
Disease-Specific α-Synuclein Seeding in Lewy Body Disease and Multiple System Atrophy Are Preserved in Formaldehyde-Fixed Paraffin-Embedded Human Brain

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
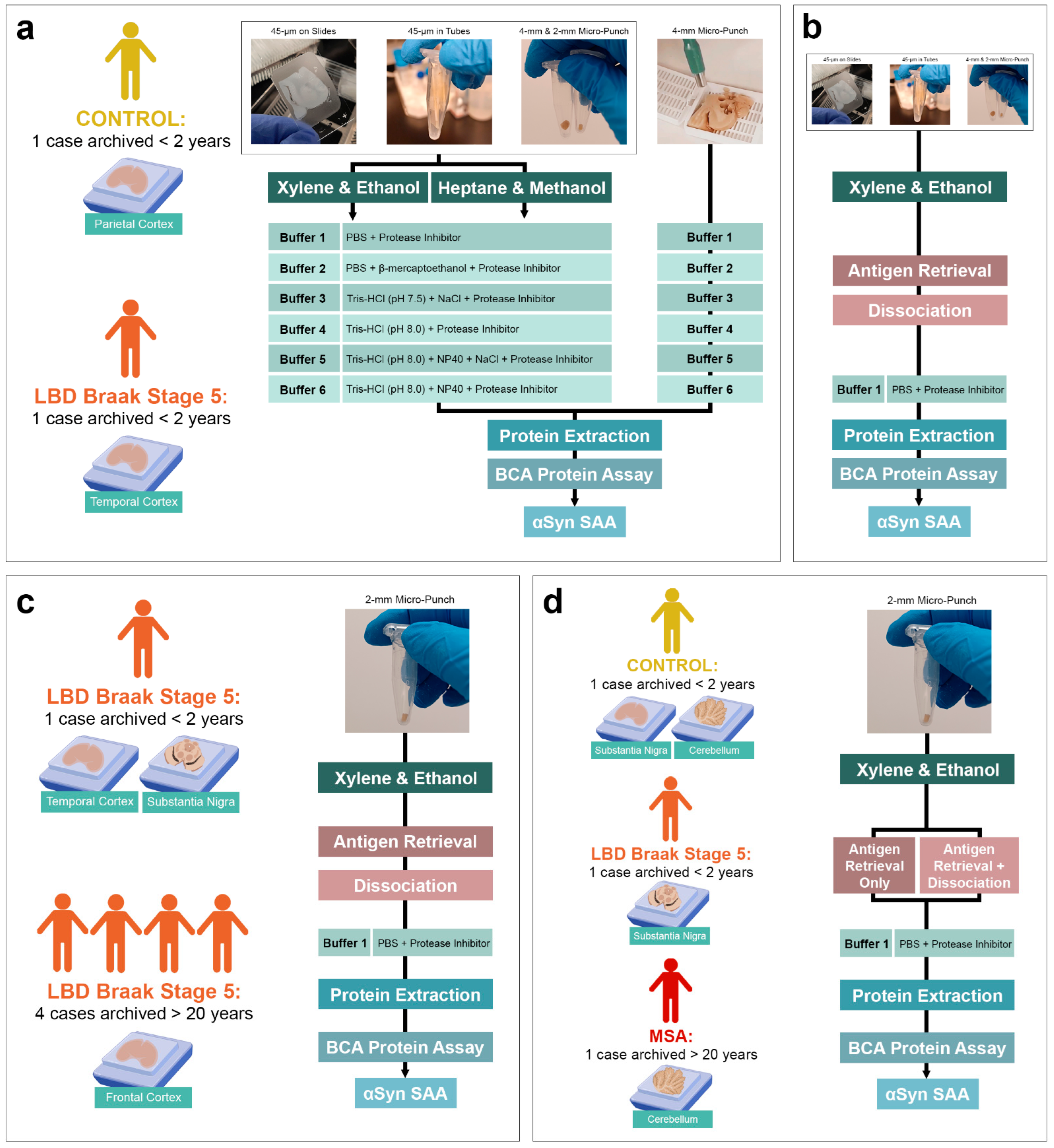
2.1. Case Selection
2.2. Immunohistochemistry
2.3. Sample Collection
2.4. Deparaffinization
2.5. Antigen Retrieval and Dissociation
2.6. Protein Extraction and Quantification
2.7. αSyn SAA Protocol
2.8. Transmission Electron Microscopy
2.9. Statistical Analysis
3. Results
3.1. Protein Extraction and SAA from FFPE Human Brain Tissue
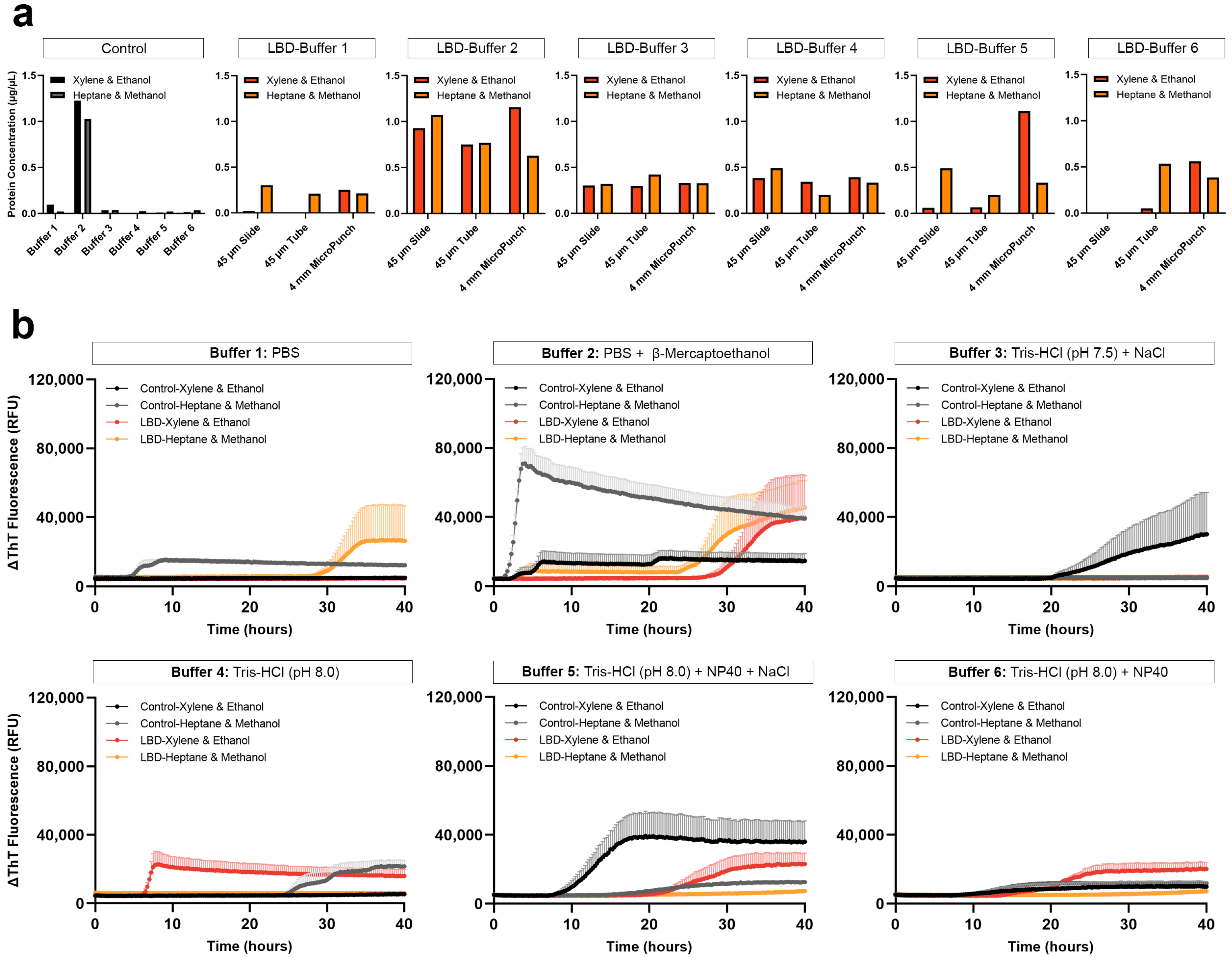
3.1.1. Comparison of Protein Extraction Efficiency Using Different Tissue Preparation, Deparaffinization and Extraction Buffers
3.1.2. Seeding of αSyn Extracted Using Different Tissue Preparation, Deparaffinization and Extraction Buffers
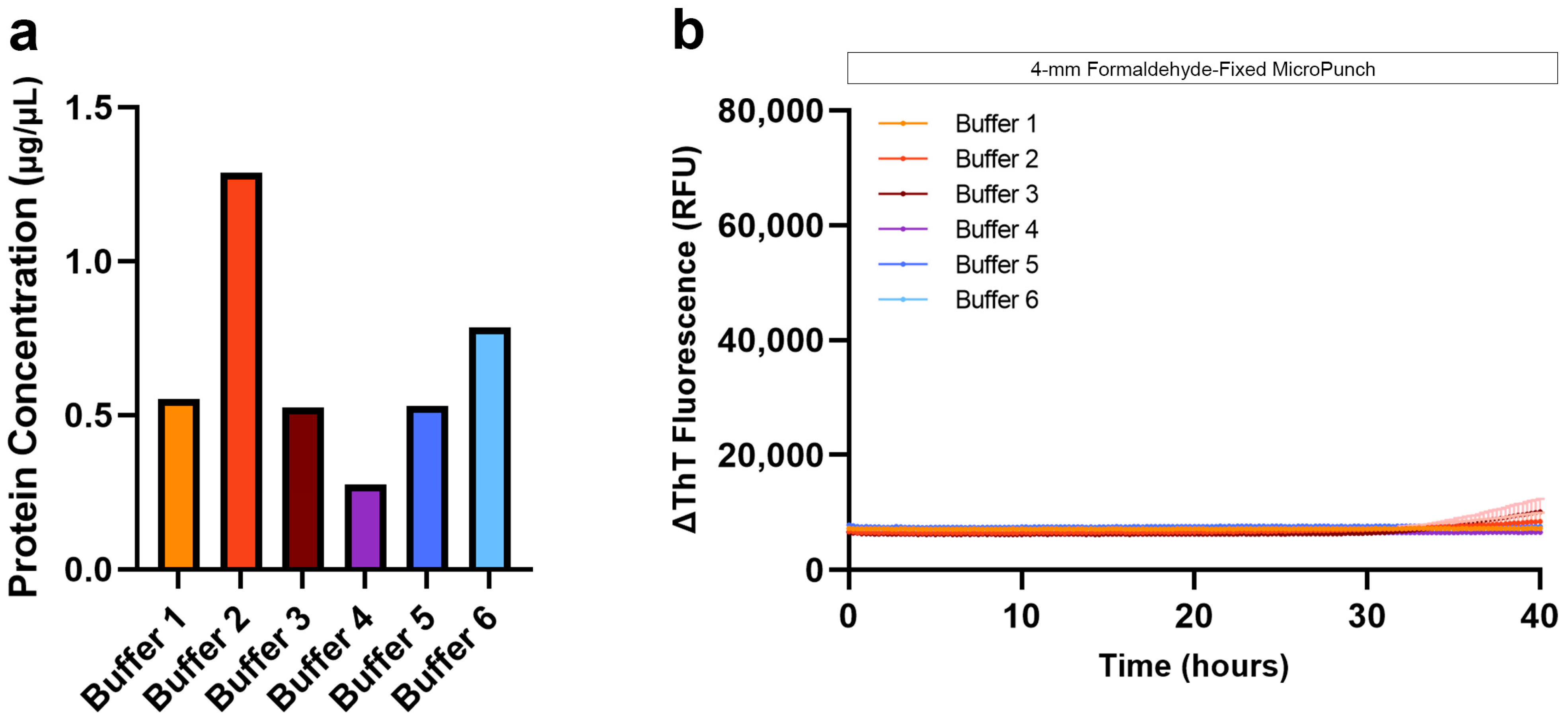
3.2. Protein Extraction and SAA from Formaldehyde-Fixed Human Brain Tissue
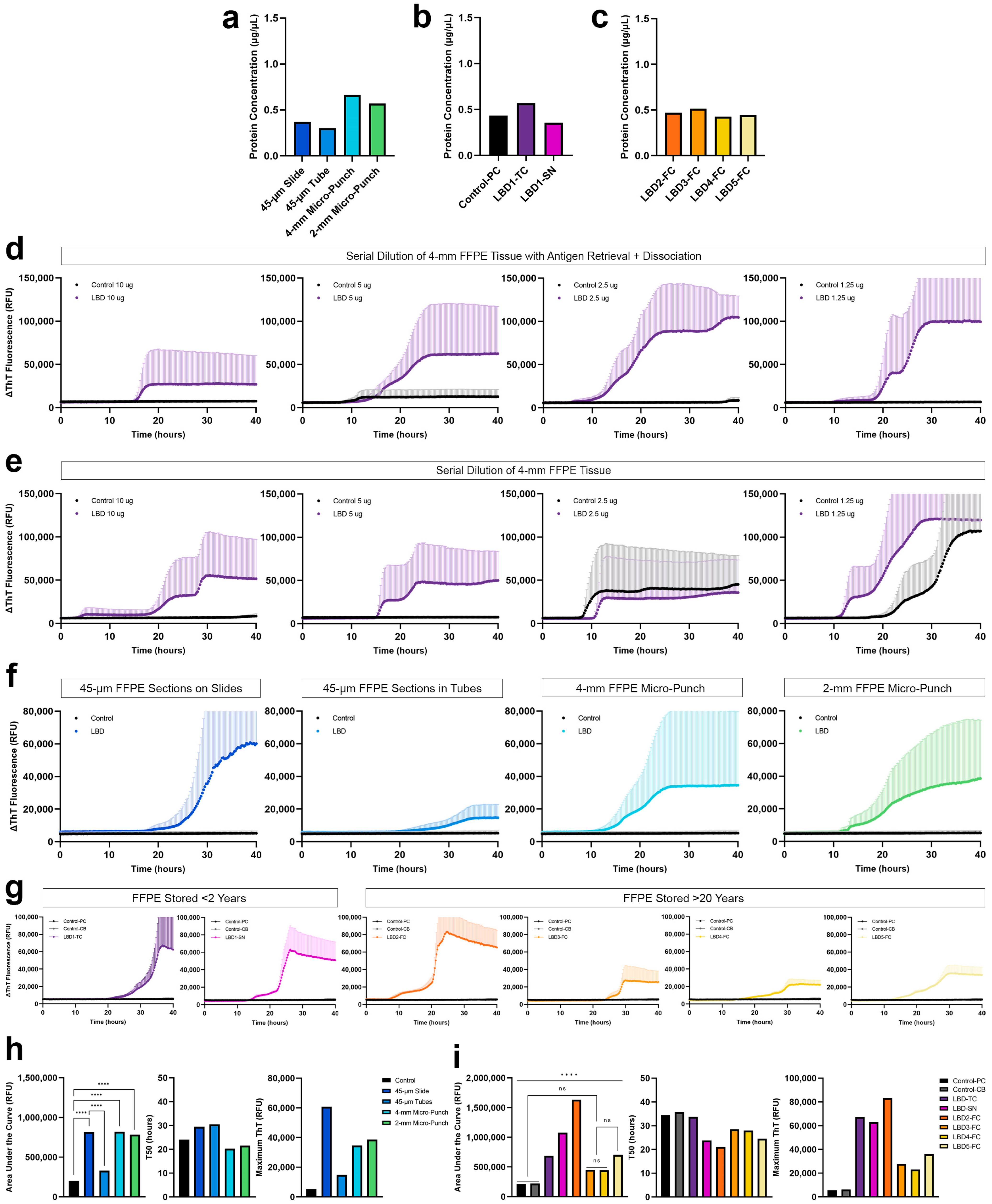
3.3. Optimization of FFPE Protein Extraction and αSyn SAA
3.3.1. FFPE Protein Extraction with Heat-Induced Antigen Retrieval and Dissociation
3.3.2. Optimization of αSyn SAA Using Proteins Extracted from FFPE
3.3.3. αSyn Seeding of Proteins Extracted from FFPE Tissue with Different Tissue Preparations
3.3.4. αSyn Seeding of Proteins Extracted from FFPE Tissue Archived for >20 Years
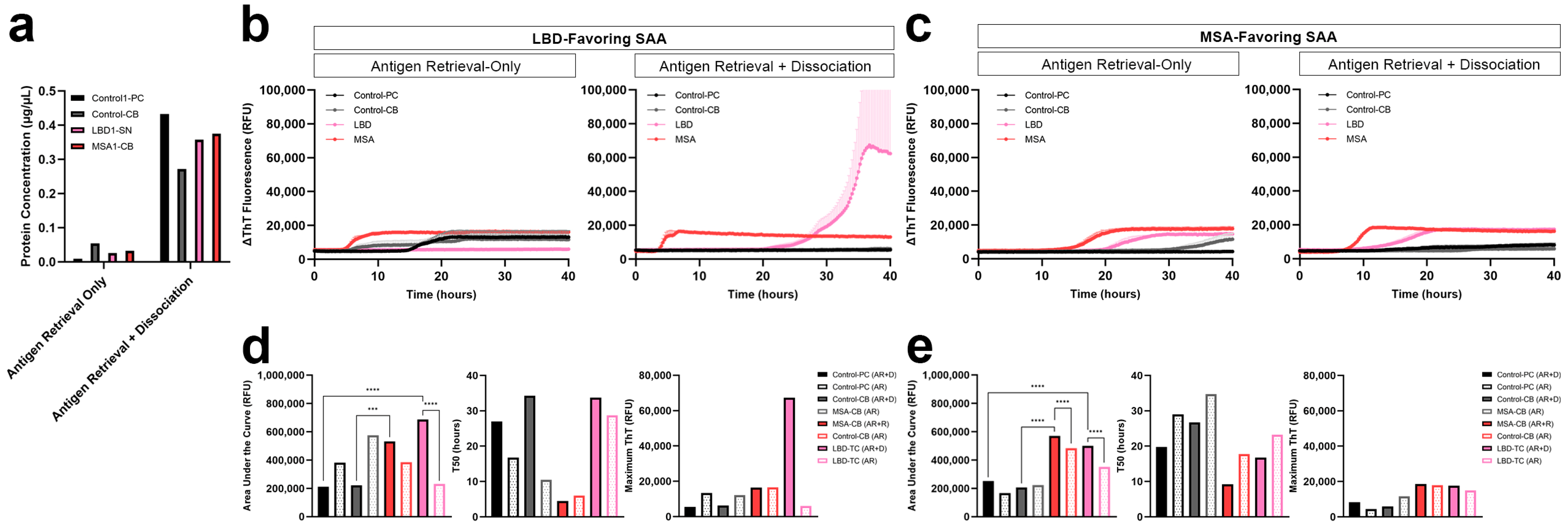
3.4. Comparison of Protein Extraction Efficiency and αSyn Seeding Activity with and without Dissociation
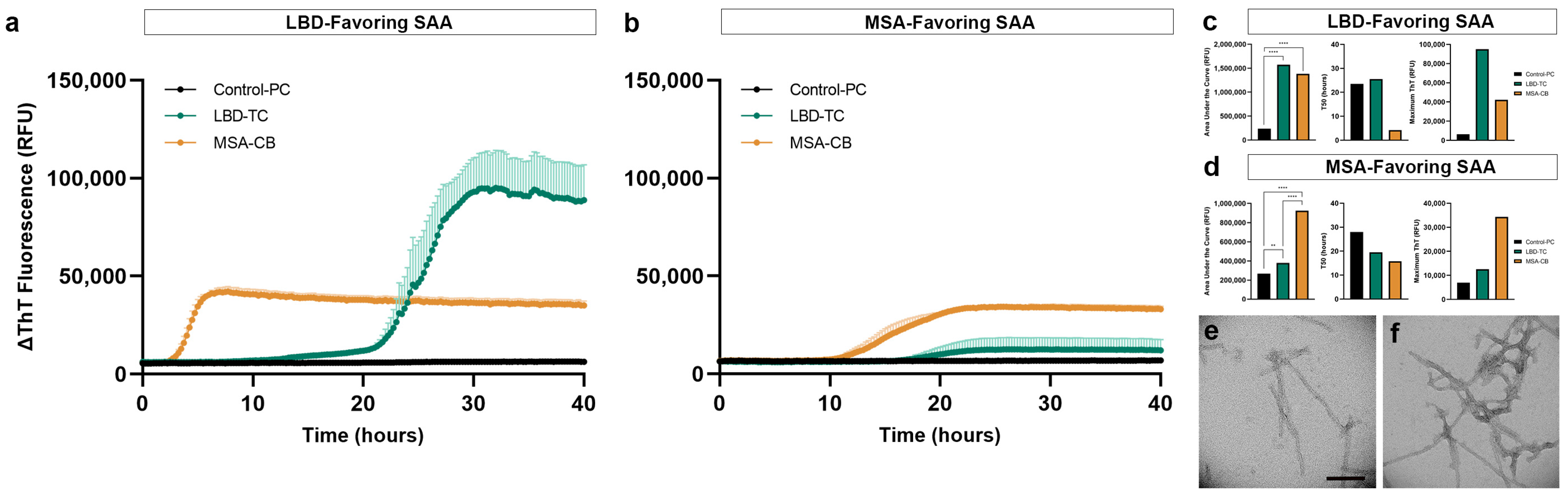
3.5. Replication of Disease-Specific Seeding Patterns in FFPE Human Brain Tissue
3.6. Structural Validation of Disease-Specific FFPE-Derived αSyn
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Maroteaux, L.; Campanelli, J.; Scheller, R. Synuclein: A Neuron-Specific Protein Localized to the Nucleus and Presynaptic Nerve Terminal. J. Neurosci. 1988, 8, 2804–2815. [Google Scholar] [CrossRef] [Green Version]
- Fujiwara, H.; Hasegawa, M.; Dohmae, N.; Kawashima, A.; Masliah, E.; Goldberg, M.S.; Shen, J.; Takio, K.; Iwatsubo, T. α-Synuclein Is Phosphorylated in Synucleinopathy Lesions. Nat. Cell Biol. 2002, 4, 160–164. [Google Scholar] [CrossRef]
