
Nociceptor–Macrophage Interactions in Apical Periodontitis: How Biomolecules Link Inflammation with Pain


Abstract
:1. Introduction
2. Overview of Macrophages in Apical Periodontitis
3. Role of Macrophages in Alveolar Bone Resorption
4. Sensory Neuronal Activation in Apical Periodontitis
5. Role of Sensory Neuropeptides in Pain and Inflammation
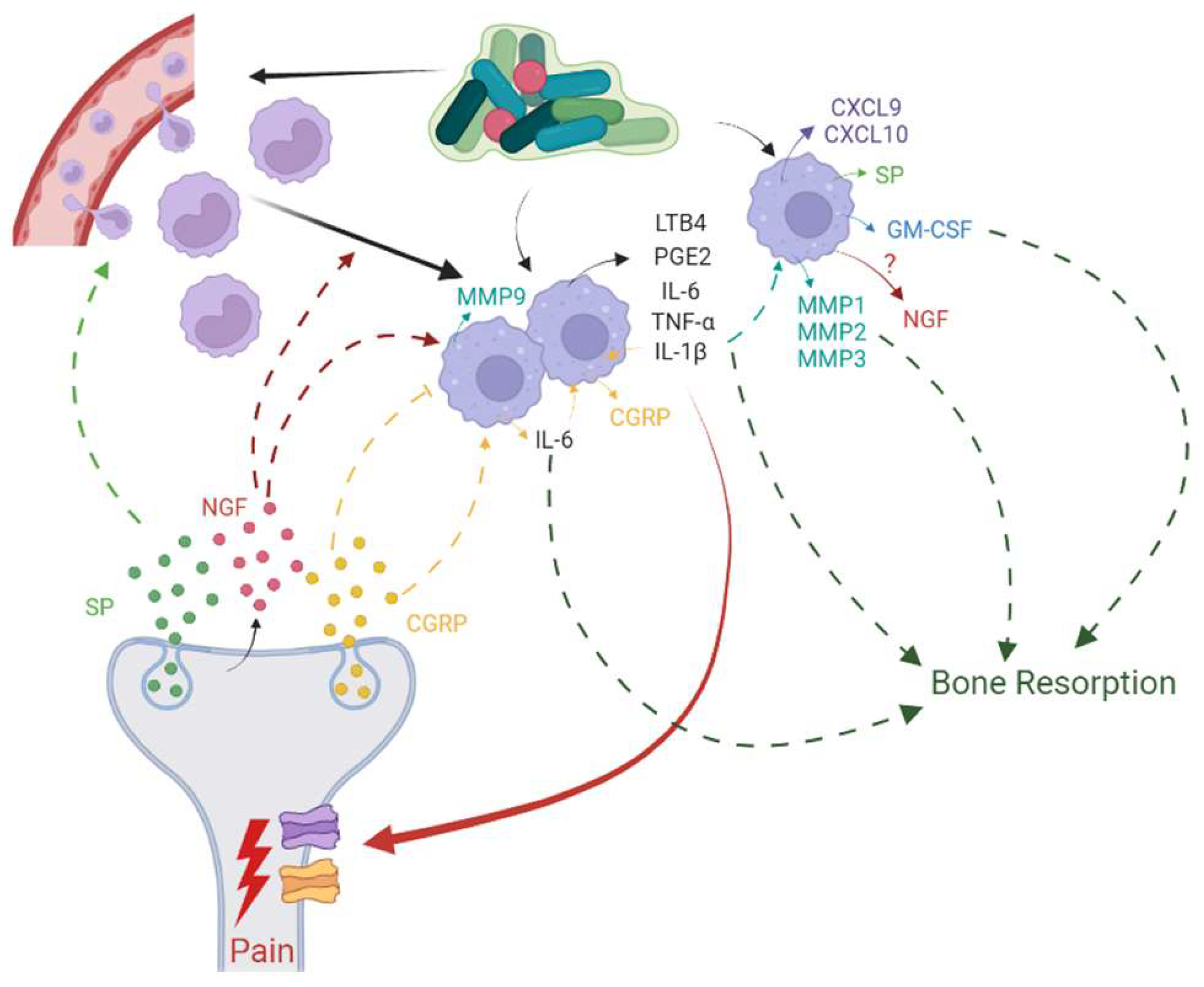
6. Neuro-Immune Interactions Modulate Inflammation and Healing in AP
7. Association of Pain with Nociceptor-Macrophage Cross-talk in AP
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A. List of Acronyms
| AP | Apical periodontitis |
| BMDMs | Bone-marrow-derived macrophages |
| CAM-1 | Cellular adhesion molecule 1 |
| cAMP | Cyclical adenosine monophosphate |
| CD | Cluster of differentiation |
| CGRP | Calcitonin Gene-Related Peptide |
| CGRP-IR | CGRP immunoreactive |
| CLR | Calcitonin-like receptor |
| CNS | Central nervous system |
| CREB | cAMP response element binding |
| CXC-L | Chemokine C-X-C motif chemokine ligand |
| DAG | Diacylglycerol |
| DAMPs | Damage-associated molecular patterns |
| ERKs | Extracellular signal-regulated kinases |
| GCF | Gingival crevicular fluid |
| GM-CSF | Granulocyte-Macrophage Colony-Stimulating Factor |
| HO-1 | Hemeoxygenase-1 |
| IFN-γ | Interferon-γ |
| IL-1 | Interleukin-1 |
| IP3 | Inositol triphosphate |
| IRAK | Interleukin-1 receptor-associated kinase |
| LDCVs | Large dense core vesicles |
| LPS | Lipopolysaccharides |
| LTB4 | Leukotriene B4 |
| M1 | Classically activated macrophages |
| M2 | Alternatively activated macrophages |
| MAPK | Mitogen-activated protein kinase |
| MCP-1 | Monocyte chemoattractant protein-1 |
| MD2 | Myeloid differentiation 2 |
| MIP-2 | Macrophage Inflammatory Protein-2 |
| MMPs | Matrix metalloproteinases |
| MQ | Macrophage |
| MyD88 | Myeloid differentiation primary response 88 |
| NFκB | Nuclear Factor Kappa B |
| NGF | Nerve Growth Factor |
| NI | Neurogenic Inflammation |
| NKA | Neurokinin A |
| NPs | Neuropeptides |
| NP γ | Neuropeptide γ |
| NPK | Neuropeptide K |
| NPY | Neuropeptide Y |
| NRS-11 | Numeric Rating Scale |
| OPG | Osteoprotegerin |
| PACAP | Pituitary adenylate cyclase-activating peptide |
| PAI | Periapical index |
| PAMPs | Pathogen-associated molecular patterns |
| PDL | Periodontal ligament |
| PGE2 | Prostaglandin E2 |
| PI3K/Akt | Phosphatidylinositol 3-kinase/protein kinase B |
| PKC | Protein Kinase C |
| RANKL | Receptor activator of nuclear factor kappa-β ligand |
| SNARE | Soluble N-ethylmaleimide-sensitive factor attachment protein receptor |
| SP | Substance P |
| TG | Trigeminal ganglion |
| TGF-β | Transforming growth factor-β |
| TIMPs | Tissue inhibitors of metalloproteinases |
| TLR | Toll-like receptor |
| TNF-α | Tumor necrosis factor-α |
| TRAF-6 | Tumor necrosis factor Receptor Associated Factor-6 |
| TrkA | Tropomyosin receptor kinase A |
| TRP | Transient receptor potential |
| VIP | Vasoactive intestinal peptide |
References
- Nair, P.N.R. On the causes of persistent apical periodontitis: A review. Int. Endod. J. 2006, 39, 249–281. [Google Scholar] [CrossRef] [PubMed]
- Korona-Glowniak, I.; Piatek, D.; Fornal, E.; Lukowiak, A.; Gerasymchuk, Y.; Kedziora, A.; Bugla-Płoskonska, G.; Grywalska, E.; Bachanek, T.; Malm, A. Patterns of Oral Microbiota in Patients with Apical Periodontitis. J. Clin. Med. 2021, 10, 2707. [Google Scholar] [CrossRef] [PubMed]
- Hassell, M.T. Tissues and cells of the periodontium. Periodontology 2000 1993, 3, 9–38. [Google Scholar] [CrossRef]
- Nanci, A.; Bosshardt, D.D. Structure of periodontal tissues in health and disease*. Periodontology 2000 2006, 40, 11–28. [Google Scholar] [CrossRef] [PubMed]
- Hussein, H.; Kishen, A. Local Immunomodulatory Effects of Intracanal Medications in Apical Periodontitis. J. Endod. 2022, 48, 430–456. [Google Scholar] [CrossRef] [PubMed]
- Jonasson, P.; Kvist, T. Diagnosis of apical periodontitis in root-filled teeth. Clin. Dent. Rev. 2018, 2, 15. [Google Scholar] [CrossRef] [Green Version]
- Li, F.-C.; Kishen, A. 3D Organoids for Regenerative Endodontics. Biomolecules 2023, 13, 900. [Google Scholar] [CrossRef]
- Chiu, I.M.; von Hehn, C.A.; Woolf, C.J. Neurogenic inflammation and the peripheral nervous system in host defense and immunopathology. Nat. Neurosci. 2012, 15, 1063–1067. [Google Scholar] [CrossRef] [Green Version]
- Assas, B.M.; Pennock, J.I.; Miyan, J.A. Calcitonin gene-related peptide is a key neurotransmitter in the neuro-immune axis. Front. Neurosci. 2014, 8, 23. [Google Scholar] [CrossRef] [Green Version]
- Kabata, H.; Artis, D. Neuro-immune crosstalk and allergic inflammation. J. Clin. Investig. 2019, 129, 1475–1482. [Google Scholar] [CrossRef] [Green Version]
- Crosson, T.; Roversi, K.; Balood, M.; Othman, R.; Ahmadi, M.; Wang, J.-C.; Seadi Pereira, P.J.; Tabatabaei, M.; Couture, R.; Eichwald, T.; et al. Profiling of how nociceptor neurons detect danger—New and old foes. J. Intern. Med. 2019, 286, 268–289. [Google Scholar] [CrossRef] [PubMed]
- Baliu-Piqué, M.; Jusek, G.; Holzmann, B. Neuroimmunological communication via CGRP promotes the development of a regulatory phenotype in TLR4-stimulated macrophages. Eur. J. Immunol. 2014, 44, 3708–3716. [Google Scholar] [CrossRef] [PubMed]
- Ji, R.-R.; Xu, Z.-Z.; Gao, Y.-J. Emerging targets in neuroinflammation-driven chronic pain. Nat. Rev. Drug Discov. 2014, 13, 533–548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuda, M.; Huh, Y.; Ji, R.-R. Roles of inflammation, neurogenic inflammation, and neuroinflammation in pain. J. Anesth. 2019, 33, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Zehnder, M.; Belibasakis, G.N. On the dynamics of root canal infections—What we understand and what we don′t. Virulence 2015, 6, 216–222. [Google Scholar] [CrossRef] [Green Version]
- Azeredo, S.V.; Brasil, S.C.; Antunes, H.; Marques, F.V.; Pires, F.R.; Armada, L. Distribution of macrophages and plasma cells in apical periodontitis and their relationship with clinical and image data. J. Clin. 2017, 9, e1060–e1065. [Google Scholar] [CrossRef] [Green Version]
- Márton, I.J.; Kiss, C. Overlapping Protective and Destructive Regulatory Pathways in Apical Periodontitis. J. Endod. 2014, 40, 155–163. [Google Scholar] [CrossRef]
- Nair, P.N.R. Pathogenesis of Apical Periodontitis and the Causes of Endodontic Failures. Crit. Rev. Oral Biol. Med. 2004, 15, 348–381. [Google Scholar] [CrossRef] [Green Version]
- Akamine, A.; Anan, H.; Hamachi, T.; Maeda, K. A histochemical study of the behavior of macrophages during experimental apical periodontitis in rats. J. Endod. 1994, 20, 474–478. [Google Scholar] [CrossRef]
- Sims, M.R. A model of the anisotropic distribution of microvascular volume in the periodontal ligament of the mouse mandibular molar. Aust. Orthod. J. 1987, 10, 21–24. [Google Scholar]
- Masset, A.; Staszyk, C.; Gasse, H. The blood vessel system in the periodontal ligament of the equine cheek teeth—Part II: The micro-architecture and its functional implications in a constantly remodelling system. Ann. Anat. Anat. Anz. 2006, 188, 535–539. [Google Scholar] [CrossRef] [PubMed]
