
Ribosomal Protein S12 Hastens Nucleation of Co-Transcriptional Ribosome Assembly

Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Fluorescent DNA Templates
2.2. Protein Purification and Fluorescent Labeling
2.3. Single-Molecule Fluorescence Microscopy
2.4. Analysis of Single-Molecule Data
3. Results
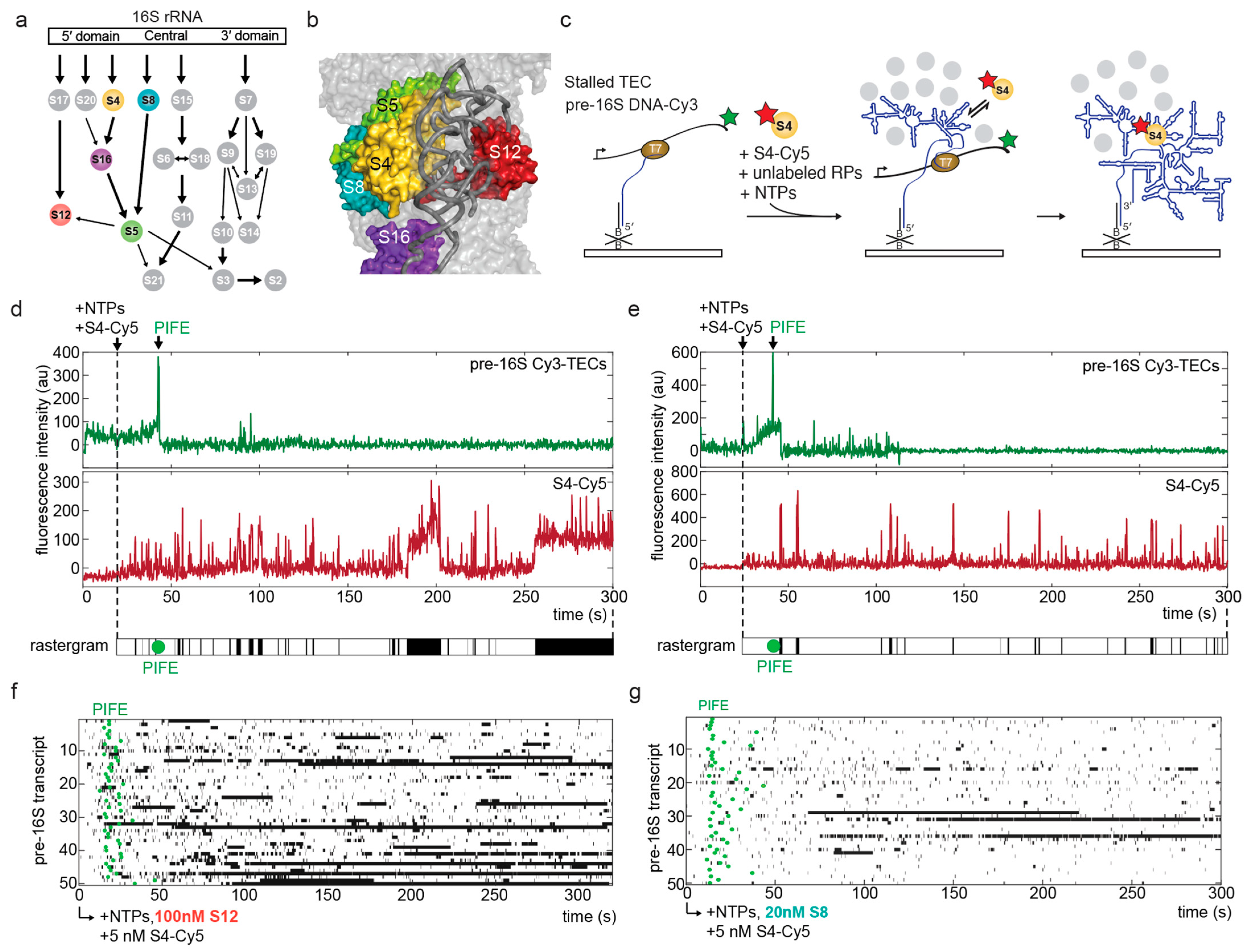
3.1. A Single-Molecule System to Study the Chaperone Activity of Ribosomal Proteins
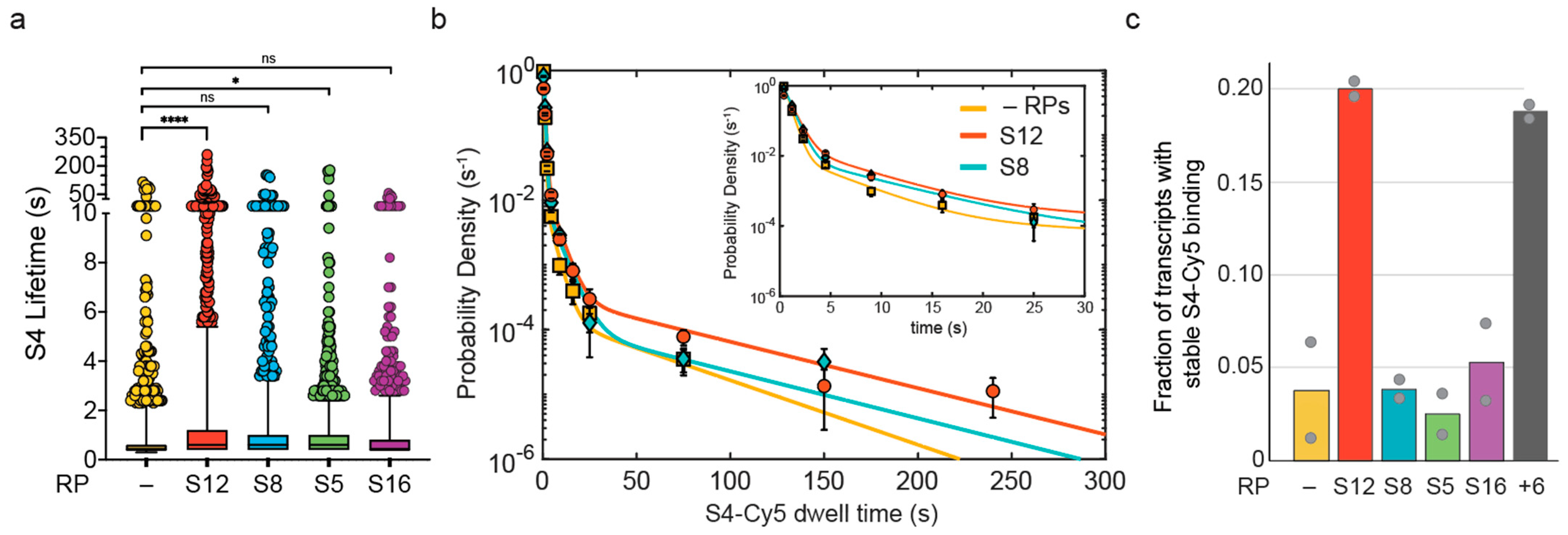
3.2. S12 Increases Likelihood of Stable S4 Recruitment to Pre-16S rRNA during Transcription
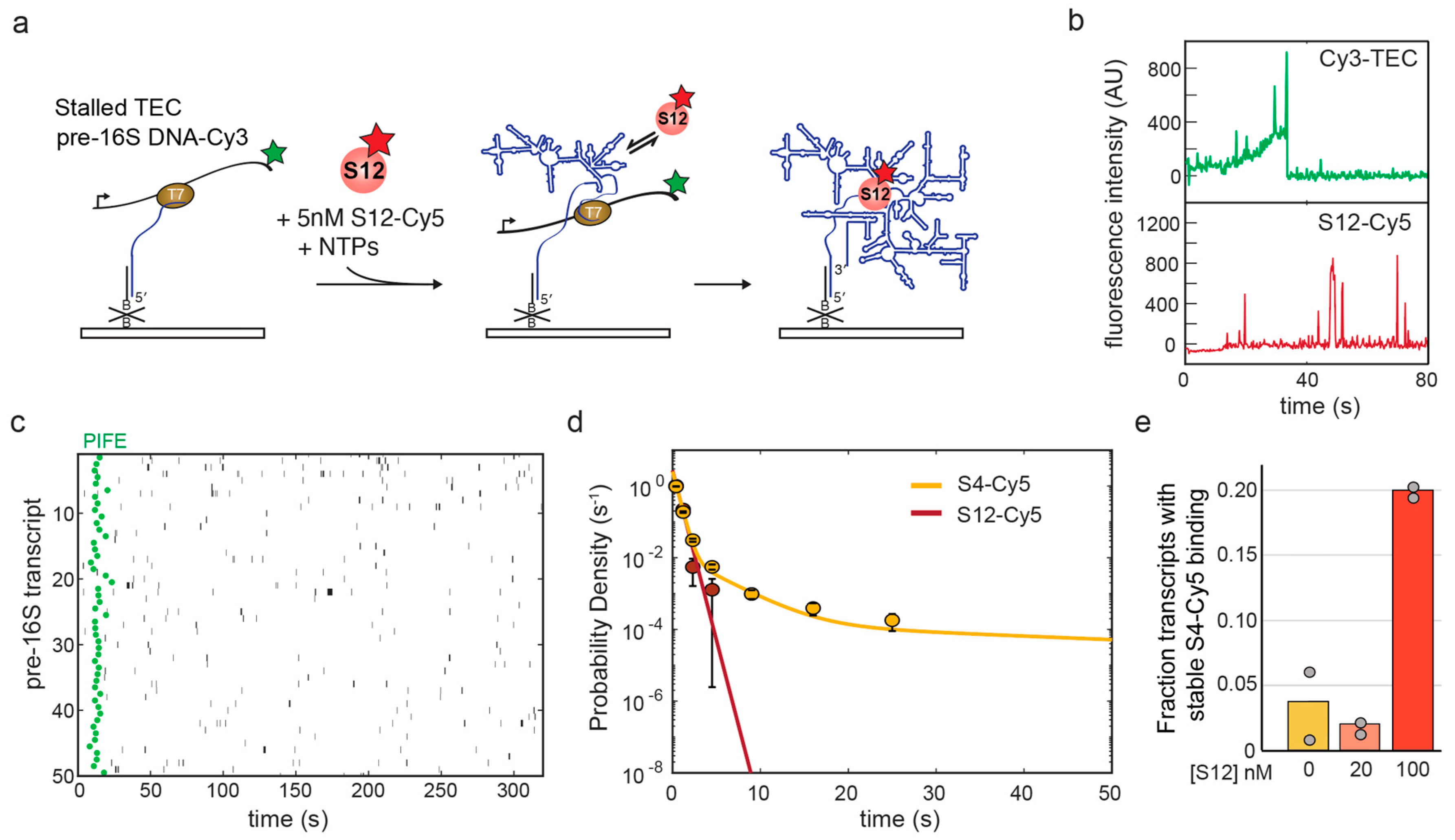
3.3. S12 Only Interacts Transiently with Pre-16S in the Absence of Other Ribosomal Proteins
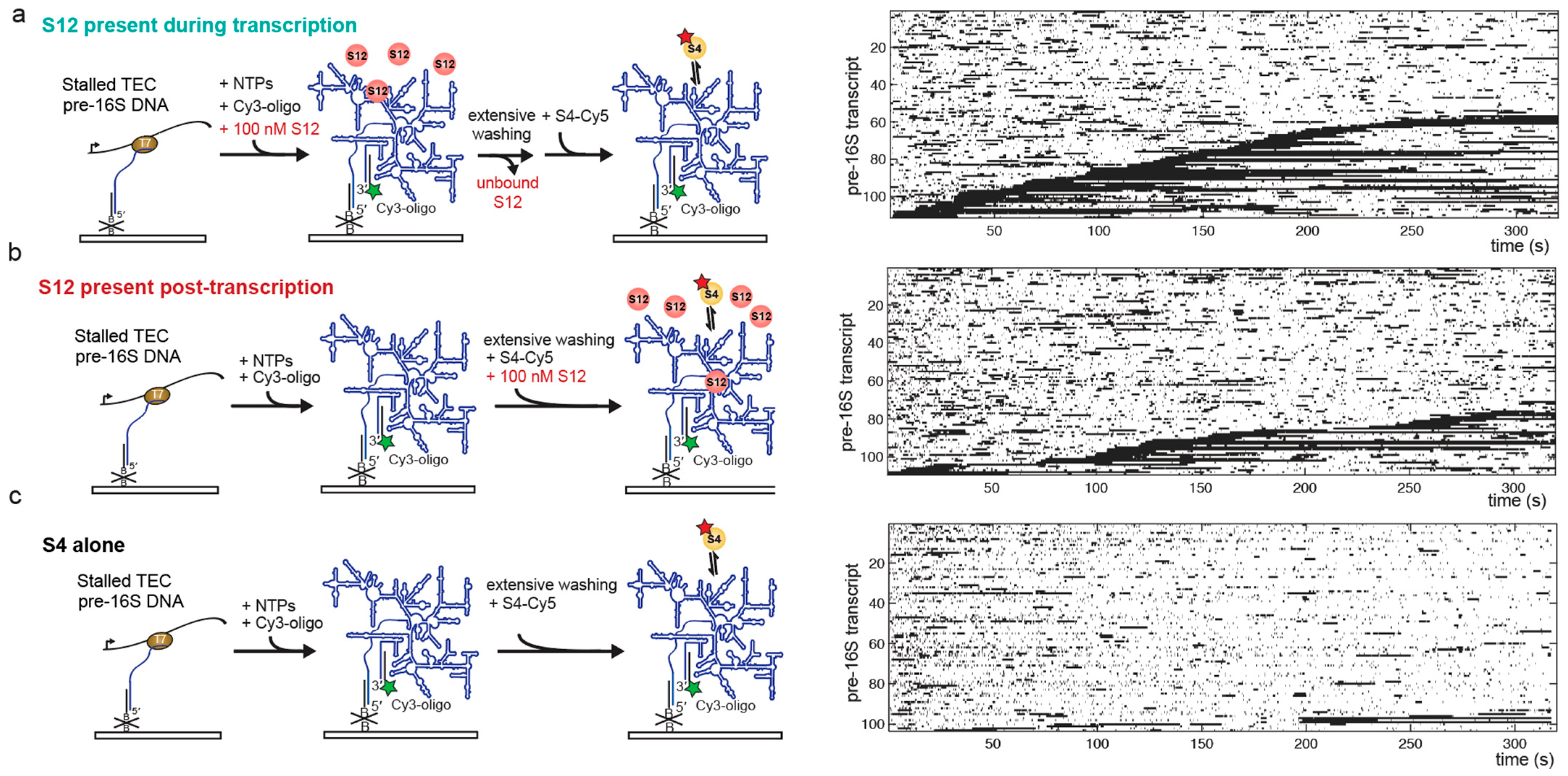
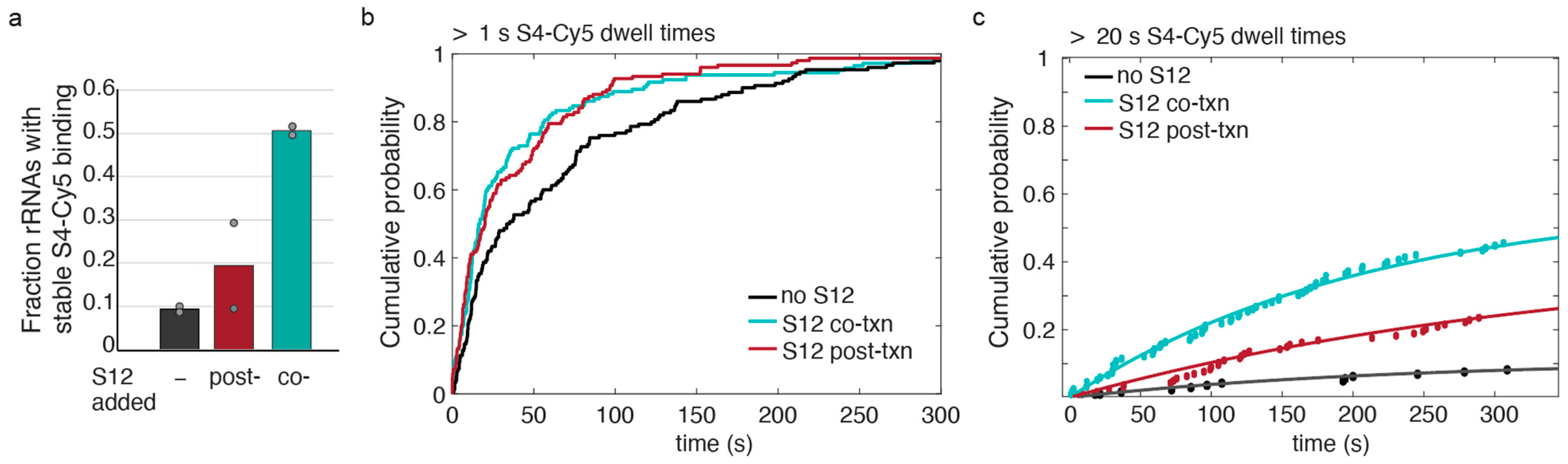
3.4. S12 Can Act on Pre-16S during and after Transcription to Increase Stable S4 Recruitment
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Noller, H.F. RNA Structure: Reading the Ribosome. Science 2005, 309, 1508–1514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stern, S.; Powers, T.; Changchien, L.-M.; Noller, H.F. RNA-Protein Interactions in 30S Ribosomal Subunits: Folding and Function of 16S rRNA. Science 1989, 244, 783–790. [Google Scholar] [CrossRef] [PubMed]
- Talkington, M.W.T.; Siuzdak, G.; Williamson, J.R. An assembly landscape for the 30S ribosomal subunit. Nature 2005, 438, 628–632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adilakshmi, T.; Bellur, D.L.; Woodson, S.A. Concurrent nucleation of 16S folding and induced