- Vilar, M.; Chou, H.T.; Lührs, T.; Maji, S.K.; Riek-Loher, D.; Verel, R.; Manning, G.; Stahlberg, H.; Riek, R. The Fold of α-Synuclein Fibrils. Proc. Natl. Acad. Sci. USA 2008, 105, 8637–8642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovacs, G.G. Molecular Pathology of Neurodegenerative Diseases: Principles and Practice. J. Clin. Pathol. 2019, 72, 725–735. [Google Scholar] [CrossRef] [PubMed]
- Spillantini, M.G.; Schmidt, M.L.; Lee, V.M.-Y.; Trojanowski, J.Q.; Jakes, R.; Goedert, M. α-Synuclein in Lewy Bodies. Nature 1997, 388, 839–840. [Google Scholar] [CrossRef] [PubMed]
- Papp, M.I.; Kahn, J.E.; Lantos, P.L. Glial Cytoplasmic Inclusions in the CNS of Patients with Multiple System Atrophy (Striatonigral Degeneration, Olivopontocerebellar Atrophy and Shy-Drager Syndrome). J. Neurol. Sci. 1989, 94, 79–100. [Google Scholar] [CrossRef]
- Gilman, S.; Wenning, G.K.; Low, P.A.; Brooks, D.J.; Mathias, C.J.; Trojanowski, J.Q.; Wood, N.W.; Colosimo, C.; Durr, A.; Fowler, C.J.; et al. Second Consensus Statement on the Diagnosis of Multiple System Atrophy. Neurology 2008, 71, 670–676. [Google Scholar] [CrossRef] [PubMed]
- Bousset, L.; Pieri, L.; Ruiz-Arlandis, G.; Gath, J.; Jensen, P.H.; Habenstein, B.; Madiona, K.; Olieric, V.; Böckmann, A.; Meier, B.H.; et al. Structural and Functional Characterization of Two Alpha-Synuclein Strains. Nat. Commun. 2013, 4, 2575. [Google Scholar] [CrossRef] [Green Version]
- Van der Perren, A.; Gelders, G.; Fenyi, A.; Bousset, L.; Brito, F.; Peelaerts, W.; Van den Haute, C.; Gentleman, S.; Melki, R.; Baekelandt, V. The Structural Differences between Patient-Derived α-Synuclein Strains Dictate Characteristics of Parkinson’s Disease, Multiple System Atrophy and Dementia with Lewy Bodies. Acta Neuropathol. 2020, 139, 977–1000. [Google Scholar] [CrossRef] [PubMed]
- Schweighauser, M.; Shi, Y.; Tarutani, A.; Kametani, F.; Murzin, A.G.; Ghetti, B.; Matsubara, T.; Tomita, T.; Ando, T.; Hasegawa, K.; et al. Structures of α-Synuclein Filaments from Multiple System Atrophy. Nature 2020, 585, 464–469. [Google Scholar] [CrossRef]
- Yang, Y.; Shi, Y.; Schweighauser, M.; Zhang, X.; Kotecha, A.; Murzin, A.G.; Garringer, H.J.; Cullinane, P.W.; Saito, Y.; Foroud, T.; et al. Structures of α-Synuclein Filaments from Human Brains with Lewy Pathology. Nature 2022, 610, 791–795. [Google Scholar] [CrossRef]
