
Quantitative Experimental Embryology: A Modern Classical Approach

Abstract
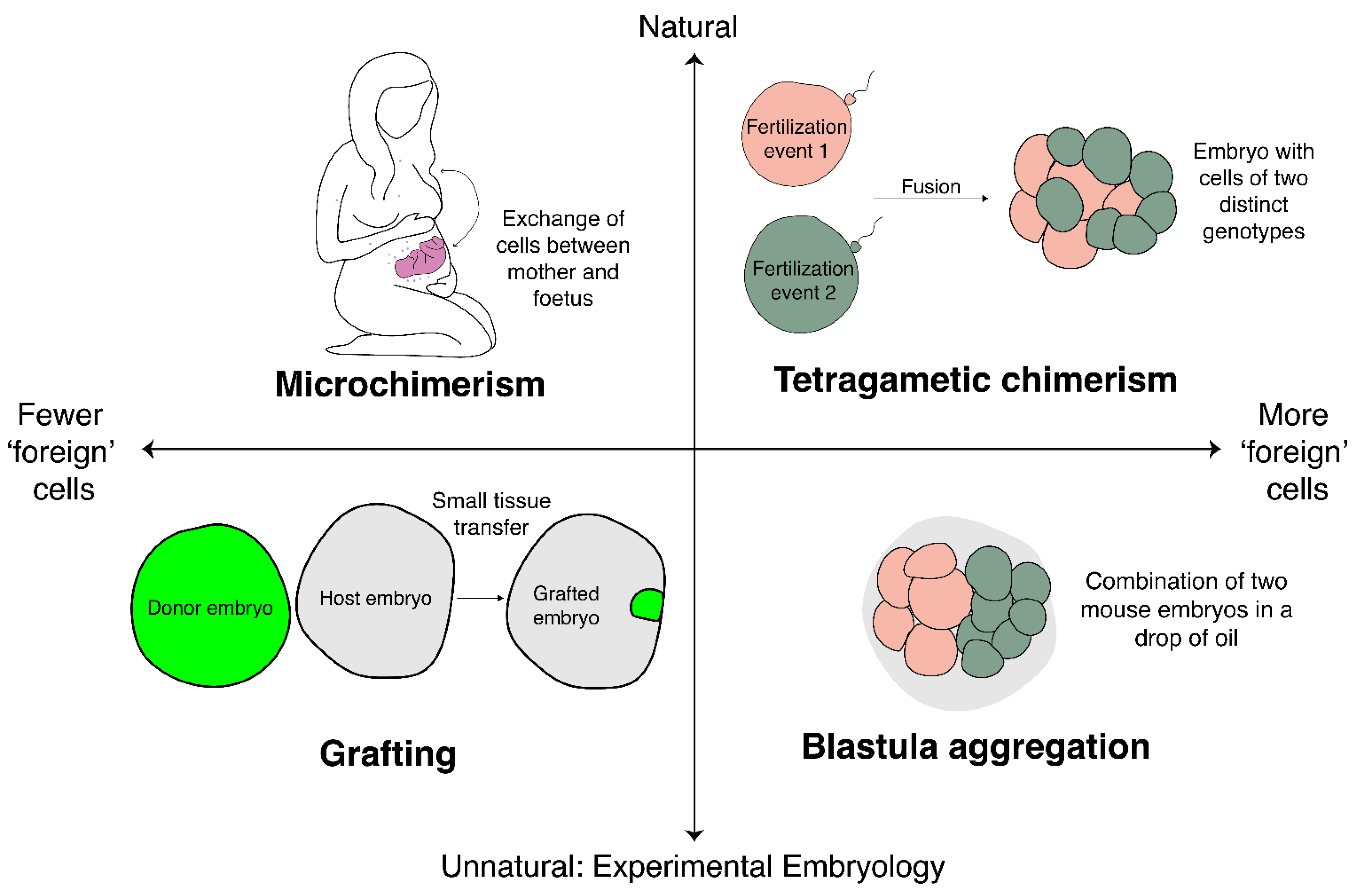
:1. Introduction
2. Cell Addition as an Essential Tool in Experimental Embryology
2.1. Chimaeras, Homotypic Grafts and Size Regulation
2.2. Blastula Aggregation
2.3. Homotypic Grafting Experiments
2.4. Inductive Reprogramming
3. Cell Removal as an Essential Tool in Experimental Embryology
4. Tissue Embedding as an Essential Tool in Experimental Embryology
4.1. Force Generation and Tissue Mechanics during Development
4.2. Intrinsic and Extrinsic Mechanical Cues in Development
4.3. Control of Mechanical and Biochemical Parameters
5. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gurdon, J.B. Embryonic induction—Molecular prospects. Development 1987, 99, 285–306. [Google Scholar] [CrossRef] [PubMed]
- Solovieva, T.; Lu, H.-C.; Moverley, A.; Plachta, N.; Stern, C.D. The embryonic node behaves as an instructive stem cell niche for axial elongation. Proc. Natl. Acad. Sci. USA 2022, 119, e2108935119. [Google Scholar] [PubMed]
- Guibentif, C.; Griffiths, J.A.; Imaz-Rosshandler, I.; Ghazanfar, S.; Nichols, J.; Wilson, V.; Göttgens, B.; Marioni, J.C. Diverse routes toward early somites in the mouse embryo. Dev. Cell 2021, 56, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, E.; Dannenberg, L.C.; Amiel, A.R.; Seaver, E.C. Regulative capacity for eye formation by first quartet micromeres of the polychaete Capitella teleta. Dev. Biol. 2016, 410, 119–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tata, P.R.; Mou, H.; Pardo-Saganta, A.; Zhao, R.; Prabhu, M.; Law, B.M.; Vinarsky, V.; Cho, J.L.; Breton, S.; Sahay, A.; et al. Dedifferentiation of committed epithelial cells into stem cells in vivo. Nature 2013, 503, 218–223. [Google Scholar] [CrossRef] [Green Version]
- Aztekin, C.; Hiscock, T.W.; Marioni, J.C.; Gurdon, J.B.; Simons, B.D.; Jullien, J. Identification of a regeneration-organizing cell in the Xenopus tail. Science 2019, 364, 653–658. [Google Scholar] [CrossRef]
- Almuedo-Castillo, M.; Bläßle, A.; Mörsdorf, D.; Marcon, L.; Soh, G.H.; Rogers, K.W.; Schier, A.F.; Müller, P. Scale-invariant patterning by size-dependent inhibition of Nodal signalling. Nat. Cell Biol. 2018, 20, 1032–1042. [Google Scholar] [CrossRef]
- Ishimatsu, K.; Hiscock, T.W.; Collins, Z.M.; Sari, D.W.K.; Lischer, K.; Richmond, D.L.; Bessho, Y.; Matsui, T.; Megason, S.G. Size-reduced embryos reveal a gradient scaling-based mechanism for zebrafish somite formation. Development 2018, 145, dev161257. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Umulis, D.M. Scale invariance of BMP signaling gradients in zebrafish. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Caldarelli, P.; Chamolly, A.; Alegria-Prévot, O.; Gros, J.; Corson, F. Self-organized tissue mechanics underlie embryonic regulation. bioRxiv 2021. [Google Scholar] [CrossRef]