fit in 30S ribosome assembly. Nature 2008, 455, 1268–1272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.; Abeysirigunawarden, S.C.; Chen, K.; Mayerle, M.; Ragunathan, K.; Luthey-Schulten, Z.; Ha, T.; Woodson, S.A. Protein-guided RNA dynamics during early ribosome assembly. Nature 2014, 506, 334–338. [Google Scholar] [CrossRef] [Green Version]
- Held, W.A.; Mizushima, S.; Nomura, M. Reconstitution of Escherichia coli 30 S Ribosomal Subunits from Purified Molecular Components. J. Biol. Chem. 1973, 248, 5720–5730. [Google Scholar] [CrossRef]
- Nowotny, V.; Nierhaus, K.H. Assembly of the 30S subunit from Escherichia coli ribosomes occurs via two assembly domains which are initiated by S4 and S7. Biochemistry 1988, 27, 7051–7055. [Google Scholar] [CrossRef]
- Ridgeway, W.K.; Millar, D.P.; Williamson, J.R. Quantitation of ten 30S ribosomal assembly intermediates using fluorescence triple correlation spectroscopy. Proc. Natl. Acad. Sci. USA 2012, 109, 13614–13619. [Google Scholar] [CrossRef] [Green Version]
- Davis, J.H.; Tan, Y.Z.; Carragher, B.; Potter, C.S.; Lyumkis, D.; Williamson, J.R. Modular Assembly of the Bacterial Large Ribosomal Subunit. Cell 2016, 167, 1610–1622.e15. [Google Scholar] [CrossRef] [Green Version]
- French, S.L.; Miller, O.L. Transcription mapping of the Escherichia coli chromosome by electron microscopy. J. Bacteriol. 1989, 171, 4207–4216. [Google Scholar] [CrossRef] [Green Version]
- Powers, T.; Daubresse, G.; Noller, H.F. Dynamics of In Vitro Assembly of 16 S rRNA into 30 S Ribosomal Subunits. J. Mol. Biol. 1993, 232, 362–374. [Google Scholar] [CrossRef] [PubMed]