- Graunaite, I.; Lodiene, G.; Maciulskiene, V. Pathogenesis of apical periodontitis: A literature review. J. Oral Maxillofac. Res. 2012, 2, e1. [Google Scholar] [CrossRef] [PubMed]
- Cekici, A.; Kantarci, A.; Hasturk, H.; Van Dyke, T.E. Inflammatory and immune pathways in the pathogenesis of periodontal disease. Periodontology 2000 2014, 64, 57–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davies, L.C.; Jenkins, S.J.; Allen, J.E.; Taylor, P.R. Tissue-resident macrophages. Nat. Immunol. 2013, 14, 986–995. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, L.; Yu, C.; Yang, X.-F.; Wang, H. Monocyte and macrophage differentiation: Circulation inflammatory monocyte as biomarker for inflammatory diseases. Biomark. Res. 2014, 2, 1. [Google Scholar] [CrossRef] [Green Version]
- Molteni, M.; Gemma, S.; Rossetti, C. The Role of Toll-Like Receptor 4 in Infectious and Noninfectious Inflammation. Mediat. Inflamm. 2016, 2016, 6978936. [Google Scholar] [CrossRef] [Green Version]
- Rayees, S.; Rochford, I.; Joshi, J.C.; Joshi, B.; Banerjee, S.; Mehta, D. Macrophage TLR4 and PAR2 Signaling: Role in Regulating Vascular Inflammatory Injury and Repair. Front. Immunol. 2020, 11, 2091. [Google Scholar] [CrossRef]
- Perros, F.; Lambrecht, B.N.; Hammad, H. TLR4 signalling in pulmonary stromal cells is critical for inflammation and immunity in the airways. Respir. Res. 2011, 12, 125. [Google Scholar] [CrossRef] [Green Version]
- Garlet, G.P.; Giannobile, W.V. Macrophages: The Bridge between Inflammation Resolution and Tissue Repair? J. Dent. Res. 2018, 97, 1079–1081. [Google Scholar] [CrossRef]
- House, I.G.; Savas, P.; Lai, J.; Chen, A.X.Y.; Oliver, A.J.; Teo, Z.L.; Todd, K.L.; Henderson, M.A.; Giuffrida, L.; Petley, E.V.; et al. Macrophage-Derived CXCL9 and CXCL10 Are Required for Antitumor Immune Responses Following Immune Checkpoint Blockade. Clin. Cancer Res. 2020, 26, 487–504. [Google Scholar] [CrossRef] [Green Version]
- Yao, Y.; Cai, X.; Ren, F.; Ye, Y.; Wang, F.; Zheng, C.; Qian, Y.; Zhang, M. The Macrophage-Osteoclast Axis in Osteoimmunity and Osteo-Related Diseases. Front. Immunol. 2021, 12, 664871. [Google Scholar] [CrossRef] [PubMed]
- Italiani, P.; Boraschi, D. From Monocytes to M1/M2 Macrophages: Phenotypical vs. Functional Differentiation. Front. Immunol. 2014, 5, 514. [Google Scholar] [CrossRef] [PubMed]
- Lappin, D.F.; Macleod, C.P.; Kerr, A.; Mitchell, T.J.; Kinane, D.F. Anti-inflammatory cytokine IL-10 and T cell cytokine profile in periodontitis granulation tissue. Clin. Exp. Immunol. 2001, 123, 294–300. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Hua, F.; Ding, W.; Ding, K.; Zhang, Y.; Xu, C. The correlation between the Th17/Treg cell balance and bone health. Immun. Ageing 2020, 17, 30. [Google Scholar] [CrossRef]
- Stern, M.H.; Dreizen, S.; Mackler, B.F.; Selbst, A.G.; Levy, B.M. Quantitative analysis of cellular composition of human periapical granuloma. J. Endod. 1981, 7, 117–122. [Google Scholar] [CrossRef]
- Shapouri-Moghaddam, A.; Mohammadian, S.; Vazini, H.; Taghadosi, M.; Esmaeili, S.-A.; Mardani, F.; Seifi, B.; Mohammadi, A.; Afshari, J.T.; Sahebkar, A. Macrophage plasticity, polarization, and function in health and disease. J. Cell. Physiol. 2018, 233, 6425–6440. [Google Scholar] [CrossRef] [PubMed]
- Lukić, A.; Danilovic, V.; Petrović, R. Comparative immunohistochemical and quantitative analysis of inflammatory cells in symptomatic and asymptomatic chronic periapical lesions. Vojnosanit. Pregl. 2008, 65, 435–440. [Google Scholar] [CrossRef]
- Kawai, T.; Matsuyama, T.; Hosokawa, Y.; Makihira, S.; Seki, M.; Karimbux, N.Y.; Goncalves, R.B.; Valverde, P.; Dibart, S.; Li, Y.-P.; et al. B and T lymphocytes are the primary sources of RANKL in the bone resorptive lesion of periodontal disease. Am. J. Pathol. 2006, 169, 987–998. [Google Scholar] [CrossRef] [Green Version]
- Dallas, S.L.; Bonewald, L.F. Dynamics of the transition from osteoblast to osteocyte. Ann. N. Y. Acad. Sci. 2010, 1192, 437–443. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Sayáns, M.; Somoza-Martín, J.M.; Barros-Angueira, F.; Rey, J.M.G.; García-García, A. RANK/RANKL/OPG role in distraction osteogenesis. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod. 2010, 109, 679–686. [Google Scholar] [CrossRef]
- Wada, T.; Nakashima, T.; Hiroshi, N.; Penninger, J.M. RANKL–RANK signaling in osteoclastogenesis and bone disease. Trends Mol. Med. 2006, 12, 17–25. [Google Scholar] [CrossRef]
- Boyce, B.F.; Xing, L. Functions of RANKL/RANK/OPG in bone modeling and remodeling. Arch. Biochem. Biophys. 2008, 473, 139–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ushach, I.; Zlotnik, A. Biological role of granulocyte macrophage colony-stimulating factor (GM-CSF) and macrophage colony-stimulating factor (M-CSF) on cells of the myeloid lineage. J. Leukoc. Biol. 2016, 100, 481–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, M.S.; Kim, H.S.; Yeon, J.-T.; Choi, S.-W.; Chun, C.H.; Kwak, H.B.; Oh, J. GM-CSF Regulates Fusion of Mononuclear Osteoclasts into Bone-Resorbing Osteoclasts by Activating the Ras/ERK Pathway1. J. Immunol. 2009, 183, 3390–3399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hienz, S.A.; Paliwal, S.; Ivanovski, S. Mechanisms of Bone Resorption in Periodontitis. J. Immunol. Res. 2015, 2015, 615486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, G. Gingival crevicular fluid as a periodontal diagnostic indicator- II: Inflammatory mediators, host-response modifiers and chair side diagnostic aids. J. Med. Life 2013, 6, 7–13. [Google Scholar]
- Lee, Y.-M.; Fujikado, N.; Manaka, H.; Yasuda, H.; Iwakura, Y. IL-1 plays an important role in the bone metabolism under physiological conditions. Int. Immunol. 2010, 22, 805–816. [Google Scholar] [CrossRef] [Green Version]
- Jimi, E.; Shuto, T.; Koga, T. Macrophage colony-stimulating factor and interleukin-1 alpha maintain the survival of osteoclast-like cells. Endocrinology 1995, 136, 808–811. [Google Scholar] [CrossRef]
- Gilbert, L.; He, X.; Farmer, P.; Boden, S.; Kozlowski, M.; Rubin, J.; Nanes, M.S. Inhibition of Osteoblast Differentiation by Tumor Necrosis Factor-α*. Endocrinology 2000, 141, 3956–3964. [Google Scholar] [CrossRef]
- Lee, H.-L.; Yi, T.; Woo, K.M.; Ryoo, H.-M.; Kim, G.-S.; Baek, J.-H. Msx2 mediates the inhibitory action of TNF-α on osteoblast differentiation. Exp. Mol. Med. 2010, 42, 437–445. [Google Scholar] [CrossRef] [Green Version]
- Marahleh, A.; Kitaura, H.; Ohori, F.; Kishikawa, A.; Ogawa, S.; Shen, W.-R.; Qi, J.; Noguchi, T.; Nara, Y.; Mizoguchi, I. TNF-α Directly Enhances Osteocyte RANKL Expression and Promotes Osteoclast Formation. Front. Immunol. 2019, 10, 2925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willis, R.D.; DiCosimo, C.J. The absence of proprioceptive nerve endings in the human periodontal ligament: The role of periodontal mechanoreceptors in the reflex control of mastication. Oral Surg. Oral Med. Oral Pathol. 1979, 48, 108–115. [Google Scholar] [CrossRef] [PubMed]
