
HIV-1 Transcriptional Activator Tat Inhibits IL2 Expression by Preventing the Presence of Pol II on the IL2 Promoter

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Cultures
2.2. Quantitative Real Time PCR (qPCR)
2.3. HIV-1 Tat siRNA Transfection
2.4. Western Immunoblotting
2.5. Co-Immunoprecipitation
2.6. Chromatin Immunoprecipitation (ChIP) Assays
2.7. Statistical Analysis
3. Results
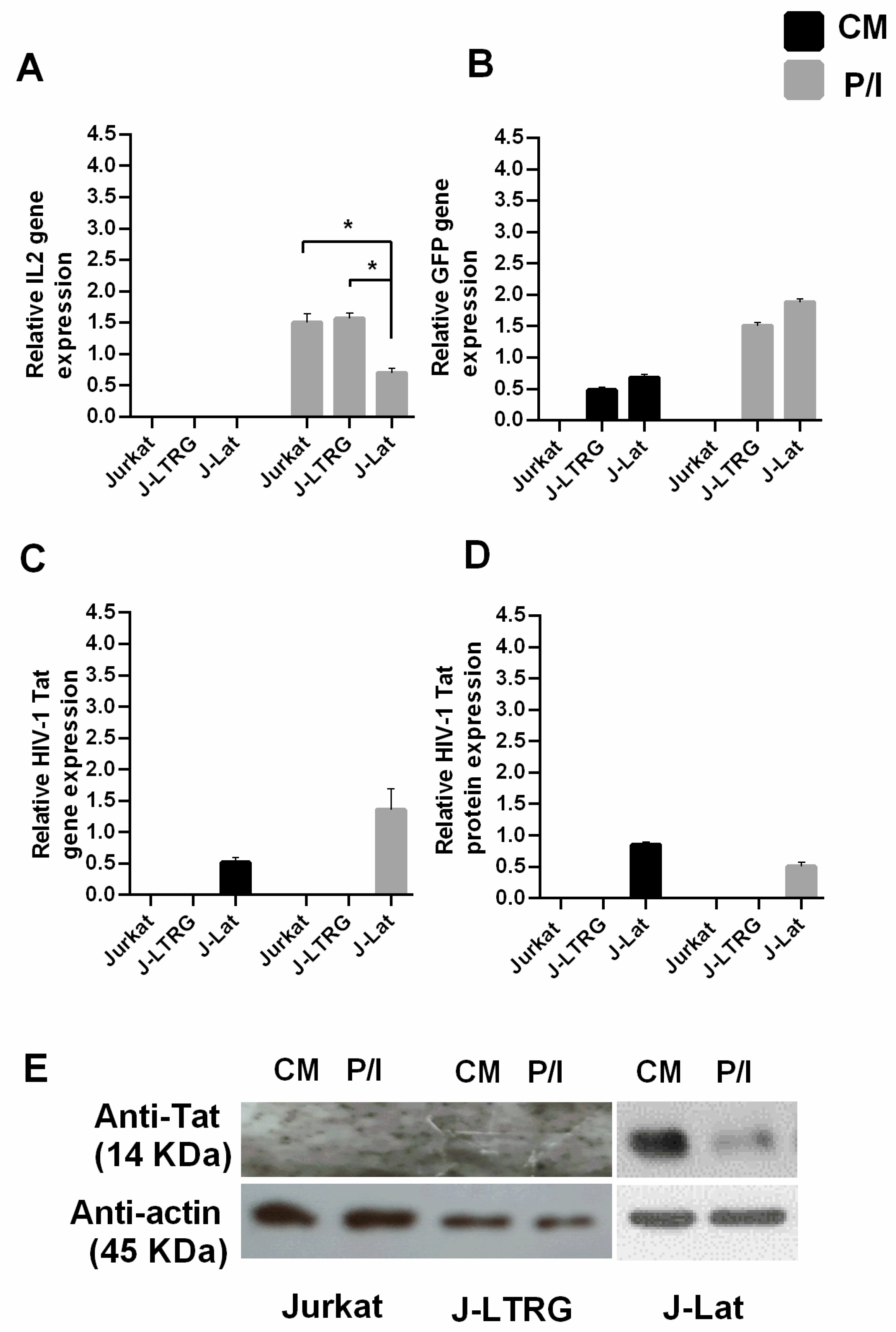
3.1. Decreased IL2 mRNA Expression in Induced J-Lat Cells Due to Tat Inhibition
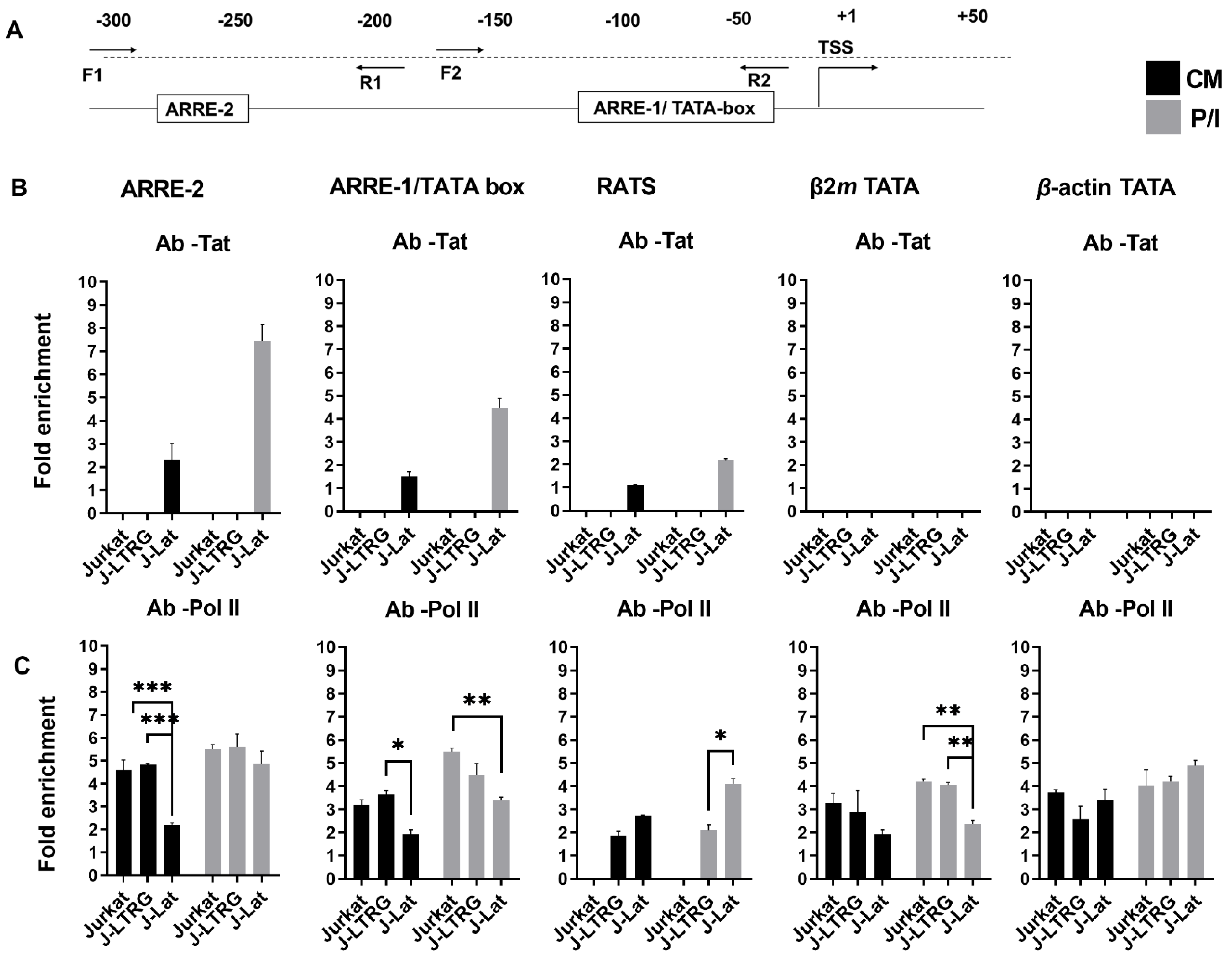
3.2. Tat Inhibits IL2 Expression by Preventing the Presence of Pol II on the IL2 Promoter
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- May, M.T.; Gompels, M.; Delpech, V.; Porter, K.; Orkin, C.; Kegg, S.; Hay, P.; Johnson, M.; Palfreeman, A.; Gilson, R.; et al. Impact on life expectancy of HIV-1 positive individuals of CD4+ cell count and viral load response to antiretroviral therapy. AIDS 2014, 28, 1193–1202. [Google Scholar] [CrossRef]
- Palmer, S.; Maldarelli, F.; Wiegand, A.; Bernstein, B.; Hanna, G.J.; Brun, S.C.; Kempf, D.J.; Mellors, J.W.; Coffin, J.M.; King, M.S. Low-level viremia persists for at least 7 years in patients on suppressive antiretroviral therapy. Proc. Natl. Acad. Sci. USA 2008, 105, 3879–3884. [Google Scholar] [CrossRef]
- Margolis, D.M.; Archin, N.M.; Cohen, M.S.; Eron, J.J.; Ferrari, G.; Garcia, J.V.; Gay, C.L.; Goonetilleke, N.; Joseph, S.B.; Swanstrom, R.; et al. Curing HIV: Seeking to Target and Clear Persistent Infection. Cell 2020, 181, 189–206. [Google Scholar] [CrossRef] [PubMed]
- Schrager, L.K.; D’Souza, M.P. Cellular and anatomical reservoirs of HIV-1 in patients receiving potent antiretroviral combination therapy. JAMA 1998, 280, 67–71. [Google Scholar] [CrossRef]
- Romani, B.; Allahbakhshi, E. Underlying mechanisms of HIV-1 latency. Virus Genes 2017, 53, 329–339. [Google Scholar] [CrossRef] [PubMed]
- Castro-Gonzalez, S.; Colomer-Lluch, M.; Serra-Moreno, R. Barriers for HIV Cure: The Latent Reservoir. AIDS Res. Hum. Retrovir. 2018, 34, 739–759. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, S.; Siliciano, R.F. Targeting the Latent Reservoir for HIV-1. Immunity 2018, 48, 872–895. [Google Scholar] [CrossRef]