- Ramaswamy, P.; Woodson, S.A. Global Stabilization of rRNA Structure by Ribosomal Proteins S4, S17, and S20. J. Mol. Biol. 2009, 392, 666–677. [Google Scholar] [CrossRef] [Green Version]
- Gerstner, R.B.; Pak, Y.; Draper, D.E. Recognition of 16S rRNA by Ribosomal Protein S4 from Bacillus stearothermophilus. Biochemistry 2001, 40, 7165–7173. [Google Scholar] [CrossRef] [PubMed]
- Sapag, A.; Vartikar, J.V.; Draper, D.E. Dissection of the 16S rRNA binding site for ribosomal protein S4. Biochim. Biophys. Acta BBA-Gene Struct. Expr. 1990, 1050, 34–37. [Google Scholar] [CrossRef]
- Vartikar, J.V.; Draper, D.E. S4-16 S ribosomal RNA complex: Binding constant measurements and specific recognition of a 460-nucleotide region. J. Mol. Biol. 1989, 209, 221–234. [Google Scholar] [CrossRef]
- Powers, T.; Noller, H.F. A Temperature-dependent Conformational Rearrangement in the Ribosomal Protein S4 16 S rRNA Complex. J. Biol. Chem. 1995, 270, 1238–1242. [Google Scholar] [CrossRef] [Green Version]
- Mayerle, M.; Bellur, D.L.; Woodson, S.A. Slow Formation of Stable Complexes during Coincubation of Minimal rRNA and Ribosomal Protein S4. J. Mol. Biol. 2011, 412, 453–465. [Google Scholar] [CrossRef] [Green Version]
- Rodgers, M.L.; Woodson, S.A. Transcription Increases the Cooperativity of Ribonucleoprotein Assembly. Cell 2019, 179, 1370–1381.e12. [Google Scholar] [CrossRef]