- Byers, M.R. Sensory innervation of periodontal ligament of rat molars consists of unencapsulated Ruffini-like mechanoreceptors and free nerve endings. J. Comp. Neurol. 1985, 231, 500–518. [Google Scholar] [CrossRef] [PubMed]
- Byers, M.R. Segregation of NGF receptor in sensory receptors, nerves and local cells of teeth and periodontium demonstrated by EM immunocytochemistry. J. Neurocytol. 1990, 19, 765–775. [Google Scholar] [CrossRef] [PubMed]
- Lambrichts, I.; Creemers, J.; van Steenberghe, D. Morphology of neural endings in the human periodontal ligament: An electron microscopic study. J. Periodontal Res. 1992, 27, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Tabata, T.; Karita, K. Response properties of periodontal mechanosensitive fibers in the superior dental nerve of the cat. Exp. Neurol. 1986, 94, 469–478. [Google Scholar] [CrossRef]
- Karita, K.; Tabata, T. Response fields of the periodontal mechanosensitive units in the superior alveolar nerve of the cat. Exp. Neurol. 1985, 90, 558–565. [Google Scholar] [CrossRef]
- Tender, G.C.; Li, Y.-Y.; Cui, J.-G. Vanilloid receptor 1-positive neurons mediate thermal hyperalgesia and tactile allodynia. Spine J. 2008, 8, 351–358. [Google Scholar] [CrossRef]
- Christine, W.; Michael, G.H. Neuroanatomy, Touch Receptor; StatPearls Publishing: St. Petersburg, FL, USA, 2023. [Google Scholar]
- Mengel, M.K.C.; Jyväsjärvi, E.; Kniffki, K.-D. Identification and characterization of afferent periodontal C fibres in the cat. PAIN 1992, 48, 413–420. [Google Scholar] [CrossRef] [PubMed]
- Bender, I.B. Pulpal Pain Diagnosis—A Review. J. Endod. 2000, 26, 175–179. [Google Scholar] [CrossRef]
- Bayliss, W.M. On the origin from the spinal cord of the vaso-dilator fibres of the hind-limb, and on the nature of these fibres1. J. Physiol. 1901, 26, 173–209. [Google Scholar] [CrossRef] [PubMed]
- Lewis, T. The Blood Vessels of the Human Skin. Br. Med. J. 1926, 2, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bayliss, W.M. The Vaso-Motor System/by William M. Bayliss; Longmans, Green: New York, NY, USA, 1923. [Google Scholar]
- Langley, J.N. Antidromic action. J. Physiol. 1923, 57, 428–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Byers, M.R.; Taylor, P.E. Effect of Sensory Denervation on the Response of Rat Molar Pulp to Exposure Injury. J. Dent. Res. 1993, 72, 613–618. [Google Scholar] [CrossRef] [PubMed]
- Toledo Mauriño, J.J.; Fonseca-Camarillo, G.; Furuzawa-Carballeda, J.; Barreto-Zuñiga, R.; Martínez Benítez, B.; Granados, J.; Yamamoto-Furusho, J.K. TRPV Subfamily (TRPV2, TRPV3, TRPV4, TRPV5, and TRPV6) Gene and Protein Expression in Patients with Ulcerative Colitis. J. Immunol. Res. 2020, 2020, 2906845. [Google Scholar] [CrossRef]
- Jara-Oseguera, A.; Simon, A.S.; Rosenbaum, T. TRPV1: On the Road to Pain Relief. Curr. Mol. Pharmacol. 2008, 1, 255–269. [Google Scholar] [CrossRef]
- Samanta, A.; Hughes, T.E.T.; Moiseenkova-Bell, V.Y. Transient Receptor Potential (TRP) Channels. In Membrane Protein Complexes: Structure and Function; Harris, J.R., Boekema, E.J., Eds.; Springer: Singapore, 2018; pp. 141–165. [Google Scholar] [CrossRef]
- Zygmunt, P.M.; Högestätt, E.D. TRPA1. In Mammalian Transient Receptor Potential (TRP) Cation Channels: Volume I; Nilius, B., Flockerzi, V., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 583–630. [Google Scholar] [CrossRef]
- Cao, E.; Liao, M.; Cheng, Y.; Julius, D. TRPV1 structures in distinct conformations reveal activation mechanisms. Nature 2013, 504, 113–118. [Google Scholar] [CrossRef] [Green Version]
- Liao, M.; Cao, E.; Julius, D.; Cheng, Y. Structure of the TRPV1 ion channel determined by electron cryo-microscopy. Nature 2013, 504, 107–112. [Google Scholar] [CrossRef] [Green Version]
- Sappington, R.M.; Sidorova, T.; Long, D.J.; Calkins, D.J. TRPV1: Contribution to Retinal Ganglion Cell Apoptosis and Increased Intracellular Ca2+ with Exposure to Hydrostatic Pressure. Investig. Ophthalmol. Vis. Sci. 2009, 50, 717–728. [Google Scholar] [CrossRef] [Green Version]
- King, R.; Brain, S.D. Chapter 189—CGRP. In Handbook of Biologically Active Peptides, 2nd ed; Kastin, A.J., Ed.; Academic Press: Boston, MA, USA, 2013; pp. 1394–1401. [Google Scholar] [CrossRef]
- Russell, F.A.; King, R.; Smillie, S.-J.; Kodji, X.; Brain, S.D. Calcitonin Gene-Related Peptide: Physiology and Pathophysiology. Physiol. Rev. 2014, 94, 1099–1142. [Google Scholar] [CrossRef] [Green Version]
- Matteoli, M.; Haimann, C.; Torri-Tarelli, F.; Polak, J.M.; Ceccarelli, B.; De Camilli, P. Differential effect of alpha-latrotoxin on exocytosis from small synaptic vesicles and from large dense-core vesicles containing calcitonin gene-related peptide at the frog neuromuscular junction. Proc. Natl. Acad. Sci. USA 1988, 85, 7366–7370. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, E.; Fernandes, M.; Keeble, J. The functions of TRPA1 and TRPV1: Moving away from sensory nerves. Br. J. Pharmacol. 2012, 166, 510–521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Omari, S.A.; Adams, M.J.; Geraghty, D.P. Chapter Six—TRPV1 Channels in Immune Cells and Hematological Malignancies. In Advances in Pharmacology; Geraghty, D.P., Rash, L.D., Eds.; Academic Press: Cambridge, MA, USA, 2017; Volume 79, pp. 173–198. [Google Scholar]
- Naert, R.; López-Requena, A.; Talavera, K. TRPA1 Expression and Pathophysiology in Immune Cells. Int. J. Mol. Sci. 2021, 22, 11460. [Google Scholar] [CrossRef] [PubMed]
- Tamás, K.; Zsolt, B.; Erzsébet, L.; Enikő, T.P.; Nóra, E.; Ágnes, C.; Zoltán, P.; István, É.; Róbert, P.; Attila, T. Tissue-Specific Regulation of Microvascular Diameter: Opposite Functional Roles of Neuronal and Smooth Muscle Located Vanilloid Receptor-1. Mol. Pharmacol. 2008, 73, 1405. [Google Scholar] [CrossRef]
- Helley, M.P.; Abate, W.; Jackson, S.K.; Bennett, J.H.; Thompson, S.W.N. The expression of Toll-like receptor 4, 7 and co-receptors in neurochemical sub-populations of rat trigeminal ganglion sensory neurons. Neuroscience 2015, 310, 686–698. [Google Scholar] [CrossRef] [Green Version]
- Wadachi, R.; Hargreaves, K. Trigeminal nociceptors express TLR-4 and CD14: A mechanism for pain due to infection. J. Dent. Res. 2006, 85, 49–53. [Google Scholar] [CrossRef] [Green Version]
- Diogenes, A.; Ferraz, C.C.R.; Akopian, A.N.; Henry, M.A.; Hargreaves, K.M. LPS Sensitizes TRPV1 via Activation of TLR4 in Trigeminal Sensory Neurons. J. Dent. Res. 2011, 90, 759–764. [Google Scholar] [CrossRef]
- Meseguer, V.; Alpizar, Y.A.; Luis, E.; Tajada, S.; Denlinger, B.; Fajardo, O.; Manenschijn, J.-A.; Fernández-Peña, C.; Talavera, A.; Kichko, T.; et al. TRPA1 channels mediate acute neurogenic inflammation and pain produced by bacterial endotoxins. Nat. Commun. 2014, 5, 3125. [Google Scholar] [CrossRef] [Green Version]
- Ferraz, C.C.R.; Henry, M.A.; Hargreaves, K.M.; Diogenes, A. Lipopolysaccharide From Porphyromonas gingivalis Sensitizes Capsaicin-Sensitive Nociceptors. J. Endod. 2011, 37, 45–48. [Google Scholar] [CrossRef] [Green Version]
- Lagomarsino, V.N.; Kostic, A.D.; Chiu, I.M. Mechanisms of microbial–neuronal interactions in pain and nociception. Neurobiol. Pain 2021, 9, 100056. [Google Scholar] [CrossRef]