- Mori, L.; Valente, S.T. Key Players in HIV-1 Transcriptional Regulation: Targets for a Functional Cure. Viruses 2020, 12, 529. [Google Scholar] [CrossRef]
- Rojas, M.; Luz-Crawford, P.; Soto-Rifo, R.; Reyes-Cerpa, S.; Toro-Ascuy, D. The Landscape of IFN/ISG Signaling in HIV-1-Infected Macrophages and Its Possible Role in the HIV-1 Latency. Cells 2021, 10, 2378. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, K.; Nakamura, H.; Koga, M.; Koibuchi, T.; Fujii, T.; Miura, T.; Iwamoto, A.; Kawana-Tachikawa, A. Imbalanced production of cytokines by T cells associates with the activation/exhaustion status of memory T cells in chronic HIV type 1 infection. AIDS Res. Hum. Retrovir. 2012, 28, 702–714. [Google Scholar] [CrossRef]
- Murphy, K.; Weaver, C. Janeway’s Immunobiology, 9th ed.; Garland Science, Taylor & Francis Group, LLC: New York, NY, USA; London, UK, 2017. [Google Scholar]
- Lane, H.C.; Fauci, A.S. Immunologic abnormalities in the acquired immunodeficiency syndrome. Annu. Rev. Immunol. 1985, 3, 477–500. [Google Scholar] [CrossRef]
- Nakayama-Hosoya, K.; Ishida, T.; Youngblood, B.; Nakamura, H.; Hosoya, N.; Koga, M.; Koibuchi, T.; Iwamoto, A.; Kawana-Tachikawa, A. Epigenetic repression of interleukin 2 expression in senescent CD4+ T cells during chronic HIV type 1 infection. J. Infect. Dis. 2015, 211, 28–39. [Google Scholar] [CrossRef]
- Purvis, S.F.; Georges, D.L.; Williams, T.M.; Lederman, M.M. Suppression of interleukin-2 and interleukin-2 receptor expression in Jurkat cells stably expressing the human immunodeficiency virus Tat protein. Cell Immunol. 1992, 144, 32–42. [Google Scholar] [CrossRef]
- González, E.; Punzón, C.; González, M.; Fresno, M. HIV-1 Tat inhibits IL-2 gene transcription through qualitative and quantitative alterations of the cooperative Rel/AP1 complex bound to the CD28RE/AP1 composite element of the IL-2 promoter. J. Immunol. 2001, 166, 4560–4569. [Google Scholar] [CrossRef] [PubMed]
- Shin, Y.; Yoon, C.-H.; Lim, H.; Park, J.; Roh, T.-Y.; Kang, C.; Choi, B.-S. Impaired IL-2 expression in latent HIV-1 infection. Biochem. Biophys. Res. Commun. 2015, 463, 1237–1242. [Google Scholar] [CrossRef] [PubMed]