- Bellur, D.L.; Woodson, S.A. A Minimized rRNA-Binding Site for Ribosomal Protein S4 and Its Implications for 30S Assembly. Proc. Natl. Acad. Sci. USA 2009, 37, 1886–1896. [Google Scholar] [CrossRef] [Green Version]
- Duss, O.; Stepanyuk, G.A.; Puglisi, J.D.; Williamson, J.R. Transient Protein-RNA Interactions Guide Nascent Ribosomal RNA Folding. Cell 2019, 179, 1357–1369.e16. [Google Scholar] [CrossRef]
- Williamson, J.R. Induced fit in RNA-protein recognition. Nat. Struct. Mol. Biol. 2000, 7, 834–837. [Google Scholar] [CrossRef] [PubMed]
- Coetzee, T.; Herschlag, D.; Belfort, M. Escherichia coli proteins, including ribosomal protein S12, facilitate in vitro splicing of phage T4 introns by acting as RNA chaperones. Genes Dev. 1994, 8, 1575–1588. [Google Scholar] [CrossRef] [Green Version]
- Culver, G.M.; Noller, H.F. Efficient reconstitution of functional Escherichia coli 30S ribosomal subunits from a complete set of recombinant small subunit ribosomal proteins. RNA 1999, 5, 832–843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Culver, G.M.; Noller, H.F. In vitro reconstitution of 30S ribosomal subunits using complete set of recombinant proteins. Methods Enzym. 2000, 318, 446–460. [Google Scholar] [CrossRef]
- Davanloo, P.; Rosenberg, A.H.; Dunn, J.J.; Studier, F.W. Cloning and expression of the gene for bacteriophage T7 RNA polymerase. Proc. Natl. Acad. Sci. USA 1984, 81, 2035–2039. [Google Scholar] [CrossRef] [Green Version]
- Butler, E.T.; Chamberlin, M.J. Bacteriophage SP6-specific RNA polymerase. I. Isolation and characterization of the enzyme. J. Biol. Chem. 1982, 257, 5772–5778. [Google Scholar] [CrossRef]