- Chiu, I.M.; Heesters, B.A.; Ghasemlou, N.; Von Hehn, C.A.; Zhao, F.; Tran, J.; Wainger, B.; Strominger, A.; Muralidharan, S.; Horswill, A.R.; et al. Bacteria activate sensory neurons that modulate pain and inflammation. Nature 2013, 501, 52–57. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Yang, Z.; Nie, Y.; Hou, B. Microbial Communities in the Extraradicular and Intraradicular Infections Associated With Persistent Apical Periodontitis. Front. Cell. Infect. Microbiol. 2022, 11, 798367. [Google Scholar] [CrossRef]
- Eftekhari, S.; Salvatore, C.A.; Calamari, A.; Kane, S.A.; Tajti, J.; Edvinsson, L. Differential distribution of calcitonin gene-related peptide and its receptor components in the human trigeminal ganglion. Neuroscience 2010, 169, 683–696. [Google Scholar] [CrossRef] [Green Version]
- Messlinger, K.; Balcziak, L.K.; Russo, A.F. Cross-talk signaling in the trigeminal ganglion: Role of neuropeptides and other mediators. J. Neural Transm. 2020, 127, 431–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Messlinger, K. The big CGRP flood—Sources, sinks and signalling sites in the trigeminovascular system. J. Headache Pain 2018, 19, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Zhang, Z.; Li, X.; Cai, J.; Li, D.; Du, J.; Zhang, B.; Xiang, D.; Li, N.; Li, Y. CGRP derived from cardiac fibroblasts is an endogenous suppressor of cardiac fibrosis. Cardiovasc. Res. 2020, 116, 1335–1348. [Google Scholar] [CrossRef]
- Kim, Y.J.; Granstein, R.D. Roles of calcitonin gene-related peptide in the skin, and other physiological and pathophysiological functions. Brain Behav. Immun. Health 2021, 18, 100361. [Google Scholar] [CrossRef] [PubMed]
- Holzmann, B. Modulation of immune responses by the neuropeptide CGRP. Amino Acids 2013, 45, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Jacobson, A.; Meerschaert, K.A.; Sifakis, J.J.; Wu, M.; Chen, X.; Yang, T.; Zhou, Y.; Anekal, P.V.; Rucker, R.A.; et al. Nociceptor neurons direct goblet cells via a CGRP-RAMP1 axis to drive mucus production and gut barrier protection. Cell 2022, 185, 4190–4205.e4125. [Google Scholar] [CrossRef]
- Sert, S.; Sakallioğlu, U.; Lütfioğlu, M.; Aydoğdu, A.; Acarel, E.; Günaydın, M. Neurogenic inflammation in periimplant and periodontal disease: A case–control split-mouth study. Clin. Oral Implant. Res. 2019, 30, 800–807. [Google Scholar] [CrossRef] [PubMed]
- Lundy, F.T.; Shaw, C.; McKinnell, J.; Lamey, P.-J.; Linden, G.J. Calcitonin gene-related peptide in gingival crevicular fluid in periodontal health and disease. J. Clin. Periodontol. 1999, 26, 212–216. [Google Scholar] [CrossRef] [PubMed]
- Nelson, M.T.; Huang, Y.; Brayden, J.E.; Hescheler, J.; Standen, N.B. Arterial dilations in response to calcitonin gene-related peptide involve activation of K+ channels. Nature 1990, 344, 770–773. [Google Scholar] [CrossRef]
- Ma, W.; Dumont, Y.; Vercauteren, F.; Quirion, R. Lipopolysaccharide induces calcitonin gene-related peptide in the RAW264. 7 macrophage cell line. Immunology 2010, 130, 399–409. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.; Quirion, R. Increased calcitonin gene-related peptide in neuroma and invading macrophages is involved in the up-regulation of interleukin-6 and thermal hyperalgesia in a rat model of mononeuropathy. J. Neurochem. 2006, 98, 180–192. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Tao, R.; Zhang, M.; Cao, X.; Wang, H.; Xue, L.; Wu, M. Expression of Calcitonin Gene-related Peptide in Rat Pulp and Periodontal Tissues by Indirect Immunofluorescence Method. Monoclon. Antibodies Immunodiagn. Immunother. 2013, 32, 404–408. [Google Scholar] [CrossRef] [Green Version]
- Niedermühlbichler, M.; Wiedermann, C.J. Suppression of superoxide release from human monocytes by somatostatin-related peptides. Regul. Pept. 1992, 41, 39–47. [Google Scholar] [CrossRef]
- Fox, F.E.; Kubin, M.; Cassin, M.; Niu, Z.; Hosoi, J.; Torii, H.; Granstein, R.D.; Trinchieri, G.; Rook, A.H. Calcitonin Gene-Related Peptide Inhibits Proliferation and Antigen Presentation by Human Peripheral Blood Mononuclear Cells: Effects on B7, Interleukin 10, and Interleukin12. J. Investig. Dermatol. 1997, 108, 43–48. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, N.; Matsuda, Y.; Sato, K.; de Jong, P.R.; Bertin, S.; Tabeta, K.; Yamazaki, K. Neuronal TRPV1 activation regulates alveolar bone resorption by suppressing osteoclastogenesis via CGRP. Sci. Rep. 2016, 6, 29294. [Google Scholar] [CrossRef]
- Duan, Y.; Liang, Y.; Yang, F.; Ma, Y. Neural Regulations in Tooth Development and Tooth–Periodontium Complex Homeostasis: A Literature Review. Int. J. Mol. Sci. 2022, 23, 14150. [Google Scholar] [CrossRef]
- Johnson, M.B.; Young, A.D.; Marriott, I. The Therapeutic Potential of Targeting Substance P/NK-1R Interactions in Inflammatory CNS Disorders. Front. Cell. Neurosci. 2017, 10, 296. [Google Scholar] [CrossRef] [Green Version]
- Zieglgänsberger, W. Substance P and pain chronicity. Cell Tissue Res. 2019, 375, 227–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mashaghi, A.; Marmalidou, A.; Tehrani, M.; Grace, P.M.; Pothoulakis, C.; Dana, R. Neuropeptide substance P and the immune response. Cell. Mol. Life Sci. 2016, 73, 4249–4264. [Google Scholar] [CrossRef] [Green Version]
- Graefe, S.B.; Mohiuddin, S.S. Biochemistry, Substance P.; StatPearls Publishing: St. Petersburg, FL, USA, 2022. [Google Scholar]
- Perner, C.; Flayer, C.H.; Zhu, X.; Aderhold, P.A.; Dewan, Z.N.A.; Voisin, T.; Camire, R.B.; Chow, O.A.; Chiu, I.M.; Sokol, C.L. Substance P Release by Sensory Neurons Triggers Dendritic Cell Migration and Initiates the Type-2 Immune Response to Allergens. Immunity 2020, 53, 1063–1077.e1067. [Google Scholar] [CrossRef] [PubMed]
- Pradeep, A.R.; Raj, S.; Aruna, G.; Chowdhry, S. Gingival crevicular fluid and plasma levels of neuropeptide Substance-P in periodontal health, disease and after nonsurgical therapy. J. Periodontal Res. 2009, 44, 232–237. [Google Scholar] [CrossRef] [PubMed]
- Avellán, N.-L.; Sorsa, T.; Tervahartiala, T.; Forster, C.; Kemppainen, P. Experimental tooth pain elevates substance P and matrix metalloproteinase-8 levels in human gingival crevice fluid. Acta Odontol. Scand. 2008, 66, 18–22. [Google Scholar] [CrossRef]
- Sacerdote, P.; Levrini, L. Peripheral Mechanisms of Dental Pain: The Role of Substance P. Mediat. Inflamm. 2012, 2012, 951920. [Google Scholar] [CrossRef]
- Niedermair, T.; Schirner, S.; Seebröker, R.; Straub, R.H.; Grässel, S. Substance P modulates bone remodeling properties of murine osteoblasts and osteoclasts. Sci. Rep. 2018, 8, 9199. [Google Scholar] [CrossRef] [Green Version]
- Yan, K.; Lin, Q.; Tang, K.; Liu, S.; Du, Y.; Yu, X.; Li, S. Substance P participates in periodontitis by upregulating HIF-1α and RANKL/OPG ratio. BMC Oral Health 2020, 20, 27. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Dorscher-Kim, J. Effects of Substance P on Blood Flow of the Dental Pulp in Dogs. In Proceedings of the Substance P and Neurokinins; Springer-Verlag New York Inc.: New York, NY, USA, 1987; pp. 197–199. [Google Scholar]