- Seder, R.A.; Grabstein, K.H.; Berzofsky, J.A.; McDyer, J.F. Cytokine interactions in human immunodeficiency virus-infected individuals: Roles of interleukin (IL)-2, IL-12, and IL-15. J. Exp. Med. 1995, 182, 1067–1077. [Google Scholar] [CrossRef]
- Sellier, P.; Lafuente-Lafuente, C.; Bergmann, J.F. Interleukin-2 therapy in patients with HIV infection. N. Engl. J. Med. 2010, 362, 270–271. [Google Scholar] [PubMed]
- Chun, T.-W.; Engel, D.; Mizell, S.B.; Hallahan, C.W.; Fischette, M.; Park, S.; Davey, R.T.; Dybul, M.; Kovacs, J.A.; Metcalf, J.A.; et al. Effect of interleukin-2 on the pool of latently infected, resting CD4+ T cells in HIV-1-infected patients receiving highly active anti-retroviral therapy. Nat. Med. 1999, 5, 651–655. [Google Scholar] [CrossRef]
- Sullivan, A.K.; Hardy, G.A.; Nelson, M.R.; Gotch, F.; Gazzard, B.G.; Imami, N. Interleukin-2-associated viral breakthroughs induce HIV-1-specific CD4 T cell responses in patients on fully suppressive highly active antiretroviral therapy. AIDS 2003, 17, 628–629. [Google Scholar] [CrossRef] [PubMed]
- Bunting, K.; Wang, J.; Shannon, M.F. Control of interleukin-2 gene transcription: A paradigm for inducible, tissue-specific gene expression. Vitam Horm. 2006, 74, 105–145. [Google Scholar]
- Serfling, E.; Avots, A.; Neumann, M. The architecture of the interleukin-2 promoter: A reflection of T lymphocyte activation. Biochim. Biophys. Acta. 1995, 1263, 181–200. [Google Scholar] [CrossRef]
- Chow, C.W.; Rincón, M.; Davis, R.J. Requirement for transcription factor NFAT in interleukin-2 expression. Mol. Cell. Biol. 1999, 19, 2300–2307. [Google Scholar] [CrossRef] [PubMed]
- Clark, E.; Nava, B.; Caputi, M. Tat is a multifunctional viral protein that modulates cellular gene expression and functions. Oncotarget 2017, 8, 27569–27581. [Google Scholar] [CrossRef]
- Mouzaki, A.; Doucet, A.; Mavroidis, E.; Muster, L.; Rungger, D. A repression-derepression mechanism regulating the transcription of human immunodeficiency virus type 1 in primary T cells. Mol. Med. 2000, 6, 377–390. [Google Scholar] [CrossRef]
- Panagoulias, I.; Georgakopoulos, T.; Aggeletopoulou, I.; Agelopoulos, M.; Thanos, D.; Mouzaki, A. Transcription Factor Ets-2 Acts as a Preinduction Repressor of Interleukin-2 (IL-2) Transcription in Naive T Helper Lymphocytes. J. Biol. Chem. 2016, 291, 26707–26721. [Google Scholar] [CrossRef]