- Savin, T.; Kurpios, N.A.; Shyer, A.E.; Florescu, P.; Liang, H.; Mahadevan, L.; Tabin, C.J. On the growth and form of the gut. Nature 2011, 476, 57–62. [Google Scholar] [CrossRef]
- Reinhardt, D.; Frenz, M.; Mandel, T.; Kuhlemeier, C. Microsurgical and laser ablation analysis of leaf positioning and dorsoventral patterning in tomato. Development 2005, 132, 15–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McLaren, S.B.P.; Steventon, B.J. Anterior expansion and posterior addition to the notochord mechanically coordinate zebrafish embryo axis elongation. Development 2021, 148, dev199459. [Google Scholar] [CrossRef]
- Ozelci, E.; Mailand, E.; Ruegg, M.; Sakar, M.S.; Oates, A.C. Deconstructing body axis morphogenesis in zebrafish embryos using robot-assisted tissue micromanipulation. bioRxiv 2022. [Google Scholar] [CrossRef]
- Zhou, J.; Pal, S.; Maiti, S.; Davidson, L.A. Force production and mechanical accommodation during convergent extension. Development 2015, 142, 692–701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huebner, R.J.; Weng, S.; Lee, C.; Sarıkaya, S.; Papoulas, O.; Cox, R.M.; Marcotte, E.M.; Wallingford, J.B. ARVCF catenin controls force production during vertebrate convergent extension. Dev. Cell 2022, 57, 1119–1131.e5. [Google Scholar] [CrossRef]
- Ranga, A.; Girgin, M.; Meinhardt, A.; Eberle, D.; Caiazzo, M.; Tanaka, E.M.; Lutolf, M.P. Neural tube morphogenesis in synthetic 3D microenvironments. Proc. Natl. Acad. Sci. USA 2016, 113, E6831–E6839. [Google Scholar] [CrossRef] [Green Version]
- Elosegui-Artola, A.; Gupta, A.; Najibi, A.J.; Seo, B.R.; Garry, R.; Darnell, M.; Gu, W.; Zhou, Q.; Weitz, D.A.; Mahadevan, L.; et al. Matrix viscoelasticity controls spatio-temporal tissue organization. bioRxiv 2022. [Google Scholar] [CrossRef]
- Spemann, H.; Mangold, H. über Induktion von Embryonalanlagen durch Implantation artfremder Organisatoren. Arch. Mikrosk. Anat. Und Entwickl. 1924, 100, 599–638. [Google Scholar]
- le Douarin, N. Chapter 2 Migration and Differentiation of Neural Crest Cells. Curr. Top. Dev. Biol. 1980, 16, 31–85. [Google Scholar]
- Becchio, B. Encyclopedia of World Religions; Foreign Media Group: Leeuwarden, The Netherlands, 2006. [Google Scholar]
- Dunsford, I.; Bowley, C.C.; Hutchinson, A.M.; Thompson, J.S.; Sanger, R.; Race, R.R. A human blood-group chimera. Br. Med. J. 1953, 2, 81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madan, K. Natural human chimeras: A review. Eur. J. Med. Genet. 2020, 63, 103971. [Google Scholar] [CrossRef] [PubMed]
- Tarkowski, A.K. Mouse chimaeras developed from fused eggs. Nature 1961, 190, 857–860. [Google Scholar] [CrossRef] [PubMed]
- Mintz, B. Experimental study of the developing mammalian egg: Removal of the zona pellucida. Science 1962, 138, 594–595. [Google Scholar] [CrossRef] [PubMed]