- Augustyniak, D.; Roszkowiak, J.; Wiśniewska, I.; Skała, J.; Gorczyca, D.; Drulis-Kawa, Z. Neuropeptides SP and CGRP Diminish the Moraxella catarrhalis Outer Membrane Vesicle- (OMV-) Triggered Inflammatory Response of Human A549 Epithelial Cells and Neutrophils. Mediat. Inflamm. 2018, 2018, 4847205. [Google Scholar] [CrossRef] [Green Version]
- Iwasaki, M.a.A.Y.; Kaunitz, J.D. Recent advances in vasoactive intestinal peptide physiology and pathophysiology: Focus on the gastrointestinal system [version 1; peer review: 4 approved]. F1000Research 2019, 8, 1629. [Google Scholar] [CrossRef] [Green Version]
- Linden, G.J.; Mullally, B.H.; Burden, D.J.; Lamey, P.-J.; Shaw, C.; Ardill, J.; Lundy, F.T. Changes in vasoactive intestinal peptide in gingival crevicular fluid in response to periodontal treatment. J. Clin. Periodontol. 2002, 29, 484–489. [Google Scholar] [CrossRef] [PubMed]
- Luthman, J.; Friskopp, J.; Dahliöf, G.; Ahlström, U.; Sjöström, L.; Johansson, O. Immunohistochemical study of neurochemical markers in gingiva obtained from periodontitis-affected sites. J. Periodontal Res. 1989, 24, 267–278. [Google Scholar] [CrossRef] [PubMed]
- Abualsaud, N.; Caprio, L.; Galli, S.; Krawczyk, E.; Alamri, L.; Zhu, S.; Gallicano, G.I.; Kitlinska, J. Neuropeptide Y/Y5 Receptor Pathway Stimulates Neuroblastoma Cell Motility Through RhoA Activation. Front. Cell Dev. Biol. 2021, 8, 627090. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, C.-Y.; Chen, W.-C.; Shi, Y.-C.; Wang, C.-M.; Lin, S.; He, H.-F. Regulation of neuropeptide Y in body microenvironments and its potential application in therapies: A review. Cell Biosci. 2021, 11, 151. [Google Scholar] [CrossRef]
- Chen, W.-c.; Liu, Y.-b.; Liu, W.-f.; Zhou, Y.-y.; He, H.-f.; Lin, S. Neuropeptide Y Is an Immunomodulatory Factor: Direct and Indirect. Front. Immunol. 2020, 11, 580378. [Google Scholar] [CrossRef]
- Haririan, H.; Andrukhov, O.; Böttcher, M.; Pablik, E.; Wimmer, G.; Moritz, A.; Rausch-Fan, X. Salivary neuropeptides, stress, and periodontitis. J. Periodontol. 2018, 89, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Winning, L.; El Karim, I.A.; Linden, G.J.; Irwin, C.R.; Killough, S.A.; Lundy, F.T. Differential regulation of NPY and SP receptor expression in STRO-1+ve PDLSCs by inflammatory cytokines. J. Periodontal Res. 2022, 57, 186–194. [Google Scholar] [CrossRef]
- Okragly, A.J.; Morin, S.M.; DeRosa, D.; Martin, A.P.; Johnson, K.W.; Johnson, M.P.; Benschop, R.J. Human mast cells release the migraine-inducing factor pituitary adenylate cyclase-activating polypeptide (PACAP). Cephalalgia 2018, 38, 1564–1574. [Google Scholar] [CrossRef]
- Maugeri, G.; D’Amico, A.G.; Musumeci, G.; Reglodi, D.; D’Agata, V. Effects of PACAP on Schwann Cells: Focus on Nerve Injury. Int. J. Mol. Sci. 2020, 21, 8233. [Google Scholar] [CrossRef]
- Kuri, B.A.; Chan, S.-A.; Smith, C.B. PACAP regulates immediate catecholamine release from adrenal chromaffin cells in an activity-dependent manner through a protein kinase C-dependent pathway. J. Neurochem. 2009, 110, 1214–1225. [Google Scholar] [CrossRef] [Green Version]
- Edvinsson, J.C.A.; Grell, A.-S.; Warfvinge, K.; Sheykhzade, M.; Edvinsson, L.; Haanes, K.A. Differences in pituitary adenylate cyclase-activating peptide and calcitonin gene-related peptide release in the trigeminovascular system. Cephalalgia 2020, 40, 1296–1309. [Google Scholar] [CrossRef] [PubMed]
- Delgado, M.; Munoz-Elias, E.J.; Gomariz, R.P.; Ganea, D. VIP and PACAP inhibit IL-12 production in LPS-stimulated macrophages. Subsequent effect on IFNγ synthesis by T cells. J. Neuroimmunol. 1999, 96, 167–181. [Google Scholar] [CrossRef]
- Trafton, J.A.; Abbadie, C.; Basbaum, A.I. Differential Contribution of Substance P and Neurokinin A to Spinal Cord Neurokinin-1 Receptor Signaling in the Rat. J. Neurosci. 2001, 21, 3656–3664. [Google Scholar] [CrossRef]
- Onaga, T. Tachykinin: Recent developments and novel roles in health and disease. Biomol. Concepts 2014, 5, 225–243. [Google Scholar] [CrossRef] [PubMed]
- Mishra, A.; Lal, G. Neurokinin receptors and their implications in various autoimmune diseases. Curr. Res. Immunol. 2021, 2, 66–78. [Google Scholar] [CrossRef] [PubMed]
- Lundy, F.T.; El Karim, I.A.; Linden, G.J. Neuropeptide Y (NPY) and NPY Y1 receptor in periodontal health and disease. Arch. Oral Biol. 2009, 54, 258–262. [Google Scholar] [CrossRef] [PubMed]
- Amara, S.G.; Jonas, V.; Rosenfeld, M.G.; Ong, E.S.; Evans, R.M. Alternative RNA processing in calcitonin gene expression generates mRNAs encoding different polypeptide products. Nature 1982, 298, 240–244. [Google Scholar] [CrossRef]
- Fisher, L.A.; Kikkawa, D.O.; Rivier, J.E.; Amara, S.G.; Evans, R.M.; Rosenfeld, M.G.; Vale, W.W.; Brown, M.R. Stimulation of noradrenergic sympathetic outflow by calcitonin gene-related peptide. Nature 1983, 305, 534–536. [Google Scholar] [CrossRef]
- Brain, S.D.; Williams, T.J.; Tippins, J.R.; Morris, H.R.; MacIntyre, I. Calcitonin gene-related peptide is a potent vasodilator. Nature 1985, 313, 54–56. [Google Scholar] [CrossRef]
- Iyengar, S.; Ossipov, M.H.; Johnson, K.W. The role of calcitonin gene–related peptide in peripheral and central pain mechanisms including migraine. Pain 2017, 158, 543. [Google Scholar] [CrossRef] [Green Version]
- Hay, D.L.; Garelja, M.L.; Poyner, D.R.; Walker, C.S. Update on the pharmacology of calcitonin/CGRP family of peptides: IUPHAR Review 25. Br. J. Pharmacol. 2018, 175, 3–17. [Google Scholar] [CrossRef]
- v. Euler, U.S.; Gaddum, J.H. An unidentified depressor substance in certain tissue extracts. J. Physiol. 1931, 72, 74–87. [Google Scholar] [CrossRef]
- Krause, J.E.; Takeda, Y.; Hershey, A.D. Structure, Functions, and Mechanisms of Substance P Receptor Action. J. Investig. Dermatol. 1992, 98, S2–S7. [Google Scholar] [CrossRef] [Green Version]
- Arslan, H.; Yıldız, E.D.; Köseoğlu, S. Effects of endodontic treatment on salivary levels of CGRP and substance P: A pilot study. Restor. Dent. Endod. 2020, 45, e40. [Google Scholar] [CrossRef]
- Wakisaka, S. Neuropeptides in the dental pulp: Distribution, origins, and correlation. J. Endod. 1990, 16, 67–69. [Google Scholar] [CrossRef] [PubMed]
- Caviedes-Bucheli, J.; Muñoz, H.R.; Azuero-Holguín, M.M.; Ulate, E. Neuropeptides in Dental Pulp: The Silent Protagonists. J. Endod. 2008, 34, 773–788. [Google Scholar] [CrossRef]
- Kato, J.; Wakisaka, S.; Kurisu, K. Immunohistochemical Changes in the Distribution of Nerve Fibers in the Periodontal Ligament during an Experimental Tooth Movement of the Rat Molar. Acta Anat. 2008, 157, 53–62. [Google Scholar] [CrossRef]
- Caviedes-Bucheli, J.; Castellanos, F.; Vasquez, N.; Ulate, E.; Munoz, H.R. The influence of two reciprocating single-file and two rotary-file systems on the apical extrusion of debris and its biological relationship with symptomatic apical periodontitis. A systematic review and meta-analysis. Int. Endod. J. 2016, 49, 255–270. [Google Scholar] [CrossRef] [PubMed]