- Lau, A.; So, R.W.L.; Lau, H.H.C.; Sang, J.C.; Ruiz-Riquelme, A.; Fleck, S.C.; Stuart, E.; Menon, S.; Visanji, N.P.; Meisl, G.; et al. α-Synuclein Strains Target Distinct Brain Regions and Cell Types. Nat. Neurosci. 2020, 23, 21–31. [Google Scholar] [CrossRef]
- Telling, G.C.; Parchi, P.; DeArmond, S.J.; Cortelli, P.; Montagna, P.; Gabizon, R.; Mastrianni, J.; Lugaresi, E.; Gambetti, P.; Prusiner, S.B. Evidence for the Conformation of the Pathologic Isoform of the Prion Protein Enciphering and Propagating Prion Diversity. Science 1996, 274, 2079–2082. [Google Scholar] [CrossRef] [Green Version]
- Groveman, B.R.; Orrù, C.D.; Hughson, A.G.; Raymond, L.D.; Zanusso, G.; Ghetti, B.; Campbell, K.J.; Safar, J.; Galasko, D.; Caughey, B. Rapid and Ultra-Sensitive Quantitation of Disease-Associated α-Synuclein Seeds in Brain and Cerebrospinal Fluid by ASyn RT-QuIC. Acta Neuropathol. Commun. 2018, 6, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Becker, K.; Donadio, V.; Siedlak, S.; Yuan, J.; Rezaee, M.; Incensi, A.; Kuzkina, A.; Orrú, C.D.; Tatsuoka, C.; et al. Skin α-Synuclein Aggregation Seeding Activity as a Novel Biomarker for Parkinson Disease. JAMA Neurol. 2021, 78, 30. [Google Scholar] [CrossRef] [PubMed]
- Biancalana, M.; Koide, S. Molecular Mechanism of Thioflavin-T Binding to Amyloid Fibrils. Biochim. Biophys. Acta-Proteins Proteom. 2010, 1804, 1405–1412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buell, A.K.; Galvagnion, C.; Gaspar, R.; Sparr, E.; Vendruscolo, M.; Knowles, T.P.J.; Linse, S.; Dobson, C.M. Solution Conditions Determine the Relative Importance of Nucleation and Growth Processes in α-Synuclein Aggregation. Proc. Natl. Acad. Sci. USA 2014, 111, 7671–7676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramis, R.; Ortega-Castro, J.; Vilanova, B.; Adrover, M.; Frau, J. Unraveling the NaCl Concentration Effect on the First Stages of α-Synuclein Aggregation. Biomacromolecules 2020, 21, 5200–5212. [Google Scholar] [CrossRef]
- Metrick, M.A.; do Carmo Ferreira, N.; Saijo, E.; Hughson, A.G.; Kraus, A.; Orrú, C.; Miller, M.W.; Zanusso, G.; Ghetti, B.; Vendruscolo, M.; et al. Million-Fold Sensitivity Enhancement in Proteopathic Seed Amplification Assays for Biospecimens by Hofmeister Ion Comparisons. Proc. Natl. Acad. Sci. USA 2019, 116, 23029–23039. [Google Scholar] [CrossRef]