- Petters, R.M.; Mettus, R.V. Survival rate to term of chimeric morulae produced by aggregation of five to nine embryos in the mouse, musmusculus. Theriogenology 1984, 22, 167–174. [Google Scholar] [CrossRef]
- Lewis, N.E.; Rossant, J. Mechanism of size regulation in mouse embryo aggregates. Development 1982, 72, 169–181. [Google Scholar] [CrossRef]
- van Heyningen, P.; Calver, A.R.; Richardson, W.D. Control of progenitor cell number by mitogen supply and demand. Curr. Biol. 2001, 11, 232–241. [Google Scholar] [CrossRef] [Green Version]
- Bowling, S.; di Gregorio, A.; Sancho, M.; Pozzi, S.; Aarts, M.; Signore, M.; D Schneider, M.; Martinez-Barbera, J.P.; Gil, J.; Rodríguez, T.A. P53 and mTOR signalling determine fitness selection through cell competition during early mouse embryonic development. Nat. Commun. 2018, 9, 1–12. [Google Scholar]
- Montero, S.P.; Bowling, S.; Pérez-Carrasco, R.; Rodriguez, T.A. Levels of p53 expression determine the competitive ability of embryonic stem cells during the onset of differentiation. bioRxiv 2022. [Google Scholar] [CrossRef]
- Sancho, M.; Di-Gregorio, A.; George, N.; Pozzi, S.; Sánchez, J.M.; Pernaute, B.; Rodríguez, T.A. Competitive interactions eliminate unfit embryonic stem cells at the onset of differentiation. Dev. Cell 2013, 26, 19–30. [Google Scholar] [CrossRef] [Green Version]
- Nichols, J.; Lima, A.; Rodríguez, T.A. Cell competition and the regulative nature of early mammalian development. Cell Stem Cell 2022, 29, 1018–1030. [Google Scholar] [CrossRef] [PubMed]
- Bożyk, K.; Gilecka, K.; Humiecka, M.; Szpila, M.; Suwińska, A.; Tarkowski, A.K. Mouse↔ rat aggregation chimaeras can develop to adulthood. Dev. Biol. 2017, 427, 106–120. [Google Scholar] [CrossRef]
- Li, T.; Wang, L.; Zhang, X.; Jiang, L.; Li, Y.; Mao, J.; Cui, T.; Li, W.; Wang, L.; Zhou, Q. Generation of rat-mouse chimeras by introducing single cells of rat inner cell masses into mouse blastocysts. Science 2018, 1673, 8527. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Li, H.; Hu, Z.; Jiang, H.; Stablewski, A.B.; Marzullo, B.J.; Yergeau, D.A.; Feng, J. Generation of mouse–human chimeric embryos. Nat. Protoc. 2021, 16, 3954–3980. [Google Scholar] [CrossRef] [PubMed]
- Teillet, M.-A.; Ziller, C.; le Douarin, N.M. Quail—Chick Chimeras. Mol. Embryol. 2008, 461, 337–350. [Google Scholar]
- Gross, J.B.; Hanken, J.; Oglesby, E.; Marsh-Armstrong, N. Use of a ROSA26: GFP transgenic line for long-term Xenopus fate-mapping studies. J. Anat. 2006, 209, 401–413. [Google Scholar] [CrossRef] [PubMed]
- McGrew, M.J.; Sherman, A.; Lillico, S.G.; Ellard, F.M.; Radcliffe, P.A.; Gilhooley, H.J.; Mitrophanous, K.A.; Cambray, N.; Wilson, V.; Sang, H. Localised axial progenitor cell populations in the avian tail bud are not committed to a posterior Hox identity. Development 2008, 135, 2289–2299. [Google Scholar] [CrossRef] [Green Version]