- Panagoulias, I.; Karagiannis, F.; Aggeletopoulou, I.; Georgakopoulos, T.; Argyropoulos, C.P.; Akinosoglou, K.; Gogos, C.; Skoutelis, A.; Mouzaki, A. Ets-2 Acts as a Transcriptional Repressor of the Human Immunodeficiency Virus Type 1 through Binding to a Repressor-Activator Target Sequence of 5’-LTR. Front. Immunol. 2018, 8, 1924. [Google Scholar] [CrossRef]
- Pessler, F.; Cron, R.Q. Reciprocal regulation of the nuclear factor of activated T cells and HIV-1. Genes Immun. 2004, 5, 158–167. [Google Scholar] [CrossRef]
- Ne, E.; Palstra, R.J.; Mahmoudi, T. Transcription: Insights from the HIV-1 Promoter. Int. Rev. Cell. Mol. Biol. 2018, 335, 191–243. [Google Scholar]
- El Kharroubi, A.; Piras, G.; Zensen, R.; Martin, M.A. Transcriptional activation of the integrated chromatin-associated human immunodeficiency virus type 1 promoter. Mol. Cell. Biol. 1998, 18, 2535–2544. [Google Scholar] [CrossRef]
- Carvallo, L.; Lopez, L.; Fajardo, J.E.; Jaureguiberry-Bravo, M.; Fiser, A.; Berman, J.W. HIV-Tat regulates macrophage gene expression in the context of neuroAIDS. PLoS ONE 2017, 12, e0179882. [Google Scholar] [CrossRef]
- Ott, M.; Geyer, M.; Zhou, Q. The control of HIV transcription: Keeping RNA polymerase II on track. Cell. Host. Microbe 2011, 10, 426–435. [Google Scholar] [CrossRef]
- Mousseau, G.; Valente, S.T. Role of Host Factors on the Regulation of Tat-Mediated HIV-1 Transcription. Curr. Pharm. Des. 2017, 23, 4079–4090. [Google Scholar] [CrossRef]
- Macián, F.; Rao, A. Reciprocal modulatory interaction between human immunodeficiency virus type 1 Tat and transcription factor NFAT1. Mol. Cell. Biol. 1999, 19, 3645–3653. [Google Scholar] [CrossRef]
- Fiume, G.; Vecchio, E.; De Laurentiis, A.; Trimboli, F.; Palmieri, C.; Pisano, A.; Falcone, C.; Pontoriero, M.; Rossi, A.; Scialdone, A.; et al. Human immunodeficiency virus-1 Tat activates NF-κB via physical interaction with IκB-α and p65. Nucleic Acids Res. 2012, 40, 3548–3562. [Google Scholar] [CrossRef]
- Westendorp, M.O.; Li-Weber, M.; Frank, R.W.; Krammer, P.H. Human immunodeficiency virus type 1 Tat upregulates interleukin-2 secretion in activated T cells. J. Virol. 1994, 68, 4177–4185. [Google Scholar] [CrossRef]