- Martinez-Valbuena, I.; Visanji, N.P.; Kim, A.; Lau, H.H.C.; So, R.W.L.; Alshimemeri, S.; Gao, A.; Seidman, M.A.; Luquin, M.R.; Watts, J.C.; et al. Alpha-Synuclein Seeding Shows a Wide Heterogeneity in Multiple System Atrophy. Transl. Neurodegener. 2022, 11, 7. [Google Scholar] [CrossRef]
- Martinez-Valbuena, I.; Swinkin, E.; Santamaria, E.; Fernandez-Irigoyen, J.; Sackmann, V.; Kim, A.; Li, J.; Gonzalez-Latapi, P.; Kuhlman, G.; Bhowmick, S.S.; et al. α-Synuclein Molecular Behavior and Nigral Proteomic Profiling Distinguish Subtypes of Lewy Body Disorders. Acta Neuropathol. 2022, 144, 167–185. [Google Scholar] [CrossRef]
- Sutherland, B.W.; Toews, J.; Kast, J. Utility of Formaldehyde Cross-Linking and Mass Spectrometry in the Study of Protein–Protein Interactions. J. Mass Spectrom. 2008, 43, 699–715. [Google Scholar] [CrossRef] [PubMed]
- Loiacono, C.M.; Beckwith, N.; Kunkle, R.A.; Orcutt, D.; Hall, S.M. Detection of PrP Sc in Formalin-Fixed, Paraffin-Embedded Tissue by Western Blot Differentiates Classical Scrapie, Nor98 Scrapie, and Bovine Spongiform Encephalopathy. J. Vet. Diagn. Investig. 2010, 22, 684–689. [Google Scholar] [CrossRef] [Green Version]
- Priola, S.A.; Ward, A.E.; McCall, S.A.; Trifilo, M.; Choi, Y.P.; Solforosi, L.; Williamson, R.A.; Cruite, J.T.; Oldstone, M.B.A. Lack of Prion Infectivity in Fixed Heart Tissue from Patients with Creutzfeldt-Jakob Disease or Amyloid Heart Disease. J. Virol. 2013, 87, 9501–9510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoover, C.E.; Davenport, K.A.; Henderson, D.M.; Pulscher, L.A.; Mathiason, C.K.; Zabel, M.D.; Hoover, E.A. Detection and Quantification of CWD Prions in Fixed Paraffin Embedded Tissues by Real-Time Quaking-Induced Conversion. Sci. Rep. 2016, 6, 25098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, T.-T.-T.; Akagi, A.; Nonaka, T.; Nakagaki, T.; Mihara, B.; Takao, M.; Iwasaki, Y.; Nishida, N.; Satoh, K. Formalin RT-QuIC Assay Detects Prion-Seeding Activity in Formalin-Fixed Brain Samples from Sporadic Creutzfeldt–Jakob Disease Patients. Neurobiol. Dis. 2021, 159, 105504. [Google Scholar] [CrossRef]
- Watts, J.C. Calling α-Synuclein a Prion Is Scientifically Justifiable. Acta Neuropathol. 2019, 138, 505–508. [Google Scholar] [CrossRef]
- Manne, S.; Kondru, N.; Jin, H.; Serrano, G.E.; Anantharam, V.; Kanthasamy, A.; Adler, C.H.; Beach, T.G.; Kanthasamy, A.G. Blinded RT-QuIC Analysis of α-Synuclein Biomarker in Skin Tissue from Parkinson’s Disease Patients. Mov. Disord. 2020, 35, 2230–2239. [Google Scholar] [CrossRef]