- Selleck, M.A.; Stern, C.D. Fate mapping and cell lineage analysis of Hensen’s node in the chick embryo. Development 1991, 112, 615–626. [Google Scholar]
- Brown, J.M.; Storey, K.G. A region of the vertebrate neural plate in which neighbouring cells can adopt neural or epidermal fates. Curr. Biol. 2000, 10, 869–872. [Google Scholar] [CrossRef] [Green Version]
- Gray, S.D.; Dale, J.K. Notch signalling regulates the contribution of progenitor cells from the chick Hensen’s node to the floor plate and notochord. Development 2010, 137, 561–568. [Google Scholar] [CrossRef] [Green Version]
- Iimura, T.; Yang, X.; Weijer, C.J.; Pourquié, O. Dual mode of paraxial mesoderm formation during chick gastrulation. Proc. Natl. Acad. Sci. USA 2007, 104, 2744–2749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patten, I.; Kulesa, P.; Shen, M.M.; Fraser, S.; Placzek, M. Distinct modes of floor plate induction in the chick embryo. Development 2003, 130, 4809–4821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Psychoyos, D.; Stern, C.D. Fates and migratory routes of primitive streak cells in the chick embryo. Development 1996, 122, 1523–1534. [Google Scholar] [CrossRef]
- Sawada, K.; Aoyama, H. Fate maps of the primitive streak in chick and quail embryo: Ingression timing of progenitor cells of each rostro-caudal axial level of somites. Int. J. Dev. Biol. 2002, 43, 809–815. [Google Scholar]
- Schoenwolf, G.C.; Sheard, P. Fate mapping the avian epiblast with focal injections of a fluorescent-histochemical marker: Ectodermal derivatives. J. Exp. Zool. 1990, 255, 323–339. [Google Scholar] [CrossRef]
- Selleck, M.A.J.; Stern, C.D. Commitment of mesoderm cells in Hensen’s node of the to notochord and somite. Development 1992, 114, 403–415. [Google Scholar] [CrossRef]
- Saunders, J.W. Ectodermal-mesenchymal interactions in the origins of limb symmetry. Epithel. Mesenchymal Interact. 1968, 28, 78–97. [Google Scholar] [CrossRef]
- Raven, C.P.; Kloos, J. Induction by medial and lateral pieces of the archenteron roof with special reference to the determination of the neural crest. Acta Neerl. Morphol. 1945, 5, 348–362. [Google Scholar]
- Essex, L.J.; Mayor, R.; Sargent, M.G. Expression of Xenopus snail in mesoderm and prospective neural fold ectoderm. Dev. Dyn. 1993, 198, 108–122. [Google Scholar] [CrossRef]
- Bonstein, L.; Elias, S.; Frank, D. Paraxial-Fated Mesoderm Is Required for Neural Crest Induction in Xenopus Embryos. Dev. Biol. 1998, 193, 156–168. [Google Scholar] [CrossRef] [Green Version]
- Marchant, L.; Linker, C.; Ruiz, P.; Guerrero, N.; Mayor, R. The inductive properties of mesoderm suggest that the neural crest cells are specified by a BMP gradient. Dev. Biol. 1998, 198, 319–329. [Google Scholar] [CrossRef]