- Vacca, A.; Farina, M.; Maroder, M.; Alesse, E.; Screpanti, I.; Frati, L.; Gulino, A. Human immunodeficiency virus type-1 tat enhances interleukin-2 promoter activity through synergism with phorbol ester and calcium-mediated activation of the NF-AT cis-regulatory motif. Biochem. Biophys. Res. Commun. 1994, 205, 467–474. [Google Scholar] [CrossRef]
- Kwon, H.-S.; Brent, M.M.; Getachew, R.; Jayakumar, P.; Chen, L.-F.; Schnolzer, M.; McBurney, M.W.; Marmorstein, R.; Greene, W.C.; Ott, M. Human immunodeficiency virus type 1 Tat protein inhibits the SIRT1 deacetylase and induces T cell hyperactivation. Cell. Host. Microbe 2008, 3, 158–167. [Google Scholar] [CrossRef]
- Markle, T.J.; Philip, M.; Brockman, M.A. HIV-1 Nef and T-cell activation: A history of contradictions. Future Virol. 2013, 8, 391–404. [Google Scholar] [CrossRef]
- Jordan, A.; Bisgrove, D.; Verdin, E. HIV Reproducibly Establishes a Latent Infection after Acute Infection of T Cells in Vitro. EMBO J. 2003, 22, 1868–1877. [Google Scholar] [CrossRef]
- Ochsenbauer-Jambor, C.; Jones, J.; Heil, M.; Zammit, K.P.; Kutsch, O. T-cell line for HIV drug screening using EGFP as a quantitative marker of HIV-1 replication. Biotechniques 2006, 40, 91–100. [Google Scholar] [CrossRef]
- Kutsch, O.; Levy, D.N.; Bates, P.J.; Decker, J.; Kosloff, B.R.; Shaw, G.M.; Priebe, W.; Benveniste, E.N. Bis-anthracycline antibiotics inhibit human immunodeficiency virus type 1 transcription. Antimicrob Agents Chemother. 2004, 48, 1652–1663. [Google Scholar] [CrossRef]
- Pereira, L.A.; Bentley, K.; Peeters, A.; Churchill, M.J.; Deacon, N.J. A compilation of cellular transcription factor interactions with the HIV-1 LTR promoter. Nucleic Acids Res. 2000, 28, 663–668. [Google Scholar] [CrossRef]
- Spina, C.A.; Anderson, J.; Archin, N.M.; Bosque, A.; Chan, J.; Famiglietti, M.; Greene, W.C.; Kashuba, A.; Lewin, S.R.; Margolis, D.M.; et al. An in-depth comparison of latent HIV-1 reactivation in multiple cell model systems and resting CD4+ T cells from aviremic patients. PLoS Pathog. 2013, 9, e1003834. [Google Scholar] [CrossRef]
- Telwatte, S.; Moron-Lopez, S.; Aran, D.; Kim, P.; Hsieh, C.; Joshi, S.; Montano, M.; Greene, W.C.; Butte, A.J.; Wong, J.K.; et al. Heterogeneity in HIV and cellular transcription profiles in cell line models of latent and productive infection: Implications for HIV latency. Retrovirology 2019, 16, 32. [Google Scholar] [CrossRef]