- Hickerson, R.; Majumdar, Z.K.; Baucom, A.; Clegg, R.M.; Noller, H.F. Measurement of Internal Movements within the 30S Ribosomal Subunit Using Förster Resonance Energy Transfer. J. Mol. Biol. 2005, 354, 459–472. [Google Scholar] [CrossRef]
- Hua, B.; Han, K.Y.; Zhou, R.; Kim, H.; Shi, X.; Abeysirigunawardena, S.C.; Jain, A.; Singh, D.; Aggarwal, V.; Woodson, S.A.; et al. An improved surface passivation method for single-molecule studies. Nat. Methods 2014, 11, 1233–1236. [Google Scholar] [CrossRef] [Green Version]
- Friedman, L.J.; Gelles, J. Multi-wavelength single-molecule fluorescence analysis of transcription mechanisms. Methods 2015, 86, 27–36. [Google Scholar] [CrossRef] [Green Version]
- Lindahl, L. Intermediates and time kinetics of the in vivo assembly of Escherichia coli ribosomes. J. Mol. Biol. 1975, 92, 15–37. [Google Scholar] [CrossRef]
- Chen, S.S.; Sperling, E.; Silverman, J.M.; Davis, J.H.; Williamson, J.R. Measuring the dynamics of E. coli ribosome biogenesis using pulse-labeling and quantitative mass spectrometry. Mol. Biosyst. 2012, 8, 3325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Held, W.A.; Nomura, M. Rate-determining step in the reconstitution of Escherichia coli 30S ribosomal subunits. Biochemistry 1973, 12, 3273–3281. [Google Scholar] [CrossRef]
- Woodson, S.A.; Panja, S.; Santiago-Frangos, A. Proteins That Chaperone RNA Regulation. Microbiol. Spectr. 2018, 6, 385–397. [Google Scholar] [CrossRef] [PubMed]