- Lundy, F.T.; Mullally, B.H.; Burden, D.J.; Lamey, P.-J.; Shaw, C.; Linden, G.J. Changes in substance P and neurokinin A in gingival crevicular fluid in response to periodontal treatment. J. Clin. Periodontol. 2000, 27, 526–530. [Google Scholar] [CrossRef] [PubMed]
- Burgener, B.; Ford, A.R.; Situ, H.; Fayad, M.I.; Hao, J.J.; Wenckus, C.S.; Johnson, B.R.; BeGole, E.A.; George, A. Biologic Markers for Odontogenic Periradicular Periodontitis. J. Endod. 2010, 36, 1307–1310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stashenko, P.; Teles, R.; d’Souza, R. Periapical inflammatory responses and their modulation. Crit. Rev. Oral Biol. Med. 1998, 9, 498–521. [Google Scholar] [CrossRef] [PubMed]
- Takeda, K.; Akira, S. Toll-Like Receptors. Curr. Protoc. Immunol. 2007, 77, 14.12.11–14.12.13. [Google Scholar] [CrossRef] [PubMed]
- Lai, J.-P.; Ho, W.Z.; Kilpatrick, L.E.; Wang, X.; Tuluc, F.; Korchak, H.M.; Douglas, S.D. Full-length and truncated neurokinin-1 receptor expression and function during monocyte/macrophage differentiation. Proc. Natl. Acad. Sci. USA 2006, 103, 7771–7776. [Google Scholar] [CrossRef] [PubMed]
- Marriott, I.; Bost, K.L. IL-4 and IFN-γ Up-Regulate Substance P Receptor Expression in Murine Peritoneal Macrophages1. J. Immunol. 2000, 165, 182–191. [Google Scholar] [CrossRef]
- Simeonidis, S.; Castagliuolo, I.; Pan, A.; Liu, J.; Wang, C.-C.; Mykoniatis, A.; Pasha, A.; Valenick, L.; Sougioultzis, S.; Zhao, D.; et al. Regulation of the NK-1 receptor gene expression in human macrophage cells via an NF-κB site on its promoter. Proc. Natl. Acad. Sci. USA 2003, 100, 2957–2962. [Google Scholar] [CrossRef]
- Goldman, R.; Bar-Shavit, Z.; Shezen, E.; Terry, S.; Blumberg, S. Enhancement of Phagocytosis by Neurotensin, a Newly Found Biological Activity of the Neuropeptide. In Macrophages and Natural Killer Cells: Regulation and Function; Normann, S.J., Sorkin, E., Eds.; Springer US: Boston, MA, USA, 1982; pp. 133–141. [Google Scholar] [CrossRef]
- Hartung, H.-P. Activation of macrophages by neuropeptides. Brain Behav. Immun. 1988, 2, 275–281. [Google Scholar] [CrossRef]
- Ruff, M.R.; Wahl, S.M.; Pert, C.B. Substance P receptor-mediated chemotaxis of human monocytes. Peptides 1985, 6, 107–111. [Google Scholar] [CrossRef]
- Hartung, H.-P.; Toyka, K.V. Substance P, the Immune System and Inflammation. Int. Rev. Immunol. 1989, 4, 229–249. [Google Scholar] [CrossRef]
- Cozens, P.J.; Rowe, F.M. Substance-P is a potent inducer of tnf and IL-1 secretion by macrophages—A potential role for tnf in the pathogenesis of asthma. Immunobiology 1987, 175, 7. [Google Scholar]
- Saito, I.; Ishii, K.; Hanada, K.; Sato, O.; Maeda, T. Responses of calcitonin gene-related peptide-immunopositive nerve fibres in the periodontal ligament of rat molars to experimental tooth movement. Arch. Oral Biol. 1991, 36, 689–692. [Google Scholar] [CrossRef]
- Wakisaka, S.; Nishikawa, S.; Ichikawa, H.; Matsuo, S.; Takano, Y.; Akai, M. The distribution and origin of substance P-like immunoreactivity in the rat molar pulp and periodontal tissues. Arch. Oral Biol. 1985, 30, 813–818. [Google Scholar] [CrossRef]
- Heyeraas, K.J.; Kvinnsland, I.; Byers, M.R.; Jacobsen, E.B. Nerve fibers immunoreactive to protein gene product 9.5, calcitonin gene-related peptide, substance P, and neuropeptide Y in the dental pulp, periodontal ligament, and gingiva in cats. Acta Odontol. Scand. 1993, 51, 207–221. [Google Scholar] [CrossRef] [PubMed]
- Berggreen Jacobsen, E.; Fristad, I.; Heyeraas, K.J. Nerve fibers immunoreactive to calcitonin gene-related peptide, substance P, neuropeptide Y, and dopamine ß-hydroxylase in innervated and denervated oral tissues in ferrets. Acta Odontol. Scand. 1998, 56, 220–228. [Google Scholar] [CrossRef] [PubMed]
- Kimberly, C.L.; Byers, M.R. Inflammation of rat molar pulp and periodontium causes increased calcitonin gene-related peptide and axonal sprouting. Anat. Rec. 1988, 222, 289–300. [Google Scholar] [CrossRef]
- Khayat, B.G.; Byers, M.R.; Taylor, P.E.; Mecifi, K.; Kimberly, C.L. Responses of nerve fibers to pulpal inflammation and periapical lesions in rat molars demonstrated by calcitonin gene-related peptide immunocytochemistry. J. Endod. 1988, 14, 577–587. [Google Scholar] [CrossRef]
- Gašperšič, R.; Kovačič, U.; Cör, A.; Skalerič, U. Unilateral ligature-induced periodontitis influences the expression of neuropeptides in the ipsilateral and contralateral trigeminal ganglion in rats. Arch. Oral Biol. 2008, 53, 659–665. [Google Scholar] [CrossRef]
- Kabashima, H.; Nagata, K.; Maeda, K.; Iijima, T. Involvement of substance P, mast cells, TNF-α and ICAM-1 in the infiltration of inflammatory cells in human periapical granulomas. J. Oral Pathol. Med. 2002, 31, 175–180. [Google Scholar] [CrossRef]
- de Avila, E.D.; de Molon, R.S.; de Godoi Gonçalves, D.A.; Camparis, C.M. Relationship between levels of neuropeptide Substance P in periodontal disease and chronic pain: A literature review. J. Investig. Clin. Dent. 2014, 5, 91–97. [Google Scholar] [CrossRef]
- Ferrin, J.; Kirakodu, S.; Jensen, D.; Al-Attar, A.; Peyyala, R.; Novak, M.J.; Dawson, D.; Al-Sabbagh, M.; Stromberg, A.J.; Orraca, L.; et al. Gene expression analysis of neuropeptides in oral mucosa during periodontal disease in non-human primates. J. Periodontol. 2018, 89, 858–866. [Google Scholar] [CrossRef] [PubMed]
- Liinden, G.J.; McKinneil, J.; Shaw, C.; Lundy, F.T. Substance P and neurokinin A in gingival crevicular fluid in periodontal health and disease. J. Clin. Periodontol. 1997, 24, 799–803. [Google Scholar] [CrossRef]
- Hanioka, T.; Takaya, K.; Matsumori, Y.; Matsuse, R.; Shizukuishi, S. Relationship of the substance P to indicators of host response in human gingival crevicular fluid. J. Clin. Periodontol. 2000, 27, 262–266. [Google Scholar] [CrossRef] [PubMed]
- Lundy, F.T.; Salmon, A.L.; Lamey, P.-J.; Shaw, C.; Linden, G.J. Carboxypeptidase-mediated metabolism of calcitonin gene-related peptide in human gingival crevicular fluid—A rôle in periodontal inflammation? J. Clin. Periodontol. 2000, 27, 499–505. [Google Scholar] [CrossRef]
- Owan, I.; Ibaraki, K. The role of calcitonin gene-related peptide (CGRP) in macrophages: The presence of functional receptors and effects on proliferation and differentiation into osteoclast-like cells. Bone Miner. 1994, 24, 151–164. [Google Scholar] [CrossRef]
- Fernandez, S.; Knopf, M.A.; Bjork, S.K.; McGillis, J.P. Bone Marrow-Derived Macrophages Express Functional CGRP Receptors and Respond to CGRP by Increasing Transcription of c-fos and IL-6 mRNA. Cell. Immunol. 2001, 209, 140–148. [Google Scholar] [CrossRef] [PubMed]
- Frielingsdorf, H.; Simpson, D.R.; Thal, L.J.; Pizzo, D.P. Nerve growth factor promotes survival of new neurons in the adult hippocampus. Neurobiol. Dis. 2007, 26, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Ulupinar, E.; Jacquin, M.F.; Erzurumlu, R.S. Differential effects of NGF and NT-3 on embryonic trigeminal axon growth patterns. J. Comp. Neurol. 2000, 425, 202–218. [Google Scholar] [CrossRef]