- Manne, S.; Kondru, N.; Jin, H.; Anantharam, V.; Huang, X.; Kanthasamy, A.; Kanthasamy, A.G. A-Synuclein Real-Time Quaking-Induced Conversion in the Submandibular Glands of Parkinson’s Disease Patients. Mov. Disord. 2020, 35, 268–278. [Google Scholar] [CrossRef]
- Shin, C.; Han, J.Y.; Kim, S.I.; Park, S.H.; Yang, H.K.; Lee, H.J.; Kong, S.H.; Suh, Y.S.; Kim, H.J.; Choi, Y.P.; et al. In Vivo and Autopsy Validation of Alpha-Synuclein Seeding Activity Using RT-QuIC Assay in the Gastrointestinal Tract of Patients with Parkinson’s Disease. Park. Relat. Disord. 2022, 103, 23–28. [Google Scholar] [CrossRef]
- Fowler, C.B.; O’Leary, T.J.; Mason, J.T. Toward Improving the Proteomic Analysis of Formalin-Fixed, Paraffin-Embedded Tissue. Expert Rev. Proteom. 2013, 10, 389–400. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, E.A.; Frey, B.L.; Smith, L.M.; Auble, D.T. Formaldehyde Crosslinking: A Tool for the Study of Chromatin Complexes. J. Biol. Chem. 2015, 290, 26404–26411. [Google Scholar] [CrossRef] [Green Version]
- Kamps, J.J.A.G.; Hopkinson, R.J.; Schofield, C.J.; Claridge, T.D.W. How Formaldehyde Reacts with Amino Acids. Commun. Chem. 2019, 2, 126. [Google Scholar] [CrossRef] [Green Version]
- Giusti, L.; Lucacchini, A. Proteomic Studies of Formalin-Fixed Paraffin-Embedded Tissues. Expert Rev. Proteom. 2013, 10, 165–177. [Google Scholar] [CrossRef]
- Kawashima, Y.; Kodera, Y.; Singh, A.; Matsumoto, M.; Matsumoto, H. Efficient Extraction of Proteins from Formalin-Fixed Paraffin-Embedded Tissues Requires Higher Concentration of Tris(Hydroxymethyl)Aminomethane. Clin. Proteom. 2014, 11, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, K.; Sun, J.; Cao, X.; Zhou, D.; Li, J. Comparison of Different Buffers for Protein Extraction from Formalin-Fixed and Paraffin-Embedded Tissue Specimens. PLoS ONE 2015, 10, e0142650. [Google Scholar] [CrossRef] [Green Version]
- O’Rourke, M.B.; Padula, M.P. Analysis of Formalin-Fixed, Paraffin-Embedded (FFPE) Tissue via Proteomic Techniques and Misconceptions of Antigen Retrieval. Biotechniques 2016, 60, 229–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thacker, J.S.; Andersen, D.; Liang, S.; Zieniewicz, N.; Trivino-Paredes, J.S.; Nahirney, P.C.; Christie, B.R. Unlocking the Brain: A New Method for Western Blot Protein Detection from Fixed Brain Tissue. J. Neurosci. Methods 2021, 348, 108995. [Google Scholar] [CrossRef]
