.; Argyrokastritis, I. Effect of alleviating products with different mode of action on physiology and yield of olive under drought. Sci. Hort. 2010, 125, 700–711. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembrans. Methods Enzymol. 1987, 148, 350–382. [Google Scholar] [CrossRef]
- Hodges, D.M.; DeLong, J.M.; Forney, C.F.; Prange, R.K. Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta 1999, 207, 604–611. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant. Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | AUDPC | Disease Severity Index | Vascular Browning |
|---|---|---|---|
| Foph+ | 69.3 a 1 | 3.20 a | 4.80 a |
| Mix/Foph+ | 56.6 b | 2.83 b | 3.66 c |
| MixRh/Foph+ | 63.8 ab | 2.97 ab | 4.25 ab |
| MixChi/Foph+ | 57.6 b | 2.95 ab | 4.02 bc |
| Significance | ** 2 | * | *** |
| CV (%) 3 | 7.28 | 4.67 | 6.47 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chaves-Gómez, J.L.; Chávez-Arias, C.C.; Prado, A.M.C.; Gómez-Caro, S.; Restrepo-Díaz, H. Mixtures of Biological Control Agents and Organic Additives Improve Physiological Behavior in Cape Gooseberry Plants under Vascular Wilt Disease. Plants 2021, 10, 2059. https://doi.org/10.3390/plants10102059
Chaves-Gómez JL, Chávez-Arias CC, Prado AMC, Gómez-Caro S, Restrepo-Díaz H. Mixtures of Biological Control Agents and Organic Additives Improve Physiological Behavior in Cape Gooseberry Plants under Vascular Wilt Disease. Plants. 2021; 10(10):2059. https://doi.org/10.3390/plants10102059
Chicago/Turabian StyleChaves-Gómez, José Luis, Cristian Camilo Chávez-Arias, Alba Marina Cotes Prado, Sandra Gómez-Caro, and Hermann Restrepo-Díaz. 2021. "Mixtures of Biological Control Agents and Organic Additives Improve Physiological Behavior in Cape Gooseberry Plants under Vascular Wilt Disease" Plants 10, no. 10: 2059. https://doi.org/10.3390/plants10102059