- Selleck, M.A.; Bronner-Fraser, M. Origins of the avian neural crest: The role of neural plate-epidermal interactions. Development 1995, 121, 525–538. [Google Scholar] [CrossRef] [PubMed]
- Steventon, B.; Araya, C.; Linker, C.; Kuriyama, S.; Mayor, R. Differential requirements of BMP and Wnt signalling during gastrulation and neurulation define two steps in neural crest induction. Development 2009, 136, 771–779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, R.M.; Candido-Ferreira, I.; Repapi, E.; Gavriouchkina, D.; Senanayake, U.; Ling, I.T.C.; Telenius, J.; Taylor, S.; Hughes, J.; Sauka-Spengler, T. Reconstruction of the global neural crest gene regulatory network in vivo. Dev. Cell 2019, 51, 255–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Čapek, D.; Müller, P. Positional information and tissue scaling during development and regeneration. Development 2019, 146, dev177709. [Google Scholar] [CrossRef]
- Spratt, N.T., Jr. Location of organ-specific regions and their relationship to the development of the primitive streak in the early chick blastoderm. J. Exp. Zool. 1942, 89, 69–101. [Google Scholar] [CrossRef]
- Spratt, N.T., Jr.; Haas, H. Integrative mechanisms in development of the early chick blastoderm. I. Regulative potentiality of separated parts. J. Exp. Zool. 1960, 145, 97–137. [Google Scholar] [CrossRef]
- Serrano Nájera, G.; Weijer, C.J. Cellular processes driving gastrulation in the avian embryo. Mech. Dev. 2020, 163, 103624. [Google Scholar] [CrossRef]
- Chuai, M.; Serrano Nájera, G.; Serra, M.; Mahadavan, L.; Weijer, C.J. Reconstruction of distinct vertebrate gastrulation modes via modulation of key cell behaviours in the chick embryo. bioRxiv 2021. [Google Scholar] [CrossRef]
- Tlili, S.; Yin, J.; Rupprecht, J.-F.; Mendieta-Serrano, M.A.; Weissbart, G.; Verma, N.; Teng, X.; Toyama, Y.; Prost, J.; Saunders, T.E. Shaping the zebrafish myotome by intertissue friction and active stress. Proc. Natl. Acad. Sci. USA 2019, 116, 25430–25439. [Google Scholar] [CrossRef] [Green Version]
- Driesch, H. Entwicklungsmechanische Studien III–VI. Z Wiss Zool 1982, 55, 1–62. [Google Scholar]
- Collinet, C.; Lecuit, T. Programmed and self-organized flow of information during morphogenesis. Nat. Rev. Mol. Cell Biol. 2021, 22, 245–265. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Kim, H.Y.; Davidson, L.A. Actomyosin stiffens the vertebrate embryo during crucial stages of elongation and neural tube closure. Development 2009, 136, 677–688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pouille, P.-A.; Ahmadi, P.; Brunet, A.-C.; Farge, E. Mechanical Signals Trigger Myosin II Redistribution and Mesoderm Invagination in Drosophila Embryos. Sci. Signal. 2009, 2, ra16. [Google Scholar] [CrossRef]