- Tompa, P.; Csermely, P. The role of structural disorder in the function of RNA and protein chaperones. FASEB J. 2004, 18, 1169–1175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bear, D.G.; Ng, R.; Van Derveer, D.; Johnson, N.P.; Thomas, G.; Schleich, T.; Noller, H.F. Alteration of polynucleotide secondary structure by ribosomal protein S1. Proc. Natl. Acad. Sci. USA 1976, 73, 1824–1828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolb, A.; Hermoso, J.M.; Thomas, J.O.; Szer, W. Nucleic acid helix-unwinding properties of ribosomal protein S1 and the role of S1 in mRNA binding to ribosomes. Proc. Natl. Acad. Sci. USA 1977, 74, 2379–2383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hajnsdorf, E.; Boni, I.V. Multiple activities of RNA-binding proteins S1 and Hfq. Biochimie 2012, 94, 1544–1553. [Google Scholar] [CrossRef]
- Lund, P.E.; Chatterjee, S.; Daher, M.; Walter, N.G. Protein unties the pseudoknot: S1-mediated unfolding of RNA higher order structure. Nucleic Acids Res. 2020, 48, 2107–2125. [Google Scholar] [CrossRef] [Green Version]
- Andrade, J.M.; dos Santos, R.F.; Chelysheva, I.; Ignatova, Z.; Arraiano, C.M. The RNA-binding protein Hfq is important for ribosome biogenesis and affects translation fidelity. EMBO J. 2018, 37, e97631. [Google Scholar] [CrossRef]
- Brodersen, D.; Clemons, W.; Carter, A.; Wimberly, B.T.; Ramakrishnan, V. Crystal structure of the 30 s ribosomal subunit from Thermus thermophilus: Structure of the proteins and their interactions with 16 s RNA. J. Mol. Biol. 2002, 316, 725–768. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RP Added | Nmol | (s) | (s) | (s) 2 | 1 | 2 | |