- Addis, M.F.; Tanca, A.; Pagnozzi, D.; Crobu, S.; Fanciulli, G.; Cossu-Rocca, P.; Uzzau, S. Generation of High-Quality Protein Extracts from Formalin-Fixed, Paraffin-Embedded Tissues. Proteomics 2009, 9, 3815–3823. [Google Scholar] [CrossRef]
- Rodríguez-Rigueiro, T.; Valladares-Ayerbes, M.; Haz-Conde, M.; Blanco, M.; Aparicio, G.; Fernández-Puente, P.; Blanco, F.J.; Lorenzo, M.J.; Aparicio, L.A.; Figueroa, A. A Novel Procedure for Protein Extraction from Formalin-Fixed Paraffin-Embedded Tissues. Proteomics 2011, 11, 2555–2559. [Google Scholar] [CrossRef] [Green Version]
- Araújo, J.E.; Oliveira, E.; Otero-Glez, A.; Santos Nores, J.; Igrejas, G.; Lodeiro, C.; Capelo, J.L.; Santos, H.M. A Comprehensive Factorial Design Study of Variables Affecting Protein Extraction from Formalin-Fixed Kidney Tissue Samples. Talanta 2014, 119, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Dressler, F.F.; Schoenfeld, J.; Revyakina, O.; Vogele, D.; Kiefer, S.; Kirfel, J.; Gemoll, T.; Perner, S. Systematic Evaluation and Optimization of Protein Extraction Parameters in Diagnostic FFPE Specimens. Clin. Proteom. 2022, 19, 10. [Google Scholar] [CrossRef] [PubMed]
- Montine, T.J.; Phelps, C.H.; Beach, T.G.; Bigio, E.H.; Cairns, N.J.; Dickson, D.W.; Duyckaerts, C.; Frosch, M.P.; Masliah, E.; Mirra, S.S.; et al. National Institute on Aging–Alzheimer’s Association Guidelines for the Neuropathologic Assessment of Alzheimer’s Disease: A Practical Approach. Acta Neuropathol. 2012, 123, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Kovacs, G.G.; Wagner, U.; Dumont, B.; Pikkarainen, M.; Osman, A.A.; Streichenberger, N.; Leisser, I.; Verchère, J.; Baron, T.; Alafuzoff, I.; et al. An Antibody with High Reactivity for Disease-Associated α-Synuclein Reveals Extensive Brain Pathology. Acta Neuropathol. 2012, 124, 37–50. [Google Scholar] [CrossRef] [PubMed]
- Koo, H.-J.; Lee, H.-J.; Im, H. Sequence Determinants Regulating Fibrillation of Human α-Synuclein. Biochem. Biophys. Res. Commun. 2008, 368, 772–778. [Google Scholar] [CrossRef]
- Mansour, A.G.; Abou-Khalil, P.; Bejjani, N.; Chatila, R.; Dagher-Hamalian, C.; Faour, W.H. An Optimized Xylene-Free Protein Extraction Method Adapted to Formalin-Fixed Paraffin Embedded Tissue Sections for Western Blot Analysis. Histol. Histopathol. 2017, 32, 307–313. [Google Scholar] [CrossRef]