- Baral, A.; Aryal, B.; Jonsson, K.; Morris, E.; Demes, E.; Takatani, S.; Verger, S.; Xu, T.; Bennett, M.; Hamant, O.; et al. External Mechanical Cues Reveal a Katanin-Independent Mechanism behind Auxin-Mediated Tissue Bending in Plants. Dev. Cell 2021, 56, 67–80.e3. [Google Scholar] [CrossRef] [PubMed]
- Hiramatsu, R.; Matsuoka, T.; Kimura-Yoshida, C.; Han, S.W.; Mochida, K.; Adachi, T.; Takayama, S.; Matsuo, I. External mechanical cues trigger the establishment of the anterior-posterior axis in early mouse embryos. Dev. Cell 2013, 27, 131–144. [Google Scholar] [CrossRef] [Green Version]
- Steventon, B.; Busby, L.; Arias, A.M. Establishment of the vertebrate body plan: Rethinking gastrulation through stem cell models of early embryogenesis. Dev. Cell 2021, 56, 2405–2418. [Google Scholar] [CrossRef]
- McDole, K.; Guignard, L.; Amat, F.; Berger, A.; Malandain, G.; Royer, L.A.; Turaga, S.C.; Branson, K.; Keller, P.J. In toto imaging and reconstruction of post-implantation mouse development at the single-cell level. Cell 2018, 175, 859–876. [Google Scholar] [CrossRef]
- Labouesse, C.; Tan, B.X.; Agley, C.C.; Hofer, M.; Winkel, A.K.; Stirparo, G.G.; Stuart, H.T.; Verstreken, C.M.; Mulas, C.; Mansfield, W.; et al. StemBond hydrogels control the mechanical microenvironment for pluripotent stem cells. Nat. Commun. 2021, 12, 1–17. [Google Scholar] [CrossRef]
- Barriga, E.H.; Mayor, R. Adjustable viscoelasticity allows for efficient collective cell migration. Semin. Cell Dev. Biol. 2019, 93, 55–68. [Google Scholar] [CrossRef]
- Barriga, E.H.; Franze, K.; Charras, G.; Mayor, R. Tissue stiffening coordinates morphogenesis by triggering collective cell migration in vivo. Nature 2018, 554, 523–527. [Google Scholar] [CrossRef] [PubMed]
- Steventon, B.; Mayor, R.; Streit, A. Directional cell movements downstream of Gbx2 and Otx2 control the assembly of sensory placodes. Biol. Open 2016, 5, 1620–1624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Theveneau, E.; Steventon, B.; Scarpa, E.; Garcia, S.; Trepat, X.; Streit, A.; Mayor, R. Chase-and-run between adjacent cell populations promotes directional collective migration. Nat. Cell Biol. 2013, 15, 763–772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shellard, A.; Mayor, R. Collective durotaxis along a self-generated stiffness gradient in vivo. Nature 2021, 600, 690–694. [Google Scholar] [CrossRef]
- Ferreira, F.; Moreira, S.; Barriga, E.H. Stretch-induced endogenous electric fields drive neural crest directed collective cell migration in vivo. bioRxiv 2021. [Google Scholar] [CrossRef]


{kind=link}
| Experimental Embryology Techniques | Questions |
|---|---|
| Adding cells | Scaling |
| Homotypic grafts | Cell competition |
| Heterotypic grafts | Cell autonomous vs. non cell autonomous processes |
| Embryonic aggregates | Fate mapping |
| Inductive reprogramming | |
| Removing cells | Regeneration |
| Single cell removal | Scaling |