- Lata, S.; Ali, A.; Sood, V.; Raja, R.; Banerjea, A.C. HIV-1 Rev downregulates Tat expression and viral replication via modulation of NAD(P)H:quinine oxidoreductase 1 (NQO1). Nat. Commun. 2015, 6, 7244. [Google Scholar] [CrossRef]
- Hong, H.W.; Lee, S.W.; Myung, H. Induced degradation of Tat by nucleocapsid (NC) via the proteasome pathway and its effect on HIV transcription. Viruses 2013, 5, 1143–1152. [Google Scholar] [CrossRef]
- Dhamija, N.; Choudhary, D.; Ladha, J.S.; Pillai, B.; Mitra, D. Tat predominantly associates with host promoter elements in HIV-1-infected T-cells-regulatory basis of transcriptional repression of c-Rel. FEBS J. 2015, 282, 595–610. [Google Scholar] [CrossRef]
- Carroll, I.R.; Wang, J.; Howcroft, T.K.; Singer, D.S. HIV Tat represses transcription of the beta 2-microglobulin promoter. Mol. Immunol. 1998, 35, 1171–1178. [Google Scholar] [CrossRef]
- Kamine, J.; Elangovan, B.; Subramanian, T.; Coleman, D.; Chinnadurai, G. Identification of a cellular protein that specifically interacts with the essential cysteine region of the HIV-1 Tat transactivator. Virology 1996, 216, 357–366. [Google Scholar] [CrossRef]
- Frankel, A.D.; Biancalana, S.; Hudson, D. Activity of synthetic peptides from the Tat protein of human immunodeficiency virus type 1. Proc. Natl. Acad. Sci. USA 1989, 86, 7397–7401. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anastasopoulou, S.; Georgakopoulos, T.; Mouzaki, A. HIV-1 Transcriptional Activator Tat Inhibits IL2 Expression by Preventing the Presence of Pol II on the IL2 Promoter. Biomolecules 2023, 13, 881. https://doi.org/10.3390/biom13060881
Anastasopoulou S, Georgakopoulos T, Mouzaki A. HIV-1 Transcriptional Activator Tat Inhibits IL2 Expression by Preventing the Presence of Pol II on the IL2 Promoter. Biomolecules. 2023; 13(6):881. https://doi.org/10.3390/biom13060881
Chicago/Turabian StyleAnastasopoulou, Spyridoula, Tassos Georgakopoulos, and Athanasia Mouzaki. 2023. "HIV-1 Transcriptional Activator Tat Inhibits IL2 Expression by Preventing the Presence of Pol II on the IL2 Promoter" Biomolecules 13, no. 6: 881. https://doi.org/10.3390/biom13060881