|---|---|---|---|---|---|---|---|
| S4 alone | 226 | 0.47 ± 0.01 | 3.83 ± 1.01 | 44 ± 11 | 0.957 ± 0.008 | 0.037 ± 0.007 | 0.003 ± 0.008 |
| +100 nM S12 | 200 | 0.71 ± 0.02 | 4.78 ± 0.76 | 60 ± 13 | 0.893 ± 0.012 | 0.089 ± 0.011 | 0.018 ± 0.012 |
| +20 nM S8 | 176 | 0.63 ± 0.02 | 5.79 ± 1.03 | 60 ± 21 | 0.941 ± 0.009 | 0.053 ± 0.009 | 0.006 ± 0.009 |
| +100 nM S5 | 183 | 0.67 ± 0.01 | 4.43 ± 0.93 | 118 ± 29 | 0.978 ± 0.006 | 0.020 ± 0.006 | 0.002 ± 0.006 |
| +50 nM S16 | 130 | 0.52 ± 0.02 | 2.35 ± 0.52 | 16 ± 3.8 | 0.923 ± 0.014 | 0.060 ± 0.015 | 0.016 ± 0.015 |
| Experiment | Nmol | (s−1) | |
|---|---|---|---|
| S4 alone post-txn | 153 | 0.0043 ± 0.0018 | 0.11 ± 0.02 |
| S12 added co-txn | 143 | 0.0047 ± 0.0004 | 0.59 ± 0.02 |
| S12 added post-txn | 151 | 0.0028 ± 0.0012 | 0.43 ± 0.09 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodgers, M.L.; Sun, Y.; Woodson, S.A. Ribosomal Protein S12 Hastens Nucleation of Co-Transcriptional Ribosome Assembly. Biomolecules 2023, 13, 951. https://doi.org/10.3390/biom13060951
Rodgers ML, Sun Y, Woodson SA. Ribosomal Protein S12 Hastens Nucleation of Co-Transcriptional Ribosome Assembly. Biomolecules. 2023; 13(6):951. https://doi.org/10.3390/biom13060951
Chicago/Turabian StyleRodgers, Margaret L., Yunsheng Sun, and Sarah A. Woodson. 2023. "Ribosomal Protein S12 Hastens Nucleation of Co-Transcriptional Ribosome Assembly" Biomolecules 13, no. 6: 951. https://doi.org/10.3390/biom13060951