| Genetically targeted ablation | Mechanical regulation |
| Tissue dissection | Multi tissue coupling |
| Competence | |
| Confining cells | Force generation |
| Agarose gels | Intrinsic vs. extrinsic mechanical signals |
| Matrigel | Mechanical versus biochemical signals |
| Biochemically & Mechanically defined Hydrogels | Force adaptation |
| Paper | Experimental Embryology Method | Modern/Quantitative Addition |
|---|---|---|
| Solovieva et al., 2022 [2] | Grafting | Single cell RNA-sequencing and live imaging |
| Guibentif et al., 2021 [3] | Chimera production (blastula aggregation) | Single cell RNA-sequencing |
| Yamaguchi et al., 2016 [4] | Single cell ablations | Used laser ablation. |
| Tata et al., 2013 [5]; Aztekin et al., 2019 [6] | Cell population ablation | Genetically-targeted ablation. (Tet-On diptheria toxin; Nitroreductase). |
| Almuedo-Castillo, et al., 2018 [7]; Ishimatsu et al., 2018 [8]; Huang and Umulis, 2019 [9] | Tissue removal | Used classical methods to remove cells (capillary; hairloop; needle). Combined with imaging and mathematical modelling to predict how the system scales and other perturbations, such as protein-anchoring, to test the model’s predictions. |
| Caldarelli et al., 2021 [10] | Tissue removal (embryo bisection) | Used laser ablation. Coupled with high-resolution imaging to quantify the mechanical forces of tissue movement. |
| Savin et al., 2011 [11] | Tissue removal | Used classical tissue dissection followed by mathematical modelling of tissue properties. |
| Reinhardt et al., 2005 [12] | Tissue ablation | Used laser ablation. |
| McLaren and Steventon 2021 [13]; Ozelci et al., 2022 [14] | Tissue ablation | Used laser ablation. Coupled with high resolution live imaging. |
| Zhou et al., 2015 [15]; Huebner et al., 2022 [16] | Embedding | Quantification of force generation (displacement of fluorescent micro-beads and tissue buckling) |
| Ranga et al., 2016 [17] | Embedding | Robotics and automatic image analysis |
| Elosegui-Artola et al., 2022 [18] | Embedding | Computer simulations to study how fingering behaviour depends on the mechanics of the substrate |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Busby, L.; Saunders, D.; Serrano Nájera, G.; Steventon, B. Quantitative Experimental Embryology: A Modern Classical Approach. J. Dev. Biol. 2022, 10, 44. https://doi.org/10.3390/jdb10040044
Busby L, Saunders D, Serrano Nájera G, Steventon B. Quantitative Experimental Embryology: A Modern Classical Approach. Journal of Developmental Biology. 2022; 10(4):44. https://doi.org/10.3390/jdb10040044
Chicago/Turabian StyleBusby, Lara, Dillan Saunders, Guillermo Serrano Nájera, and Benjamin Steventon. 2022. "Quantitative Experimental Embryology: A Modern Classical Approach" Journal of Developmental Biology 10, no. 4: 44. https://doi.org/10.3390/jdb10040044