- Wolff, C.; Schott, C.; Porschewski, P.; Reischauer, B.; Becker, K.-F. Successful Protein Extraction from Over-Fixed and Long-Term Stored Formalin-Fixed Tissues. PLoS ONE 2011, 6, e16353. [Google Scholar] [CrossRef]
- Geoui, T.; Urlaub, H.; Plessmann, U.; Porschewski, P. Extraction of Proteins from Formalin-Fixed, Paraffin-Embedded Tissue Using the Qproteome Extraction Technique and Preparation of Tryptic Peptides for Liquid Chromatography/Mass Spectrometry Analysis. In Current Protocols in Molecular Biology; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2010. [Google Scholar]
- Mansour, A.; Chatila, R.; Bejjani, N.; Dagher, C.; Faour, W.H. A Novel Xylene-Free Deparaffinization Method for the Extraction of Proteins from Human Derived Formalin-Fixed Paraffin Embedded (FFPE) Archival Tissue Blocks. MethodsX 2014, 1, 90–95. [Google Scholar] [CrossRef] [Green Version]
- García-Vence, M.; Chantada-Vazquez, M.d.P.; Sosa-Fajardo, A.; Agra, R.; Barcia de la Iglesia, A.; Otero-Glez, A.; García-González, M.; Cameselle-Teijeiro, J.M.; Nuñez, C.; Bravo, J.J.; et al. Protein Extraction from FFPE Kidney Tissue Samples: A Review of the Literature and Characterization of Techniques. Front. Med. 2021, 8, 657313. [Google Scholar] [CrossRef]
- Becker, K.; Wang, X.; Vander Stel, K.; Chu, Y.; Kordower, J.; Ma, J. Detecting Alpha Synuclein Seeding Activity in Formaldehyde-Fixed MSA Patient Tissue by PMCA. Mol. Neurobiol. 2018, 55, 8728–8737. [Google Scholar] [CrossRef] [Green Version]
- Fenyi, A.; Duyckaerts, C.; Bousset, L.; Braak, H.; Del Tredici, K.; Melki, R. Seeding Propensity and Characteristics of Pathogenic ASyn Assemblies in Formalin-Fixed Human Tissue from the Enteric Nervous System, Olfactory Bulb, and Brainstem in Cases Staged for Parkinson’s Disease. Cells 2021, 10, 139. [Google Scholar] [CrossRef] [PubMed]
- Hepker, M.; Clabaugh, G.; Jin, H.; Kanthasamy, A.G. New Protocol for Kinetic Assay Seeding Ability Recovery “KASAR” from Formalin-Fixed Paraffin-Embedded Tissues. Front. Mol. Biosci. 2023, 10, 1087982. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Case | Sex | Age at Death | PMD (Hours) | Storage (Years) | ABC Score * | CAA | Lewy Body Pathology | Lewy Body Braak Stage | LATE-NC Stage | Other Neuropathological Diagnosis |
|---|---|---|---|---|---|---|---|---|---|---|