- Minnone, G.; De Benedetti, F.; Bracci-Laudiero, L. NGF and Its Receptors in the Regulation of Inflammatory Response. Int. J. Mol. Sci. 2017, 18, 1028. [Google Scholar] [CrossRef] [Green Version]
- Salio, C.; Averill, S.; Priestley, J.V.; Merighi, A. Costorage of BDNF and neuropeptides within individual dense-core vesicles in central and peripheral neurons. Dev. Neurobiol. 2007, 67, 326–338. [Google Scholar] [CrossRef]
- Ehrhard, P.B.; Ganter, U.; Stalder, A.; Bauer, J.; Otten, U. Expression of functional trk protooncogene in human monocytes. Proc. Natl. Acad. Sci. USA 1993, 90, 5423–5427. [Google Scholar] [CrossRef]
- Kurihara, H.; Shinohara, H.; Yoshino, H.; Takeda, K.; Shiba, H. Neurotrophins in Cultured Cells from Periodontal Tissues. J. Periodontol. 2003, 74, 76–84. [Google Scholar] [CrossRef]
- Ochi, K.; Saito, I.; Hanada, K.; Maeda, T. Expression of TrkB-like immunoreactivity in non-neural cells of rat periodontal ligament. Arch. Oral Biol. 1997, 42, 455–464. [Google Scholar] [CrossRef] [PubMed]
- Caroleo, M.C.; Costa, N.; Bracci-Laudiero, L.; Aloe, L. Human monocyte/macrophages activate by exposure to LPS overexpress NGF and NGF receptors. J. Neuroimmunol. 2001, 113, 193–201. [Google Scholar] [CrossRef] [PubMed]
- la Sala, A.; Corinti, S.; Federici, M.; Saragovi, H.U.; Girolomoni, G. Ligand activation of nerve growth factor receptor TrkA protects monocytes from apoptosis. J. Leukoc. Biol. 2000, 68, 104–110. [Google Scholar] [CrossRef] [PubMed]
- Bracci-Laudiero, L.; Aloe, L.; Caroleo, M.C.; Buanne, P.; Costa, N.; Starace, G.; Lundeberg, T. Endogenous NGF regulates CGRP expression in human monocytes, and affects HLA-DR and CD86 expression and IL-10 production. Blood 2005, 106, 3507–3514. [Google Scholar] [CrossRef] [Green Version]
- Williams, K.S.; Killebrew, D.A.; Clary, G.P.; Seawell, J.A.; Meeker, R.B. Differential regulation of macrophage phenotype by mature and pro-nerve growth factor. J. Neuroimmunol. 2015, 285, 76–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, J.; Ramnath, R.D.; Zhi, L.; Tamizhselvi, R.; Bhatia, M. Substance P enhances NF-κB transactivation and chemokine response in murine macrophages via ERK1/2 and p38 MAPK signaling pathways. Am. J. Physiol. Cell Physiol. 2008, 294, C1586–C1596. [Google Scholar] [CrossRef]
- Lee, S.-K.; Pi, S.-H.; Kim, S.-H.; Min, K.-S.; Lee, H.-J.; Chang, H.-S.; Kang, K.-H.; Kim, H.-R.; Shin, H.-I.; Kim, E.-C. Substance P regulates macrophage inflammatory protein 3α/chemokine C-C ligand 20 (CCL20) with heme oxygenase-1 in human periodontal ligament cells. Clin. Exp. Immunol. 2007, 150, 567–575. [Google Scholar] [CrossRef]
- Keane, M.P.; Strieter, R.M.; Belperio, J.A. Macrophage inflammatory protein. In Encyclopedia of Respiratory Medicine; Laurent, G.J., Shapiro, S.D., Eds.; Academic Press: Oxford, UK, 2006; pp. 1–5. [Google Scholar] [CrossRef]
- Deshmane, S.L.; Kremlev, S.; Amini, S.; Sawaya, B.E. Monocyte Chemoattractant Protein-1 (MCP-1): An Overview. J. Interferon Cytokine Res. 2009, 29, 313–326. [Google Scholar] [CrossRef]
- Jiang, M.H.; Chung, E.; Chi, G.F.; Ahn, W.; Lim, J.E.; Hong, H.S.; Kim, D.W.; Choi, H.; Kim, J.; Son, Y. Substance P induces M2-type macrophages after spinal cord injury. Neuroreport 2012, 23, 786–792. [Google Scholar] [CrossRef]
- Hong, H.S.; Son, Y. Substance P ameliorates collagen II-induced arthritis in mice via suppression of the inflammatory response. Biochem. Biophys. Res. Commun. 2014, 453, 179–184. [Google Scholar] [CrossRef]
- Montana, G.; Lampiasi, N. Substance P Induces HO-1 Expression in RAW 264.7 Cells Promoting Switch towards M2-Like Macrophages. PLoS ONE 2016, 11, e0167420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, M.; Nakamura, K.; Kageyama, S.; Lawal, A.O.; Gong, K.W.; Bhetraratana, M.; Fujii, T.; Sulaiman, D.; Hirao, H.; Bolisetty, S.; et al. Myeloid HO-1 modulates macrophage polarization and protects against ischemia-reperfusion injury. JCI Insight 2019, 3, e120596. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.E.; Chung, E.; Son, Y. A neuropeptide, Substance-P, directly induces tissue-repairing M2 like macrophages by activating the PI3K/Akt/mTOR pathway even in the presence of IFNγ. Sci. Rep. 2017, 7, 9417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinho-Ribeiro, F.A.; Verri, W.A.; Chiu, I.M. Nociceptor Sensory Neuron–Immune Interactions in Pain and Inflammation. Trends Immunol. 2017, 38, 5–19. [Google Scholar] [CrossRef] [Green Version]
- Rechenberg, D.K.; Munir, A.; Zehnder, M. Correlation between the clinically diagnosed inflammatory process and periapical index scores in severely painful endodontically involved teeth. Int. Endod. J. 2021, 54, 172–180. [Google Scholar] [CrossRef] [PubMed]
- Wideman, T.H.; Edwards, R.R.; Walton, D.M.; Martel, M.O.; Hudon, A.; Seminowicz, D.A. The Multimodal Assessment Model of Pain: A Novel Framework for Further Integrating the Subjective Pain Experience Within Research and Practice. Clin. J. Pain 2019, 35, 212. [Google Scholar] [CrossRef]
- Wilcox, C.E.; Mayer, A.R.; Teshiba, T.M.; Ling, J.; Smith, B.W.; Wilcox, G.L.; Mullins, P.G. The Subjective Experience of Pain: An FMRI Study of Percept-Related Models and Functional Connectivity. Pain Med. 2015, 16, 2121–2133. [Google Scholar] [CrossRef] [Green Version]
- Tarnawski, L.; Olofsson, P.S. Inflammation neuroscience: Neuro-immune crosstalk and interfaces. Clin. Transl. Immunol. 2021, 10, e1352. [Google Scholar] [CrossRef]
- Takeda, M.; Matsumoto, S.; Sessle, B.J.; Shinoda, M.; Iwata, K. Peripheral and Central Mechanisms of Trigeminal Neuropathic and Inflammatory Pain. J. Oral Biosci. 2011, 53, 318–329. [Google Scholar] [CrossRef]
- Khan, A.A.; Owatz, C.B.; Schindler, W.G.; Schwartz, S.A.; Keiser, K.; Hargreaves, K.M. Measurement of Mechanical Allodynia and Local Anesthetic Efficacy in Patients with Irreversible Pulpitis and Acute Periradicular Periodontitis. J. Endod. 2007, 33, 796–799. [Google Scholar] [CrossRef]
- Owatz, C.B.; Khan, A.A.; Schindler, W.G.; Schwartz, S.A.; Keiser, K.; Hargreaves, K.M. The Incidence of Mechanical Allodynia in Patients with Irreversible Pulpitis. J. Endod. 2007, 33, 552–556. [Google Scholar] [CrossRef] [PubMed]
- Schou, W.S.; Ashina, S.; Amin, F.M.; Goadsby, P.J.; Ashina, M. Calcitonin gene-related peptide and pain: A systematic review. J. Headache Pain 2017, 18, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, X.; Wang, L.; Li, X.; Sahbaie, P.; Kingery, W.S.; Clark, J.D. Neuropeptides Contribute to Peripheral Nociceptive Sensitization by Regulating Interleukin-1β Production in Keratinocytes. Anesth. Analg. 2011, 113, 175. [Google Scholar] [CrossRef] [Green Version]
- Sun, R.-Q.; Lawand, N.B.; Willis, W.D. The role of calcitonin gene-related peptide (CGRP) in the generation and maintenance of mechanical allodynia and hyperalgesia in rats after intradermal injection of capsaicin. PAIN 2003, 104, 201–208. [Google Scholar] [CrossRef]
- França, G.M.D.; Carmo, A.F.d.; Costa Neto, H.; Andrade, A.L.D.L.d.; Lima, K.C.d.; Galvão, H.C. Macrophages subpopulations in chronic periapical lesions according to clinical and morphological aspects. Braz. Oral Res. 2019, 33, e047. [Google Scholar] [CrossRef] [Green Version]