| Control 1 | F | 52 | 28 | 3 | None | None | None | None | None | Early Fahr Disease |
| LBD 1 | M | 74 | NA | 2 | A1B1C1 | Type 2 | Neocortical | Stage 5 | None | None |
| LBD 2 | F | 73 | NA | 26 | A2B3C3 | None | Neocortical | Stage 5 | Stage 2 | None |
| LBD 3 | F | 73 | 2.5 | 25 | A2B3C2 | None | Limbic | Stage 4 | Stage 2 | None |
| LBD 4 | F | 82 | 24–36 | 25 | A2B2C2 | None | Neocortical | Stage 5 | None | ARTAG Medial Temporal GM, WM, Brainstem |
| LBD 5 | F | 62 | 15 | 25 | A3B3C3 | None | Neocortical | Stage 5 | Stage 1 | None |
| LBD 6 | M | 78 | 40 | 3 | A3B2C1 | Type 2 | Neocortical | Stage 5 | Stage 2 | Meningioma Right Frontal |
| MSA 1 | F | 68 | 4 | 36 | A1B1C0 | Type 2 | None | None | None | ARTAG Medial Temporal WM |
| MSA 2 | M | 64 | NA | 3 | A0B1C0 | None | None | None | None | PART (Braak stage II), AGD (Stage II) |
| Buffer | Buffer Composition |
|---|---|
| Buffer 1 | 1 × Phosphate-Buffered Saline (pH 7.4) Protease inhibitor |
| Buffer 2 | 1 × Phosphate-Buffered Saline (pH 7.4) 0.01% β-mercaptoethanol Protease inhibitor |
| Buffer 3 | 1 × Tris Buffered Saline (50 mM Tris-HCl (pH 7.5) 150 mM NaCl) Protease inhibitor |
| Buffer 4 | 50 mM Tris-HCl (pH 8.0) Protease inhibitor |
| Buffer 5 | 50 mM Tris-HCl (pH 8.0) 1% Nonyl Phenoxypolyethoxylethanol-40 150 mM NaCl Protease inhibitor |
| Buffer 6 | 50 mM Tris-HCl (pH 8.0) 1% Nonyl Phenoxypolyethoxylethanol-40 Protease inhibitor |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, A.; Martinez-Valbuena, I.; Li, J.; Lang, A.E.; Kovacs, G.G. Disease-Specific α-Synuclein Seeding in Lewy Body Disease and Multiple System Atrophy Are Preserved in Formaldehyde-Fixed Paraffin-Embedded Human Brain. Biomolecules 2023, 13, 936. https://doi.org/10.3390/biom13060936
Kim A, Martinez-Valbuena I, Li J, Lang AE, Kovacs GG. Disease-Specific α-Synuclein Seeding in Lewy Body Disease and Multiple System Atrophy Are Preserved in Formaldehyde-Fixed Paraffin-Embedded Human Brain. Biomolecules. 2023; 13(6):936. https://doi.org/10.3390/biom13060936
Chicago/Turabian StyleKim, Ain, Ivan Martinez-Valbuena, Jun Li, Anthony E. Lang, and Gabor G. Kovacs. 2023. "Disease-Specific α-Synuclein Seeding in Lewy Body Disease and Multiple System Atrophy Are Preserved in Formaldehyde-Fixed Paraffin-Embedded Human Brain" Biomolecules 13, no. 6: 936. https://doi.org/10.3390/biom13060936