- Toriya, Y.; Hashiguchi, I.; Maeda, K. Immunohistochemical examination of the distribution of macrophages and CGRP-immunoreactive nerve fibers in induced rat periapical lesions. Dent. Traumatol. 1997, 13, 6–12. [Google Scholar] [CrossRef] [PubMed]
- Pereira Faustino, I.S.; Azevedo, R.S.; Takahama, A. Metalloproteinases 2 and 9 Immunoexpression in Periapical Lesions from Primary Endodontic Infection: Possible Relationship with the Histopathological Diagnosis and the Presence of Pain. J. Endod. 2016, 42, 547–551. [Google Scholar] [CrossRef]
- Hong, C.-Y.; Lin, S.-K.; Kok, S.-H.; Cheng, S.-J.; Lee, M.-S.; Wang, T.-M.; Chen, C.-S.; Lin, L.-D.; Wang, J.-S. The role of lipopolysaccharide in infectious bone resorption of periapical lesion. J. Oral Pathol. Med. 2004, 33, 162–169. [Google Scholar] [CrossRef]
- Lin, S.-K.; Kok, S.-H.; Kuo, M.Y.-P.; Wang, T.-J.; Wang, J.-T.; Yeh, F.T.-C.; Hsiao, M.; Lan, W.-H.; Hong, C.-Y. Sequential expressions of MMP-1, TIMP-1, IL-6, and COX-2 genes in induced periapical lesions in rats. Eur. J. Oral Sci. 2002, 110, 246–253. [Google Scholar] [CrossRef]
- Jain, A.; Bahuguna, R. Role of matrix metalloproteinases in dental caries, pulp and periapical inflammation: An overview. J. Oral Biol. Craniofacial Res. 2015, 5, 212–218. [Google Scholar] [CrossRef] [Green Version]
- Barreiros, D.; Nelson Filho, P.; Paula-Silva, F.W.G.; Oliveira, K.M.H.d.; Lucisano, M.P.; Rossi, A.D.; Silva, L.A.B.; Küchler, E.C.; Silva, R.A.B. MMP2 and MMP9 are Associated with Apical Periodontitis Progression and Might be Modulated by TLR2 and MyD88. Braz. Dent. J. 2018, 29, 43–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luchian, I.; Goriuc, A.; Sandu, D.; Covasa, M. The Role of Matrix Metalloproteinases (MMP-8, MMP-9, MMP-13) in Periodontal and Peri-Implant Pathological Processes. Int. J. Mol. Sci. 2022, 23, 1806. [Google Scholar] [CrossRef] [PubMed]
- Laronha, H.; Caldeira, J. Structure and Function of Human Matrix Metalloproteinases. Cells 2020, 9, 1076. [Google Scholar] [CrossRef]
- Elgezawi, M.; Haridy, R.; Almas, K.; Abdalla, M.A.; Omar, O.; Abuohashish, H.; Elembaby, A.; Christine Wölfle, U.; Siddiqui, Y.; Kaisarly, D. Matrix Metalloproteinases in Dental and Periodontal Tissues and Their Current Inhibitors: Developmental, Degradational and Pathological Aspects. Int. J. Mol. Sci. 2022, 23, 8929. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Neuropeptides in AP | Cells Releasing NPs | Cells Expressing NP Receptors | Functional Effects in AP |
|---|---|---|---|
| Calcitonin Gene-Related Peptide (CGRP) | Sensory neurons [9,75] T cells [91] Monocytes [91] Endothelial cells [75] Fibroblasts [92,93] Adipocytes [75] | Sensory neurons [90] Arterial vessels, Mononuclear immune cells, Schwann cells [91] Th1 cells [94] Langerhans cells [94] Monocytes [94] Macrophages [94] Dendritic cells [94] CD34+ hematopoietic progenitor cells [94] B & T lymphocytes [94] Epithelial cells [95] | Extensive degradation, resulting in reduced levels of CGRP observed in the GCF of diseased periodontal tissues [96,97] Vasodilation [98] Activation of innate and adaptive immune cells [99,100] Increase in blood flow [101] Prevention of monocyte/macrophage activation [102,103] Stimulation of osteogenesis [104,105] Endothelial cell proliferation [101] |
| Substance P (SP) | Neurons [106,107] Astrocytes [108] Microglia [107,108] Epithelial cells [108] Endothelial cells [108] T cells [108] Macrophages [108] Dendritic cells [106,108] Eosinophils [108] Mesenchymal stem cells [108] Leukocytes [106] Natural killer cells [106] Mast cells [106] Neutrophils [106] | Epithelial cells [108] Endothelial cells [106,108,109] White blood cells [109] Fibroblasts [108,109] Neurons [90,108,109] Smooth muscle cells [106,108] B & T lymphocytes [106,108] Natural killer cells [108] Dendritic cells [108,110] Monocytes [106,108] Macrophages [106,108] Microglia [106,108] Astrocytes [106,108] Eosinophils [106,108] Mast cells [106,108] Neutrophils [106] | Increased levels observed in GCF of diseased periodontal tissues [96,111] Desensitization of nociceptors [108] Neuronal activation [112] Onset of pain and inflammation [113] Inhibition of osteoblast differentiation [114] Induction of osteoclastogenesis [115] Increase in blood flow [113,116] Reduction in pro-inflammatory response of neutrophil, epithelial cells [117] |
| Vasoactive Intestinal Polypeptide (VIP) | Neurons [118] T cells [118] B cells [118] Mast cells [118] Eosinophils [118] | Epithelial cells [118] T cells [118] Macrophages [118] Dendritic cells [118] Mast cells [118] Neutrophils [118] Smooth muscle cells [118] Endothelial cells [118] Pancreatic B cells [118] | Found in increased levels in GCF of periodontitis patients; reduced levels as inflammation reduced [119] VIP reactivity observed in gingival biopsy, but no discernible difference compared to clinically healthy sites [120] Promotion of bone regeneration [105] |
| Neuropeptide Y (NPY) | Neurons [121] Pancreatic β cells [122] Vascular smooth muscle cells [123] Monocytes [123] Macrophages [123] Enterocytes [123] Endothelial cells [121] Langerhans cells [123] Microglia [123] B & T lymphocytes [123] | Neurons [121] Monocytes [123] Macrophages [122,123] Lymphocytes [123] Dendritic Cells [123] Neutrophils [123] B & T Lymphocytes [123] Leukocytes [123] Microglia [123] Osteoblasts [122] Endothelial Cells [122] PDL Stem Cells [105] | Proposed to be involved in initiation and regulation of periodontitis [105] Reduced levels observed in diseased sites of periodontitis patients [96] High levels observed in the saliva of periodontitis patients [124] Promotion of osteogenesis [105] Increase in PDL stem cells’ osteogenic capacity in vitro [125] |
| Pituitary Adenylate Cyclase-Activating Peptide (PACAP) | Mast cells [126] Schwann cells [127] | Pancreatic cells [128] Sensory nerve terminals [128,129] Pituitary cells [127] Schwann cells [127] Satellite glial cells [129] Hepatocytes [129] Osteoblasts [129] | Promotion of bone regeneration [105] Dose-dependent inhibition of IL-12 in LPS-activated macrophages [130] |
| Neurokinin A (NKA) | Sensory neurons [131] | Sensory neurons [132] B & T cells [133] Dendritic cells [133] Macrophages [133] | Co-released with SP [131] Increased levels observed in the GCF of diseased periodontal tissues [134] Decreased levels of NK-A observed in diseased periodontal tissue [96] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Menon, N.; Kishen, A. Nociceptor–Macrophage Interactions in Apical Periodontitis: How Biomolecules Link Inflammation with Pain. Biomolecules 2023, 13, 1193. https://doi.org/10.3390/biom13081193
Menon N, Kishen A. Nociceptor–Macrophage Interactions in Apical Periodontitis: How Biomolecules Link Inflammation with Pain. Biomolecules. 2023; 13(8):1193. https://doi.org/10.3390/biom13081193
Chicago/Turabian StyleMenon, Nandita, and Anil Kishen. 2023. "Nociceptor–Macrophage Interactions in Apical Periodontitis: How Biomolecules Link Inflammation with Pain" Biomolecules 13, no. 8: 1193. https://doi.org/10.3390/biom13081193